Characterization of a Long Non-Coding RNA, the Antisense RNA of Na/K-ATPase α1 in Human Kidney Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
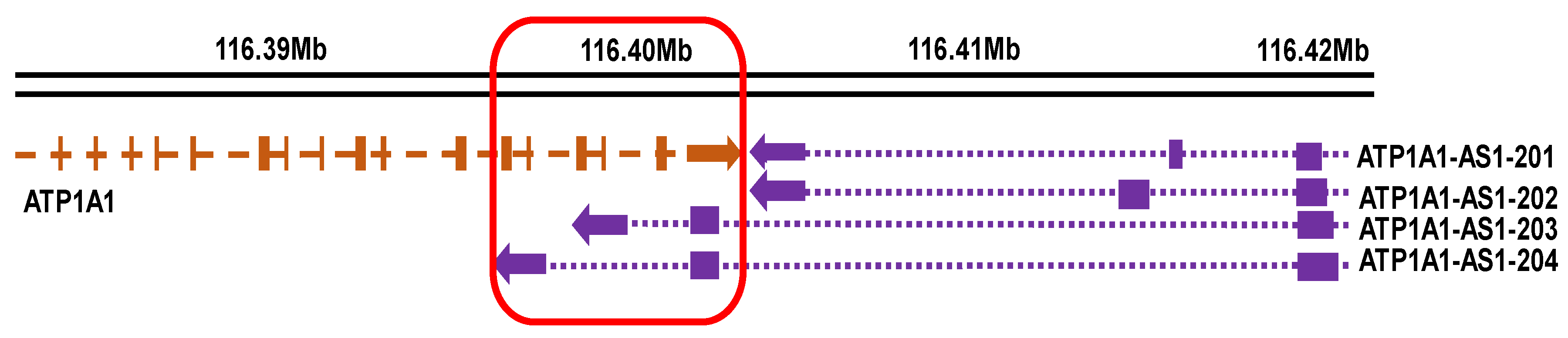
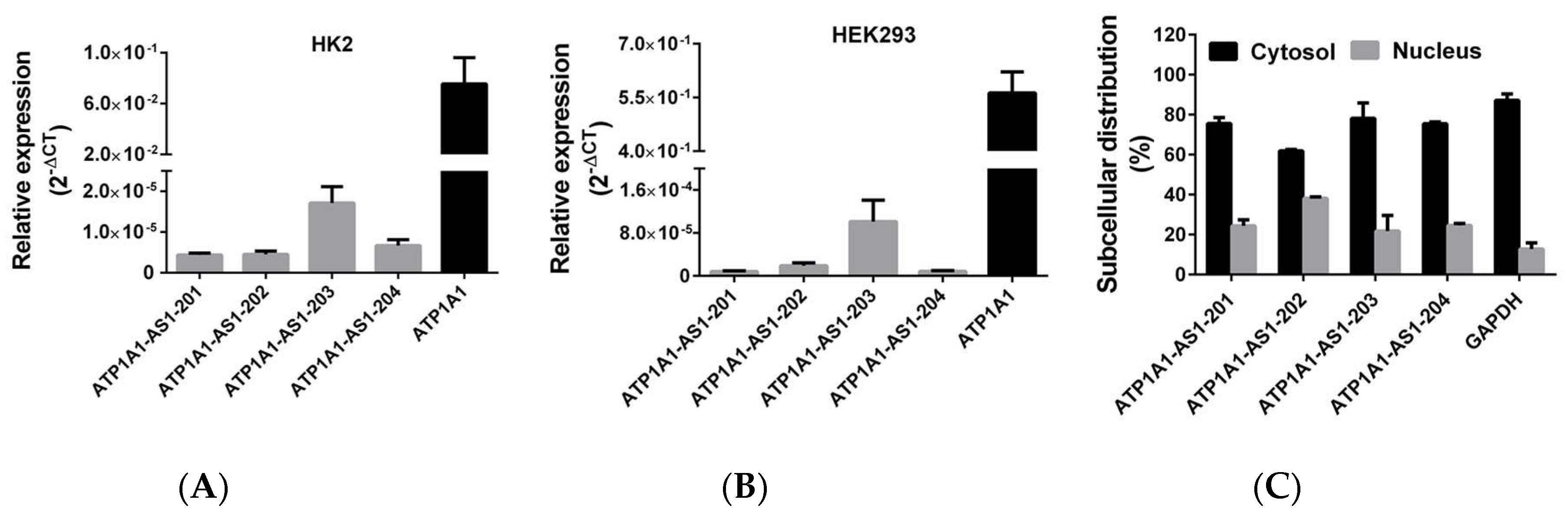
2.1. Differential Expression and Subcellular Distribution of ATP1A1-AS1 Splice Variants in Human Kidney Cells
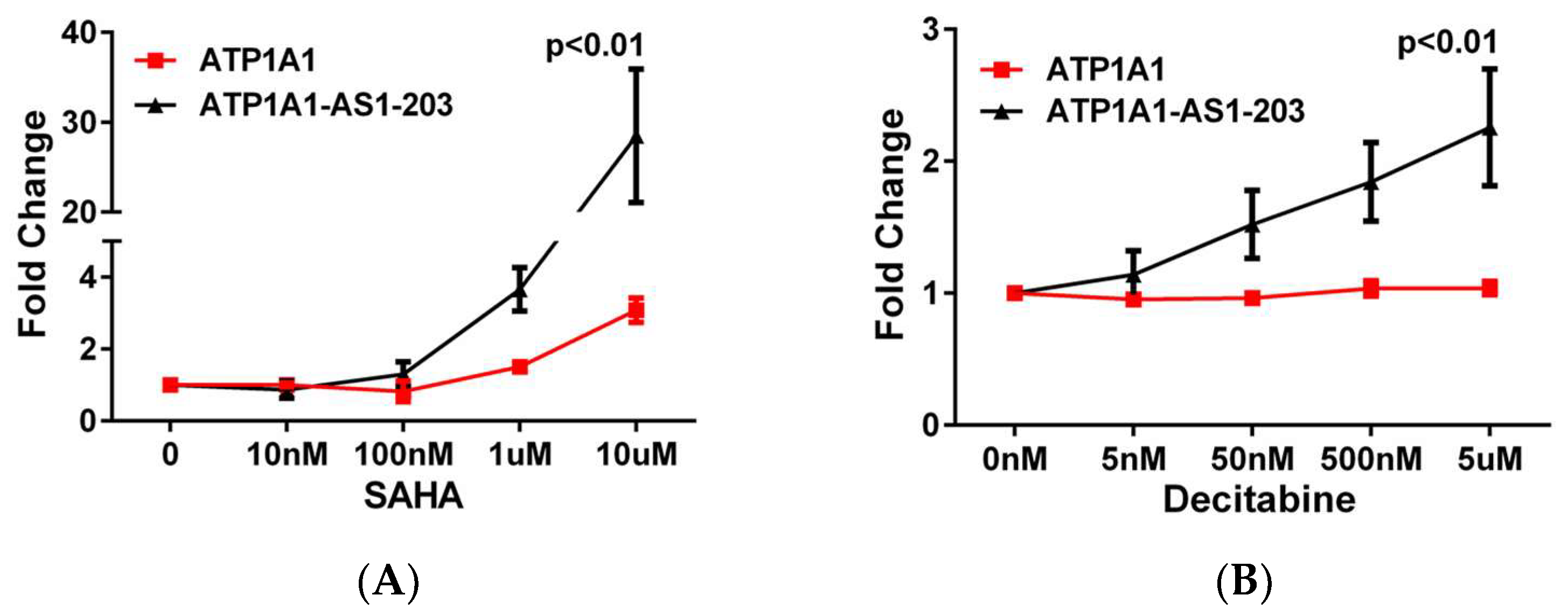
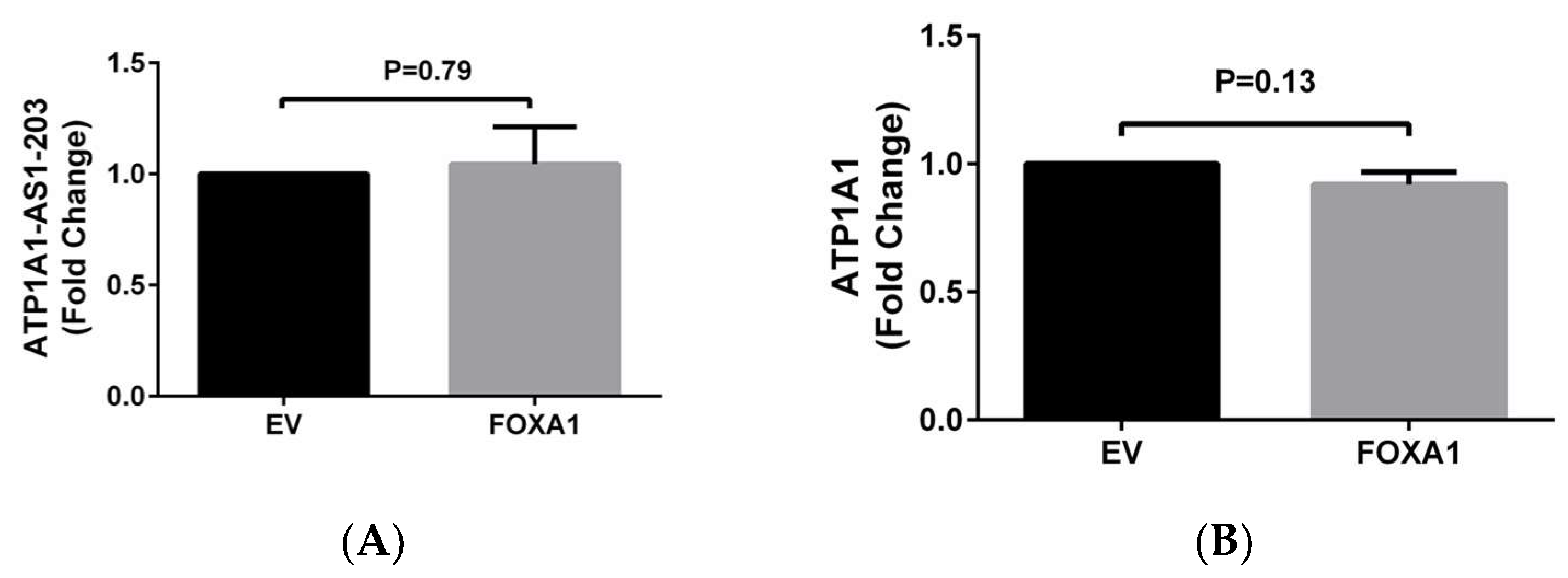
2.2. Epigenetic Regulation of ATP1A1-AS1 Expression
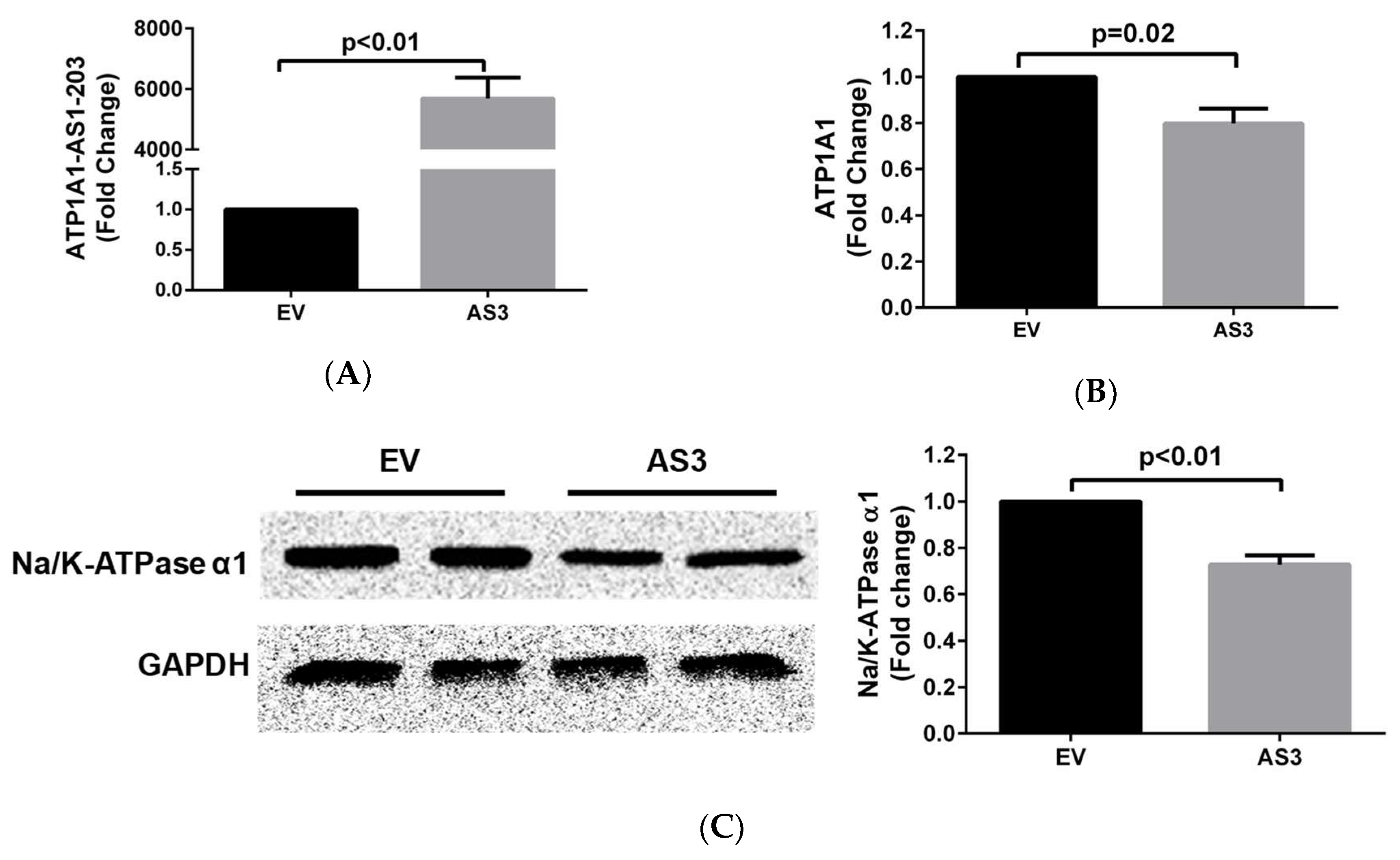
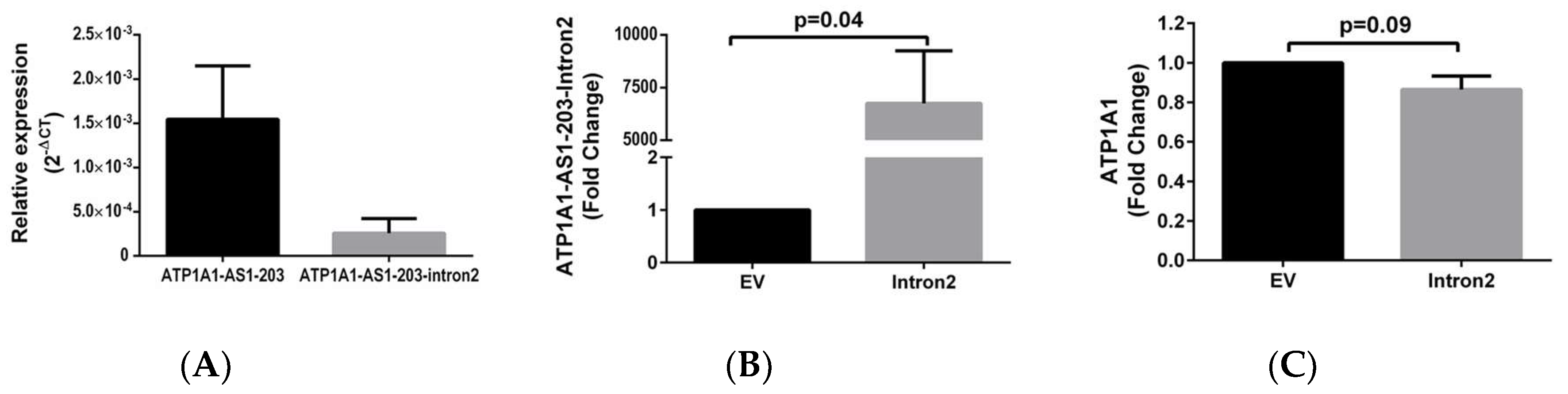
2.3. ATP1A1-AS1 Regulates the Sense Na/K-ATPase α1 Gene Expression and Protein Synthesis in HK2 Cells
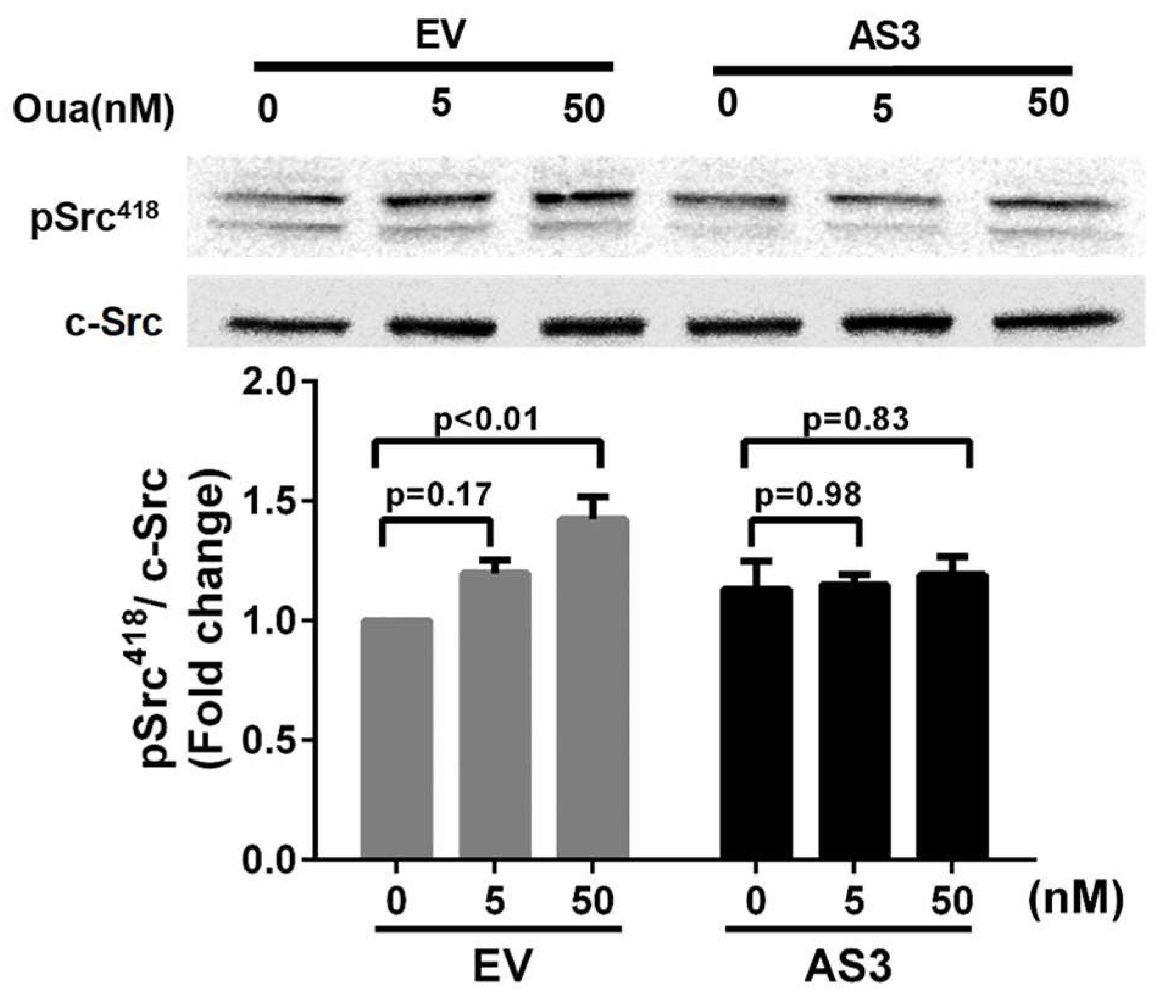
2.4. Overexpression of ATP1A1-AS1 Regulates Na/K-ATPase-Related Signaling and Cell Proliferation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Quantitation of ATP1A1-AS1 Expression Using RT-qPCR
4.3. Measurement of ATP1A1-AS1 Subcellular Distribution
4.4. Overexpression of ATP1A1-AS1 and FOXA1 in HK2 Cells
4.5. Western Blot
4.6. Cell Proliferation Assay
4.7. Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| lncRNAs | Long non-coding RNAs |
| SAHA | Suberoylanilide hydroxamic acid |
| HDAC | Histone deacetylase |
| CTS | Cardiotonic steroids |
References
- De Hoon, M.; Shin, J.W.; Carninci, P. Paradigm shifts in genomics through the fantom projects. Mamm. Genome 2015, 26, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hangauer, M.J.; Vaughn, I.W.; McManus, M.T. Pervasive transcription of the human genome produces thousands of previously unidentified long intergenic noncoding RNAs. PLoS Genet. 