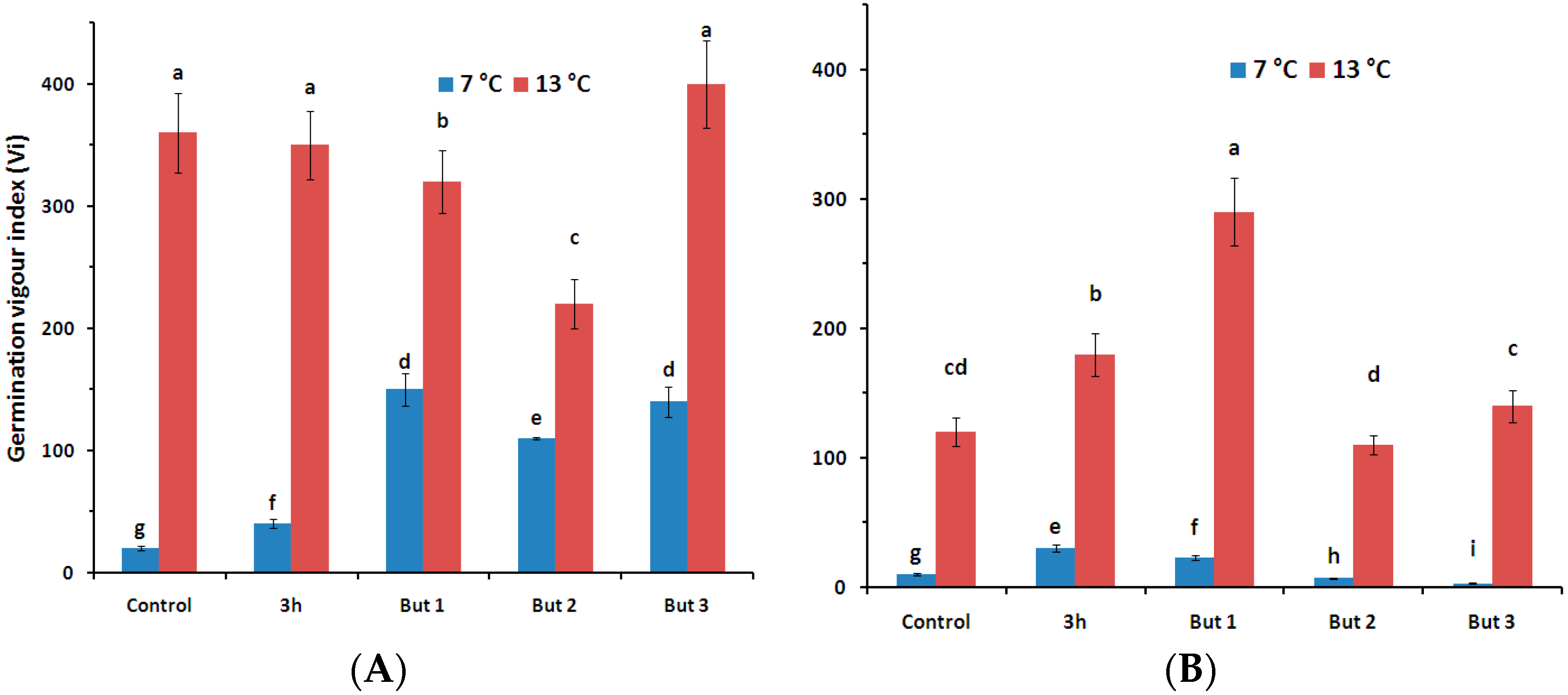
Figure 1.
Germination vigour index (Vi) of seeds of L. angustifolius cultivars (Lazur—(A) and Sethes Fruehe Rote—(B)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Values represent means (n = 5) ± SE. Different letters over error bars (a, b, c, …) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 1.
Germination vigour index (Vi) of seeds of L. angustifolius cultivars (Lazur—(A) and Sethes Fruehe Rote—(B)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Values represent means (n = 5) ± SE. Different letters over error bars (a, b, c, …) indicate significant differences (Duncan’s multiple range test, p < 0.05).
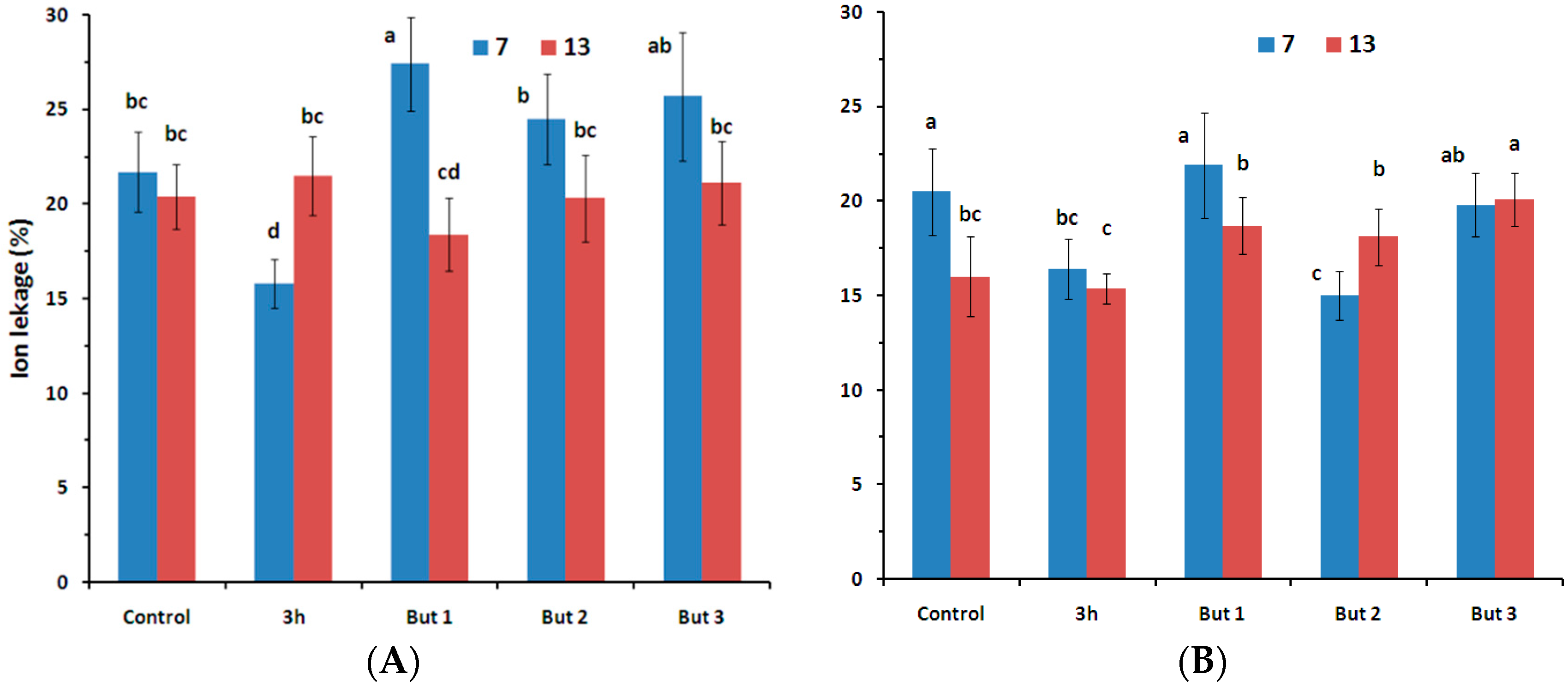
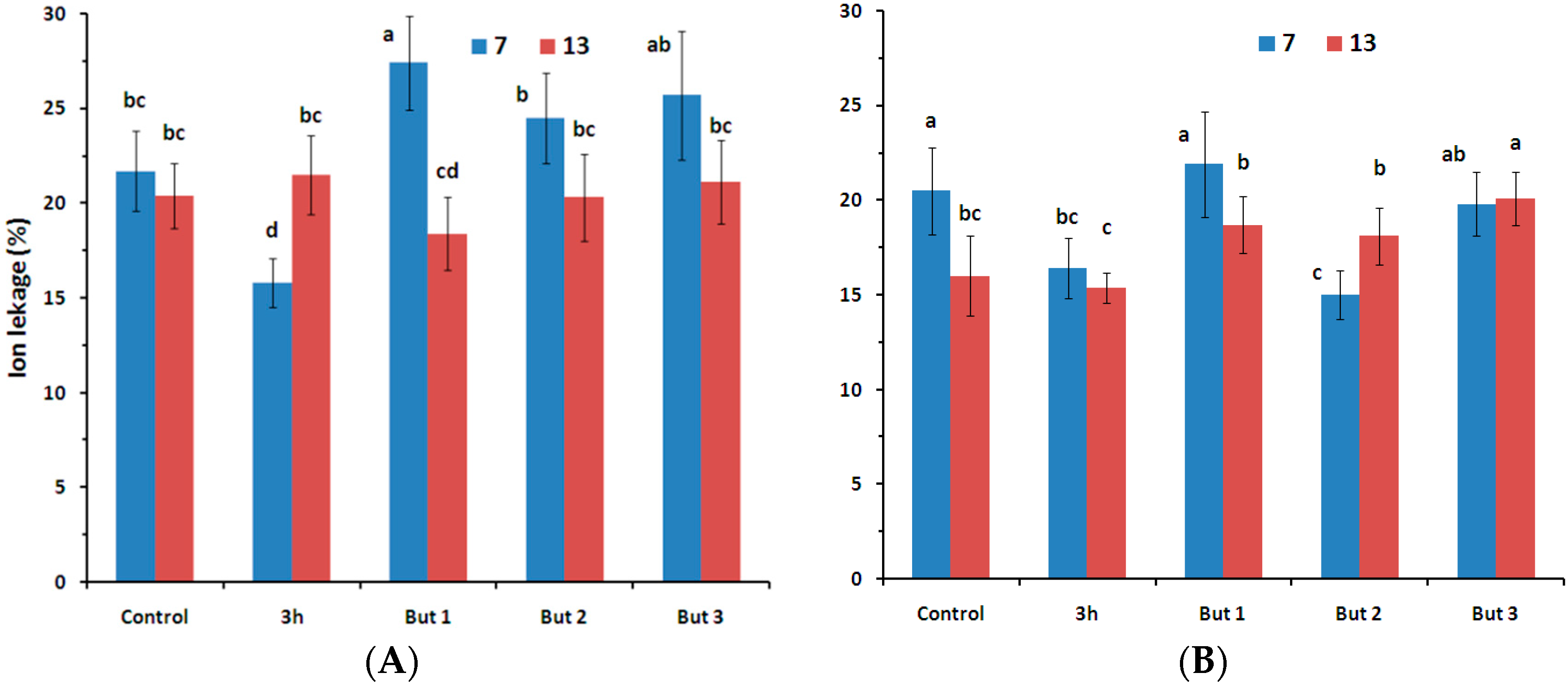
Figure 2.
Percentage ion leakage from seeds of two L. angustifolius cultivars (Lazur—(A); and Sethes Fruehe Rote—(B)) germinated at 7 °C (blue bars) or 13 °C (red bars) in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Letters over error bars (±SE; n = 10) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 2.
Percentage ion leakage from seeds of two L. angustifolius cultivars (Lazur—(A); and Sethes Fruehe Rote—(B)) germinated at 7 °C (blue bars) or 13 °C (red bars) in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Letters over error bars (±SE; n = 10) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 3.
Dehydrogenase activity (DA) [μg formazan·g−1 protein·min−1] in the seeds of two L. angustifolius Lazur cultivars (Lazur—(A); and Sethes Fruehe Rote—(B)) germinating at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Different letters over error bars (±SE; n = 5) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 3.
Dehydrogenase activity (DA) [μg formazan·g−1 protein·min−1] in the seeds of two L. angustifolius Lazur cultivars (Lazur—(A); and Sethes Fruehe Rote—(B)) germinating at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Different letters over error bars (±SE; n = 5) indicate significant differences (Duncan’s multiple range test, p < 0.05).