2013, 9, e1003569. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I.; Bartel, D.P. Lincrnas: Genomics, evolution, and mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Dykes, I.M.; Emanueli, C. Transcriptional and post-transcriptional gene regulation by long non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, D.; Nicolet, D.; Volinia, S.; Mrozek, K.; Yan, P.; Bundschuh, R.; Carroll, A.J.; Kohlschmidt, J.; Blum, W.; Powell, B.L.; et al. Prognostic and biologic significance of long non-coding RNA profiling in younger adults with cytogenetically normal acute myeloid leukemia. Haematologica 2017, 102, 1391–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlestedt, C. Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat. Rev. Drug Discov. 2013, 12, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Villegas, V.E.; Zaphiropoulos, P.G. Neighboring gene regulation by antisense long non-coding RNAs. Int. J. Mol. Sci. 2015, 16, 3251–3266. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Strausberg, R.L.; Feingold, E.A.; Grouse, L.H.; Derge, J.G.; Klausner, R.D.; Collins, F.S.; Wagner, L.; Shenmen, C.M.; Schuler, G.D.; Altschul, S.F.; et al. Generation and initial analysis of more than 15,000 full-length human and mouse cdna sequences. Proc. Natl. Acad. Sci. USA 2002, 99, 16899–16903. [Google Scholar] [PubMed]
- Duff, M.O.; Olson, S.; Wei, X.; Garrett, S.C.; Osman, A.; Bolisetty, M.; Plocik, A.; Celniker, S.E.; Graveley, B.R. Genome-wide identification of zero nucleotide recursive splicing in drosophila. Nature 2015, 521, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. MCP 2014, 13, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Skou, J.C. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim. Biophys. Acta 1957, 23, 394–401. [Google Scholar] [CrossRef]
- Aperia, A.; Akkuratov, E.E.; Fontana, J.M.; Brismar, H. Na+-K+-atpase, a new class of plasma membrane receptors. Am. J. Physiol. Cell Physiol. 2016, 310, C491–C495. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Askari, A. Na+/k+-atpase as a signal transducer. Eur. J. Biochem. 2002, 269, 2434–2439. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z. Molecular mechanisms of Na/k-atpase-mediated signal transduction. Ann. N.Y. Acad. Sci. 2003, 986, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Norgaard, A.; Bagger, J.P.; Bjerregaard, P.; Baandrup, U.; Kjeldsen, K.; Thomsen, P.E. Relation of left ventricular function and Na, k-pump concentration in suspected idiopathic dilated cardiomyopathy. Am. J. Cardiol. 1988, 61, 1312–1315. [Google Scholar] [CrossRef]
- Ishino, K.; Botker, H.E.; Clausen, T.; Hetzer, R.; Sehested, J. Myocardial adenine nucleotides, glycogen, and Na, k-atpase in patients with idiopathic dilated cardiomyopathy requiring mechanical circulatory support. Am. J. Cardiol. 1999, 83, 396–399. [Google Scholar] [CrossRef]