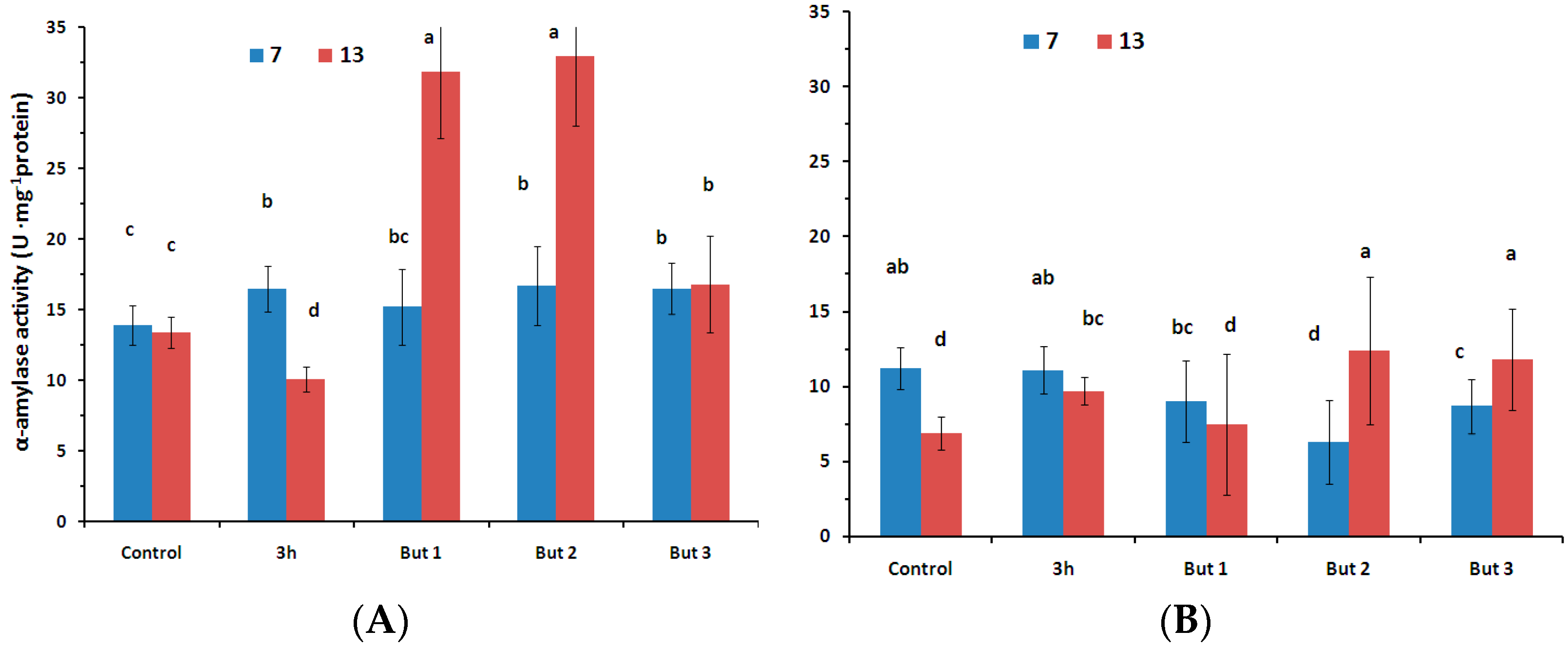
Figure 4.
Activity of α-amylase [U·mg−1 protein] in the seeds of two L. angustifolius cultivars (Lazur—(A); and Sethes Fruehe Rote—(B)) germinating at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 4.
Activity of α-amylase [U·mg−1 protein] in the seeds of two L. angustifolius cultivars (Lazur—(A); and Sethes Fruehe Rote—(B)) germinating at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide, respectively. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
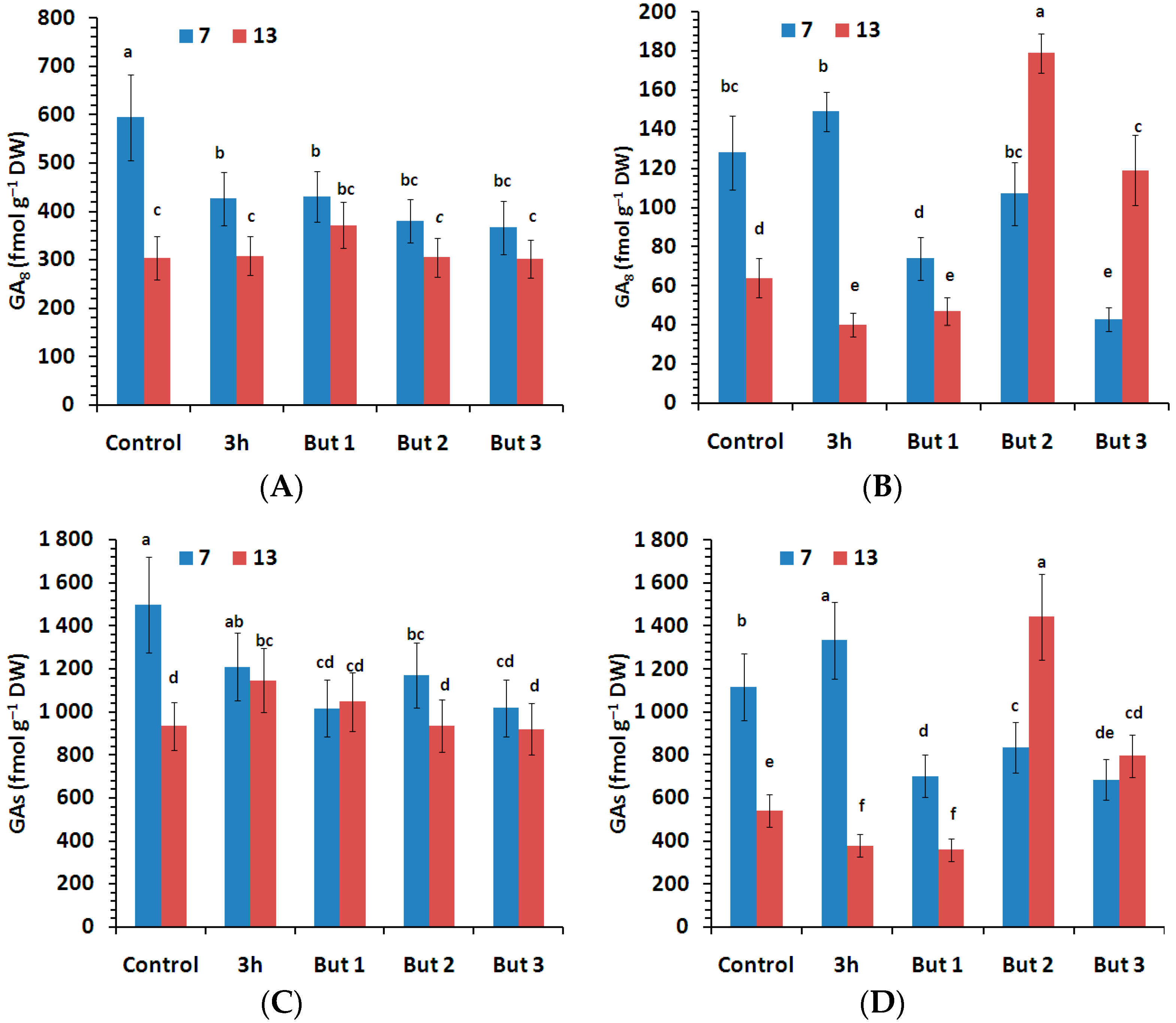
Figure 5.
Content of non-active form of gibberellins (GA8) and sum of active gibberellins (GA1, GA3, GA4, GA5, GA6) measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C); Sethes Fruehe Rote—(B,D)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 5.