- Moseley, A.E.; Cougnon, M.H.; Grupp, I.L.; El Schultz, J.; Lingrel, J.B. Attenuation of cardiac contractility in Na, k-atpase α1 isoform-deficient hearts under reduced calcium conditions. J. Mol. Cell. Cardiol. 2004, 37, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Drummond, C.A.; Sayed, M.; Evans, K.L.; Shi, H.; Wang, X.; Haller, S.T.; Liu, J.; Cooper, C.J.; Xie, Z.; Shapiro, J.I.; et al. Reduction of Na/k-atpase affects cardiac remodeling and increases c-kit cell abundance in partial nephrectomized mice. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1631–H1643. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Bai, Y.; Chen, Y.; Wang, Y.; Sottejeau, Y.; Liu, L.; Li, X.; Lingrel, J.B.; Malhotra, D.; Cooper, C.J.; et al. Reduction of Na/k-atpase potentiates marinobufagenin-induced cardiac dysfunction and myocyte apoptosis. J. Biol. Chem. 2012, 287, 16390–16398. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Li, X.; Liang, M.; Liu, L.; Xie, J.X.; Ye, Q.; Kometiani, P.; Tillekeratne, M.; Jin, R.; Xie, Z. Changes in sodium pump expression dictate the effects of ouabain on cell growth. J. Biol. Chem. 2009, 284, 14921–14929. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, X.; Fu, C.; Yang, Q.; Xie, Y.; Zhang, Z.; Ye, Z. Impaired Na+-K+-atpase signaling in renal proximal tubule contributes to hyperuricemia-induced renal tubular injury. Exp. Mol. Med. 2018, 50, e452. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.X.; Li, X.; Xie, Z. Regulation of renal function and structure by the signaling Na/k-atpase. IUBMB Life 2013, 65, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Langhans, S.A. Transcriptional regulators of Na,k-atpase subunits. Front. Cell Dev. Biol. 2015, 3, 66. [Google Scholar] [CrossRef] [PubMed]