Content of non-active form of gibberellins (GA8) and sum of active gibberellins (GA1, GA3, GA4, GA5, GA6) measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C); Sethes Fruehe Rote—(B,D)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
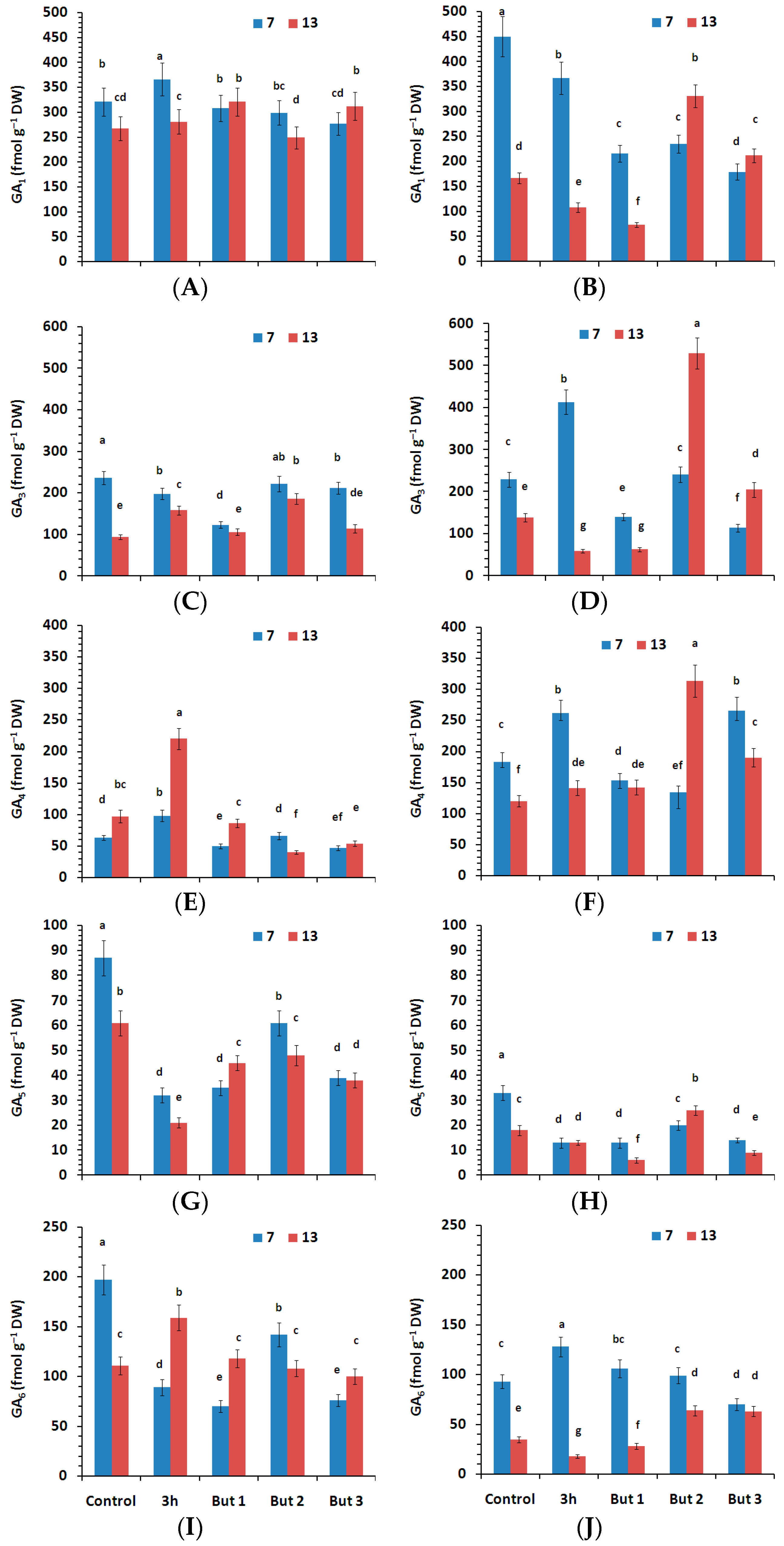
Figure 6.
Content of active gibberellins (GA1, GA3, GA4, GA5, GA6) measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C,E,G,I); Sethes Fruehe Rote—(B,D,F,H,J)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 6.
Content of active gibberellins (GA1, GA3, GA4, GA5, GA6) measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C,E,G,I); Sethes Fruehe Rote—(B,D,F,H,J)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
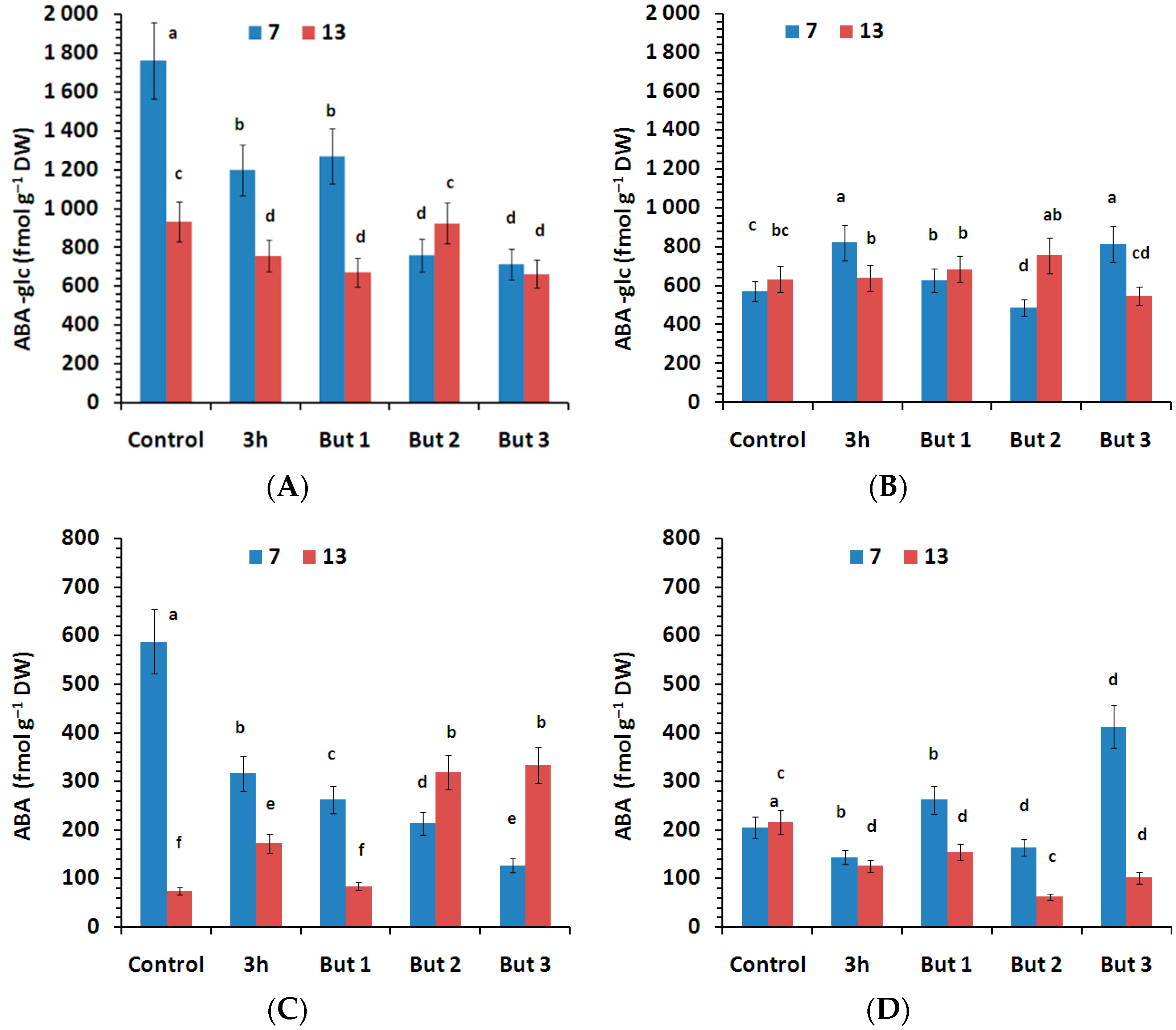
Figure 7.
Content of inactivated form of ABA (ABA-glc) and active ABA measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C); Sethes Fruehe Rote—(B,D)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 7.
Content of inactivated form of ABA (ABA-glc) and active ABA measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C); Sethes Fruehe Rote—(B,D)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
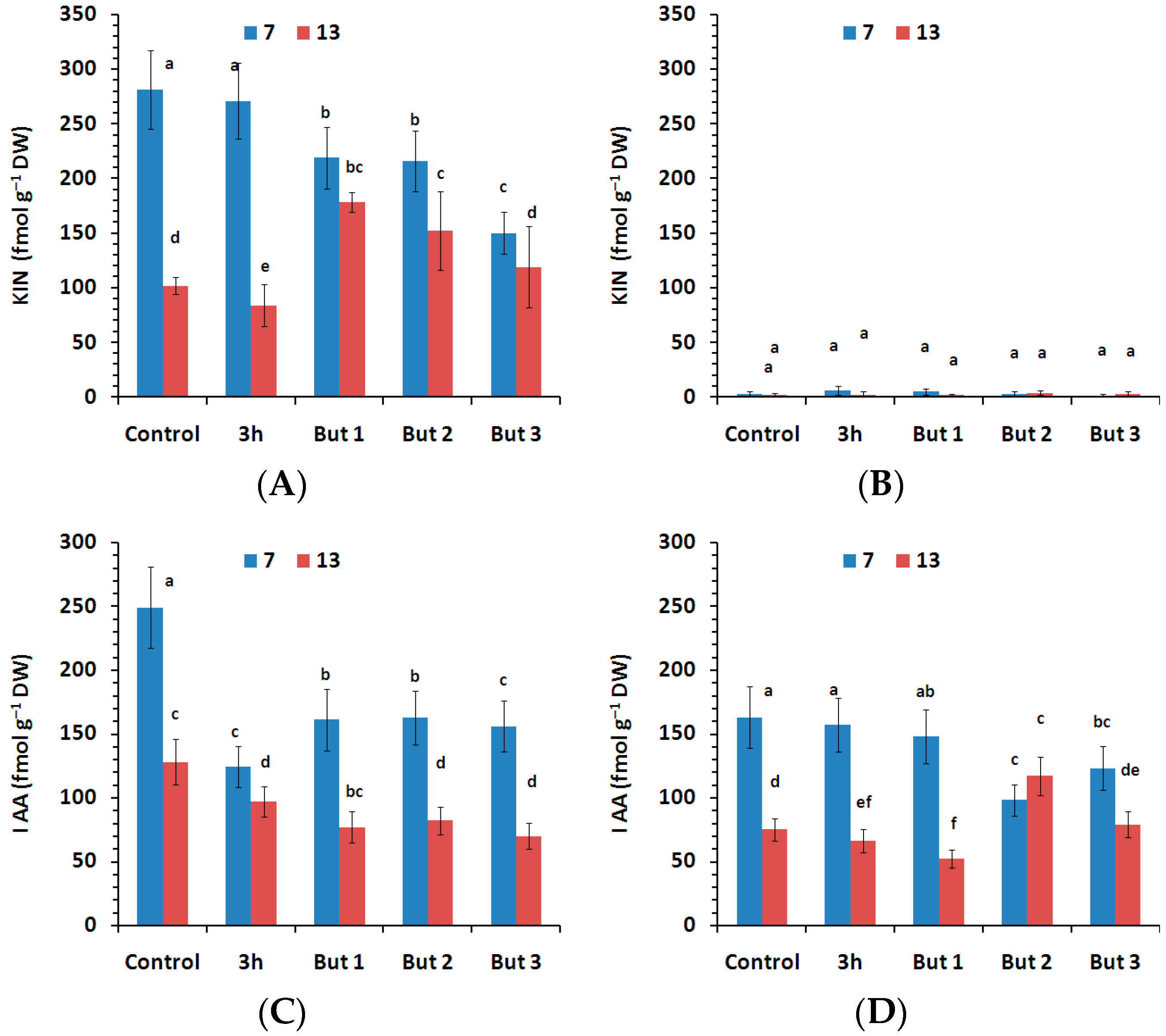
Figure 8.
Content of KIN—kinetin abd IAA—indolile-3-acetic acid measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C); Sethes Fruehe Rote—(B,D)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 8.