- Chernikova, D.; Managadze, D.; Glazko, G.V.; Makalowski, W.; Rogozin, I.B. Conservation of the exon-intron structure of long intergenic non-coding RNA genes in eutherian mammals. Life Basel 2016, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Chen, Y. Targeting long non-coding RNAs in cancers: Progress and prospects. Int. J. Biochem. Cell Boil. 2013, 45, 1895–1910. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Cai, T.; Tian, J.; Qu, W.; Xie, Z.J. Functional characterization of Src-interacting Na/k-atpase using RNA interference assay. J. Biol. Chem. 2006, 281, 19709–19719. [Google Scholar] [CrossRef] [PubMed]
- Semb, S.O.; Lunde, P.K.; Holt, E.; Tonnessen, T.; Christensen, G.; Sejersted, O.M. Reduced myocardial Na+, k+-pump capacity in congestive heart failure following myocardial infarction in rats. J. Mol. Cell. Cardiol. 1998, 30, 1311–1328. [Google Scholar] [CrossRef] [PubMed]
- El-Mallakh, R.S.; Barrett, J.L.; Jed Wyatt, R. The Na, K-atpase hypothesis for bipolar disorder: Implications of normal development. J. Child Adolesc. Psychopharmacol. 1993, 3, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Poehlman, E.T. Regulation of energy expenditure in aging humans. J. Am. Geriatr. Soc. 1993, 41, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Maurya, P.K.; Prakash, S. Decreased activity of Ca++-atpase and Na+/K+-atpase during aging in humans. Appl. Biochem. Biotechnol. 