Content of KIN—kinetin abd IAA—indolile-3-acetic acid measured in the seeds of two L. angustifolius cultivars (Lazur—(A,C); Sethes Fruehe Rote—(B,D)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
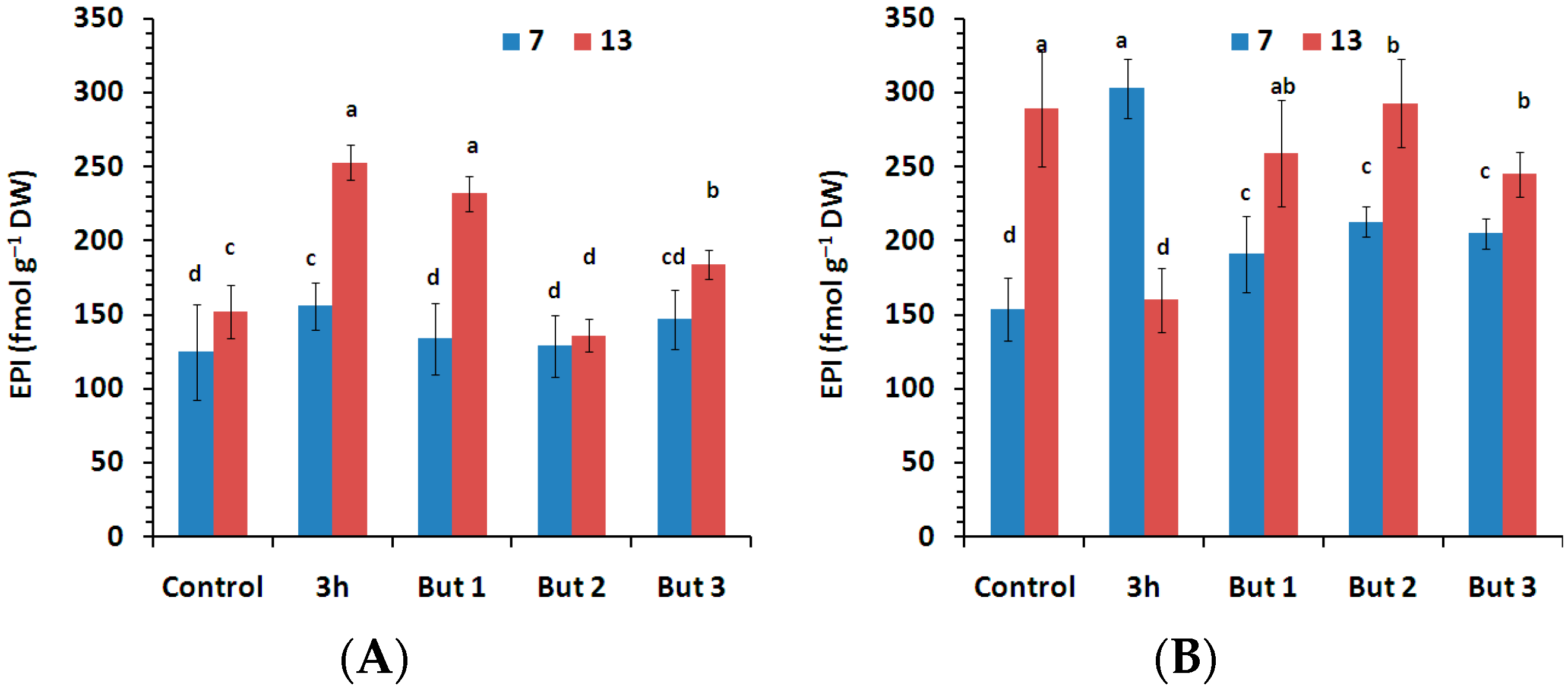
Figure 9.
Content of EPI—24-epibrassinolide measured in the seeds of two L. angustifolius cultivars (Lazur—(A); Sethes Fruehe Rote—(B)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
Figure 9.
Content of EPI—24-epibrassinolide measured in the seeds of two L. angustifolius cultivars (Lazur—(A); Sethes Fruehe Rote—(B)) germinated at 7 °C or 13 °C in water (control), butenolide or hydroprimed in water for 3 h at 20 °C. But 1, But 2, But 3: 10−6 M, 10−5 M, 10−4 M butenolide concentrations. Different letters over error bars (±SE; n = 3) indicate significant differences (Duncan’s multiple range test, p < 0.05).
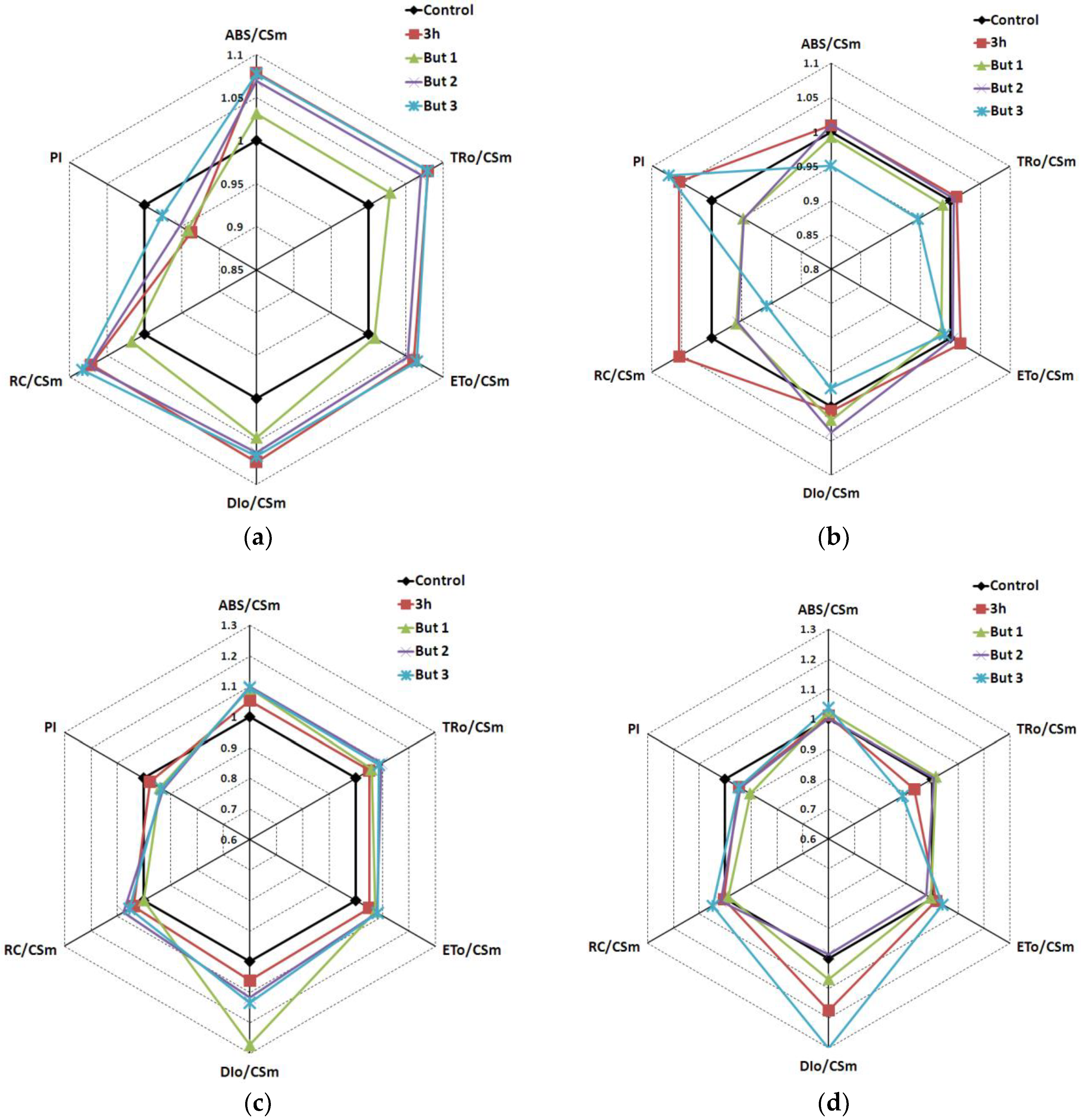
Figure 10.