2013, 170, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Tirupattur, P.R.; Ram, J.L.; Standley, P.R.; Sowers, J.R. Regulation of Na+, K+-atpase gene expression by insulin in vascular smooth muscle cells. Am. J. Hypertens. 1993, 6, 626–629. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yuan, C.; Clough, D.; Schooley, J.; Haddy, F.J.; Pamnani, M.B. Role of digitalis-like substance in the hypertension of streptozotocin-induced diabetes in reduced renal mass rats. Am. J. Hypertens. 1993, 6, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Clerico, A.; Giampietro, O. Is the endogenous digitalis-like factor the link between hypertension and metabolic disorders as diabetes mellitus, obesity and acromegaly? Clin. Physiol. Biochem. 1990, 8, 153–168. [Google Scholar] [PubMed]
- Harik, S.I.; Mitchell, M.J.; Kalaria, R.N. Ouabain binding in the human brain: Effects of Alzheimer’s disease and aging. Arch. Neurol. 1989, 46, 951–954. [Google Scholar] [CrossRef] [PubMed]
- Liguri, G.; Taddei, N.; Nassi, P.; Latorraca, S.; Nediani, C.; Sorbi, S. Changes in Na+, K+-atpase, Ca2+-atpase and some soluble enzymes related to energy metabolism in brains of patients with Alzheimer’s disease. Neurosci. Lett. 1990, 112, 338–342. [Google Scholar] [CrossRef]
- Munroe, S.H.; Zhu, J. Overlapping transcripts, double-stranded RNA and antisense regulation: A genomic perspective. Cell Mol. Life Sci. 2006, 63, 2102–2118. [Google Scholar] [CrossRef] [PubMed]
- Faghihi, M.A.; Wahlestedt, C. Regulatory roles of natural antisense transcripts. Nat. Rev. Mol. Cell Biol. 2009, 10, 637–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Ramchandran, R. Natural antisense transcript: A concomitant engagement with protein-coding transcript. Oncotarget 2010, 1, 447–452. [Google Scholar] [PubMed]
- Wight, M.; Werner, A. The functions of natural antisense transcripts. Essays Biochem. 2013, 54, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksen, C.; Kjaer-Sorensen, K.; Einholm, A.P.; Madsen, L.B.; Momeni, J.; Bendixen, C.; Oxvig, C.; Vilsen, B.; Larsen, K. Molecular cloning and characterization of porcine Na+/K+-atpase isoforms α1, α2, α3 and the atp1a3 promoter. PLoS ONE 2013, 8, e79127. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, P.; Owens, T.A.; David, J.M.; Petrelli, N.J.; Christensen, B.C.; Lakshmikuttyamma, A.; Rajasekaran, A.K. Epigenetic silencing of Na, K-atpase β1 subunit gene atp1b1 by methylation in clear cell renal cell carcinoma. Epigenetics 2014, 9, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Trcek, T.; Wells, A.L.; Chao, J.A.; Singer, R.H. An unbiased analysis method to quantify mRNA localization reveals its correlation with cell motility. Cell Rep. 2012, 1, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Berteaux, N.; Aptel, N.; Cathala, G.; Genton, C.; Coll, J.; Daccache, A.; Spruyt, N.; Hondermarck, H.; Dugimont, T.; Curgy, J.J.; et al. A novel h19 antisense RNA overexpressed in breast cancer contributes to paternal igf2 expression. Mol. Cell. Biol. 2008, 28, 6731–6745. [Google Scholar] [CrossRef] [PubMed]