Radar plot of chlorophyll a fluorescence kinetics parameters of two L. angustifolius cultivars germinated and grown for the first four weeks at 7 °C (Lazur—(a); Sethes Fruehe Rote—(b)) or 13 °C (Lazur—(c); Sethes Fruehe Rote—(d)). Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6, 10−5 and 10−4 M butenolide, respectively. Black line indicates to control plants as 100%. Values represent means (n = 10).
Figure 10.
Radar plot of chlorophyll a fluorescence kinetics parameters of two L. angustifolius cultivars germinated and grown for the first four weeks at 7 °C (Lazur—(a); Sethes Fruehe Rote—(b)) or 13 °C (Lazur—(c); Sethes Fruehe Rote—(d)). Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6, 10−5 and 10−4 M butenolide, respectively. Black line indicates to control plants as 100%. Values represent means (n = 10).
Table 1.
Effects of temperature and seed treatment (two-way ANOVA) on germination vigour index (Vi), ion leakage (EL), dehydrogenase (DA) and α-amylase (A) activity, content of inactive gibberellin (GA8), active gibberellins (GAs), conjugated abscisic acid (ABA-glc), free ABA, kinetin (KIN), indole-3-acetic acid (IAA) and 24-epibrassinolide (EPI) in the seeds of two L. angustifolius cultivars germinated at 7 °C or 13 °C in water or after pre-sowing treatments: Three-hour hydropriming or 10−6 M, 10−5 M, 10−4 M butenolide.
Table 1.
Effects of temperature and seed treatment (two-way ANOVA) on germination vigour index (Vi), ion leakage (EL), dehydrogenase (DA) and α-amylase (A) activity, content of inactive gibberellin (GA8), active gibberellins (GAs), conjugated abscisic acid (ABA-glc), free ABA, kinetin (KIN), indole-3-acetic acid (IAA) and 24-epibrassinolide (EPI) in the seeds of two L. angustifolius cultivars germinated at 7 °C or 13 °C in water or after pre-sowing treatments: Three-hour hydropriming or 10−6 M, 10−5 M, 10−4 M butenolide.
| Source of Variance | df | Vi | EL | DA | A | GA8 | GAs | ABA-Glu | Free ABA | KIN | IAA | EPI |
|---|
| Cultivar (C) | 1 | * | *** | * | * | *** | *** | ** | ** | *** | * | * |
| Temperature (t) | 1 | ** | * | ** | ns | *** | *** | * | * | ** | ** | * |
| Treatment (T) | 4 | * | ns | *** | ns | * | * | * | * | * | ns | * |
| C × t | 1 | ns | ns | ns | ns | ** | ** | ns | ns | ns | ns | ns |
| C × T | 1 | ns | ns | ns | ns | ** | ** | ns | ns | ns | ns | ns |
| C × t × T | 4 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
Table 2.
Effects of temperature and pre-sowing seed treatment (two-way ANOVA) on vegetative phase development index (VPDi), fresh (FW) and dry weight (DW) of shoots and generative development (GD) of two L. angustifolius cultivars grown for the first four weeks at 7 °C or 13 °C from seeds three-hour hydroprimed or treated with 10−6 M, 10−5 M or 10−4 M butenolide.
Table 2.
Effects of temperature and pre-sowing seed treatment (two-way ANOVA) on vegetative phase development index (VPDi), fresh (FW) and dry weight (DW) of shoots and generative development (GD) of two L. angustifolius cultivars grown for the first four weeks at 7 °C or 13 °C from seeds three-hour hydroprimed or treated with 10−6 M, 10−5 M or 10−4 M butenolide.
| Source of Variance | df | VPDi | FW | DW | GD |
|---|
| Cultivar (C) | 1 | ** | ns | ns | *** |
| Temperature (t) | 1 | *** | *** | *** | ** |
| Treatment (T) | 4 | ** | * | ns | ** |
| C × t | 1 | * | ** | ** | * |
| C × T | 1 | * | * | * | * |
| C × t × T | 4 | ns | * | * | * |
Table 3.
Effects of pre-sowing treatments on the index of vegetative phase development (VPDi) of two L. angustifolius cultivars germinated and grown for the first four weeks at 7 °C or 13 °C. Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
Table 3.
Effects of pre-sowing treatments on the index of vegetative phase development (VPDi) of two L. angustifolius cultivars germinated and grown for the first four weeks at 7 °C or 13 °C. Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
| Temperature (°C) | Treatment | Lazur | Sethes Fruehe Rote |
|---|
| 7 | Control | 251 ± 22c | 138 ± 11f |
| | 3 h | 250 ± 21c | 142 ± 12ef |
| | But 1 | 141 ± 15e | 72 ± 6g |
| | But 2 | 150 ± 16e | 52 ± 5h |
| | But 3 | 190 ± 20d | 156 ± 17e |
| 13 | Control | 356 ± 32a | 252 ± 23c |
| | 3 h | 360 ± 33a | 208 ± 19d |
| | But 1 | 352 ± 31a | 311 ± 29b |
| | But 2 | 312 ± 29b | 413 ± 39a |
| | But 3 | 324 ± 30ab | 308 ± 27b |
Table 4.
Effects of pre-sowing treatments on fresh (FW) and dry weight (DW) [g] of two L. angustifolius cultivars germinated and grown for the first four weeks at 7 °C or 13 °C. Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
Table 4.
Effects of pre-sowing treatments on fresh (FW) and dry weight (DW) [g] of two L. angustifolius cultivars germinated and grown for the first four weeks at 7 °C or 13 °C. Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
| Temperature (°C) | Treatment | Lazur | Sethes Fruehe Rote |
|---|
| FW | DW | FW | DW |
|---|
| 7 | Control | 3.54 ± 0.42d | 0.559 ± 0.071e | 5.15 ± 0.61b | 0.748 ± 0.121c |
| | 3 h | 4.31 ± 0.51c | 0.691 ± 0.093cd | 4.90 ± 0.58bc | 0.757 ± 0.122c |
| | But 1 | 3.65 ± 0.43d | 0.575 ± 0.072e | 4.59 ± 0.60c | 0.599 ± 0.077d |
| | But 2 | 4.34 ± 0.52c | 0.714 ± 0.093bc | 3.10 ± 0.37e | 0.612 ± 0.073d |
| | But 3 | 3.33 ± 0.40d | 0.546 ± 0.071e | 3.55 ± 0.42d | 0.591 ± 0.071d |
| 13 | Control | 4.48 ± 0.49c | 0.774 ± 0.100b | 7.09 ± 0.85a | 1.016 ± 0.122a |
| | 3 h | 5.91 ± 0.70a | 0.934 ± 0.112a | 4.11 ± 0.50c | 0.959 ± 0.116ab |
| | But 1 | 4.71 ± 0.52c | 0.774 ± 0.100b | 4.52 ± 0.54c | 1.006 ± 0.141a |
| | But 2 | 5.22 ± 0.63b | 0.850 ± 0.110ab | 3.86 ± 0.46d | 1.006 ± 0.141a |
| | But 3 | 3.93 ± 0.43d | 0.605 ± 0.071d | 5.56 ± 0.67b | 0.877 ± 0.123b |
Table 5.