- Kumarasamy, S.; Waghulde, H.; Gopalakrishnan, K.; Mell, B.; Morgan, E.; Joe, B. Mutation within the hinge region of the transcription factor Nr2f2 attenuates salt-sensitive hypertension. Nat. Commun. 2015, 6, 6252. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Ashraf, U.M.; Drummond, C.A.; Shi, H.; Zhang, X.; Kumarasamy, S.; Tian, J. Characterization of a Long Non-Coding RNA, the Antisense RNA of Na/K-ATPase α1 in Human Kidney Cells. Int. J. Mol. Sci. 2018, 19, 2123. https://doi.org/10.3390/ijms19072123
Fan X, Ashraf UM, Drummond CA, Shi H, Zhang X, Kumarasamy S, Tian J. Characterization of a Long Non-Coding RNA, the Antisense RNA of Na/K-ATPase α1 in Human Kidney Cells. International Journal of Molecular Sciences. 2018; 19(7):2123. https://doi.org/10.3390/ijms19072123
Chicago/Turabian StyleFan, Xiaoming, Usman M. Ashraf, Christopher A. Drummond, Huilin Shi, Xiaolu Zhang, Sivarajan Kumarasamy, and Jiang Tian. 2018. "Characterization of a Long Non-Coding RNA, the Antisense RNA of Na/K-ATPase α1 in Human Kidney Cells" International Journal of Molecular Sciences 19, no. 7: 2123. https://doi.org/10.3390/ijms19072123
APA StyleFan, X., Ashraf, U. M., Drummond, C. A., Shi, H., Zhang, X., Kumarasamy, S., & Tian, J. (2018). Characterization of a Long Non-Coding RNA, the Antisense RNA of Na/K-ATPase α1 in Human Kidney Cells. International Journal of Molecular Sciences, 19(7), 2123. https://doi.org/10.3390/ijms19072123