Effects of temperature and pre-sowing seed treatment (two-way ANOVA) on the kinetics of chlorophyll a fluorescence in plants of two L. angustifolius cultivars grown at 7 °C and 13 °C for the first four weeks from seeds germinated in water, three-hour hydroprimed or treated with 10−6 M, 10−5 M, 10−4 M butenolide.
Table 5.
Effects of temperature and pre-sowing seed treatment (two-way ANOVA) on the kinetics of chlorophyll a fluorescence in plants of two L. angustifolius cultivars grown at 7 °C and 13 °C for the first four weeks from seeds germinated in water, three-hour hydroprimed or treated with 10−6 M, 10−5 M, 10−4 M butenolide.
| Source of Variance | df | ABS/CSm | TRo/CSm | ETo/CSm | DIo/CSm | RC/CSm | PI |
|---|
| Cultivar (C) | 1 | ** | *** | ** | ** | ns | *** |
| Temperature (t) | 1 | ** | *** | *** | * | ns | *** |
| Treatment (T) | 4 | ns | ns | ns | * | * | ** |
| C × t | 1 | ns | * | ns | ns | * | ** |
| C × T | 1 | ns | ns | * | ns | ** | ns |
| C × t × T | 4 | * | ** | ns | * | ** | ** |
Table 6.
Effects of pre-sowing treatments on generative development (GD) of two L. angustifolius cultivars germinated and grown for the first four week at 7 °C or 13 °C. Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
Table 6.
Effects of pre-sowing treatments on generative development (GD) of two L. angustifolius cultivars germinated and grown for the first four week at 7 °C or 13 °C. Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in the presence of 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
| Temperature (°C) | Treatment | Lazur | Sethes Fruehe Rote |
|---|
| 7 | Control | 1.79 ± 0.13c | 0.69 ± 0.05b |
| 3 h | 2.00 ± 0.16a | 0.77 ± 0.06b |
| But 1 | 1.58 ± 0.12d | 0.31 ± 0.02c |
| But 2 | 1.50 ± 0.12d | 0.38 ± 0.03c |
| But 3 | 1.48 ± 0.11d | 0.06 ± 0.01d |
| 13 | Control | 1.83 ± 0.15bc | 0.80 ± 0.07a |
| 3 h | 2.00 ± 0.16a | 0.90 ± 0.08a |
| But 1 | 1.50 ± 0.12d | 0.83 ± 0.07a |
| But 2 | 1.92 ± 0.14ab | 0.67 ± 0.05b |
| But 3 | 1.75 ± 0.14c | 0.33 ± 0.03c |
Table 7.
Effects of temperature and pre-sowing seed treatment on the number of pods, seeds and seed weight per plant of two L. angustifolius cultivars grown for the first four weeks at 7 °C or 13 °C from seeds germinated in water, three-hour hydroprimed or treated with 10−6 M, 10−5 M, 10−4 M butenolide.
Table 7.
Effects of temperature and pre-sowing seed treatment on the number of pods, seeds and seed weight per plant of two L. angustifolius cultivars grown for the first four weeks at 7 °C or 13 °C from seeds germinated in water, three-hour hydroprimed or treated with 10−6 M, 10−5 M, 10−4 M butenolide.
| Source of Variance | df | No. of Pods | No. of Seeds | Seed Weight |
|---|
| Cultivar (C) | 1 | *** | *** | * |
| Temperature (t) | 1 | *** | * | ns |
| Treatment (T) | 4 | *** | *** | *** |
| C × t | 1 | *** | *** | * |
| C × T | 1 | ** | *** | *** |
| C × t × T | 4 | * | *** | *** |
Table 8.
Effects of pre-sowing treatments on number of pods, seeds and seed weight (g) per plant of two L. angustifolius cultivars germinated and grown for the first 4 weeks at 7 °C (cold) or 13 °C (control temperature). Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
Table 8.
Effects of pre-sowing treatments on number of pods, seeds and seed weight (g) per plant of two L. angustifolius cultivars germinated and grown for the first 4 weeks at 7 °C (cold) or 13 °C (control temperature). Control—plants grown from seeds germinated in water; 3 h—plants grown from seeds hydroprimed for 3 h at 20 °C; But 1, But 2, But 3—plants grown form seeds germinated in 10−6 M, 10−5 M, 10−4 M butenolide, respectively.
| Temperature (°C) | Treatment | Lazur | Sethes Fruehe Rote |
|---|
| No. of Pods | No. of Seeds | Seed Weight | No. of Pods | No. of Seeds | Seed Weight |
|---|
| 7 | Control | 6.3 ± 0.4a | 15.7 ± 1.2b | 1.28 ± 0.05c | 3.3 ± 0.2b | 12.9 ± 0.9bc | 1.89 ± 0.17b |
| | 3 h | 6.3 ± 0.6a | 18.6 ± 1.0a | 1.69 ± 0.16ab | 3.2 ± 0.3b | 13.4 ± 1.2b | 1.91 ± 0.14b |
| | But 1 | 5.1 ± 0.4bc | 13.8 ± 1.0c | 1.21 ± 0.09c | 3.6 ± 0.5b | 13.3 ± 2.0b | 1.77 ± 0.28bc |
| | But 2 | 5.3 ± 0.7bc | 19.0 ± 2.3a | 1.74 ± 0.27a | 3.1 ± 0.4b | 9.1 ± 0.9d | 1.31 ± 0.15e |
| | But 3 | 4.8 ± 0.4c | 13.2 ± 1.2c | 1.15 ± 0.07d | 1.7 ± 0.2d | 5.6 ± 0.6e | 0.72 ± 0.11g |
| 13 | Control | 5.8 ± 0.4ab | 11.8 ± 1.4d | 1.29 ± 0.13c | 3.6 ± 0.4b | 12.2 ± 1.5c | 1.69 ± 0.32c |
| | 3 h | 6.6 ± 0.7a | 15.8 ± 1.3b | 1.55 ± 0.10b | 4.7 ± 0.3a | 17.0 ± 1.4a | 2.28 ± 0.29a |
| | But 1 | 3.9 ± 0.6d | 9.0 ± 1.4d | 0.86 ± 0.13e | 3.0 ± 0.2b | 11.0 ± 0.6c | 1.49 ± 0.08d |
| | But 2 | 6.1 ± 0.5a | 13.9 ± 0.9c | 1.48 ± 0.12b | 2.7 ± 0.2c | 9.6 ± 0.8d | 1.17 ± 0.12f |
| | But 3 | 5.2 ± 0.3bc | 14.2 ± 1.7bc | 1.29 ± 0.17c | 2.6 ± 0.2c | 9.2 ± 1.0d | 0.77 ± 0.12g |

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









