The Effect of Pectinase-Assisted Extraction on the Physicochemical and Biological Properties of Polysaccharides from Aster scaber

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Preparation of the Extracts
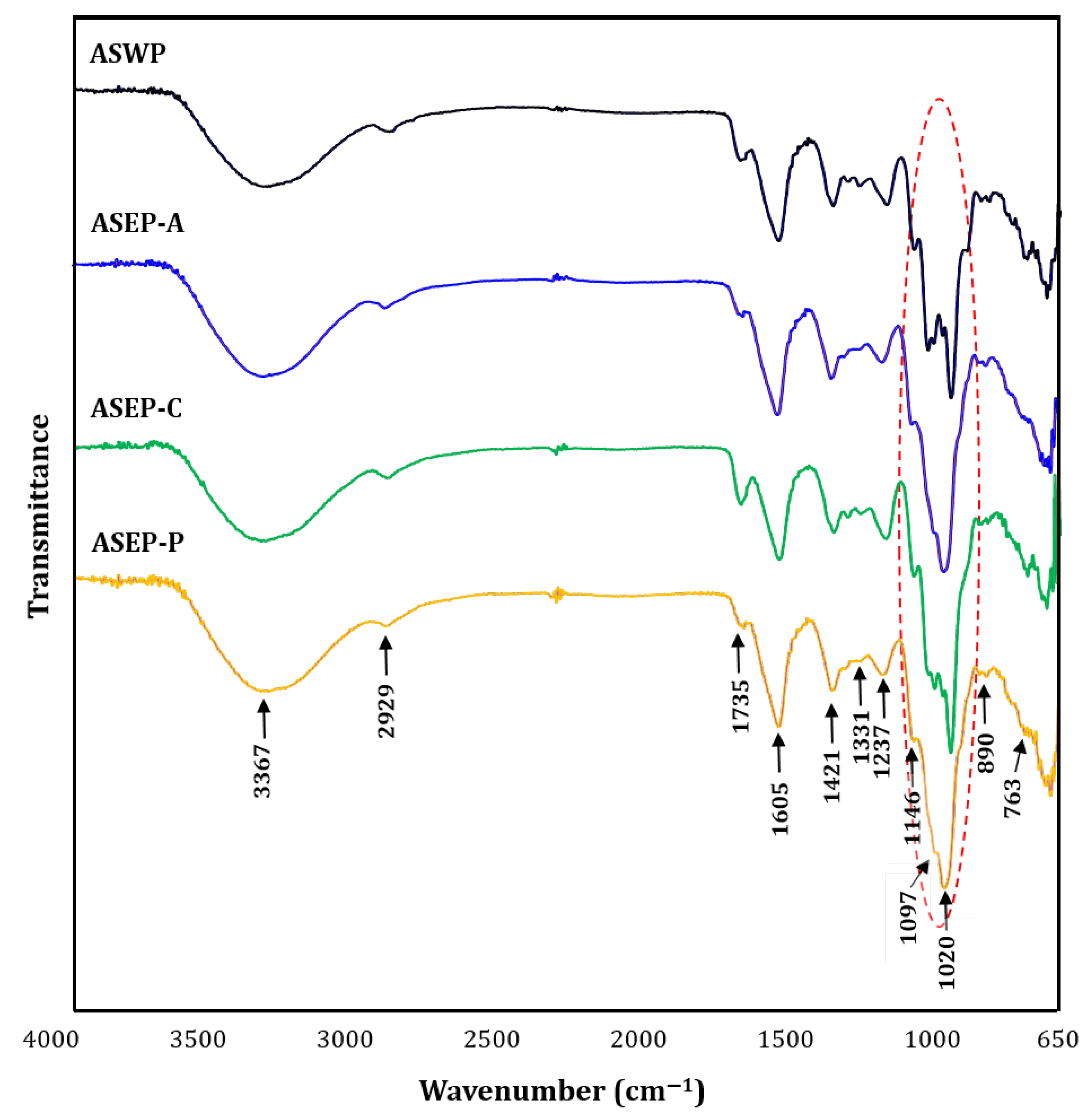
2.2. Comparison of Physicochemical Properties of ASWP and Three ASEPs
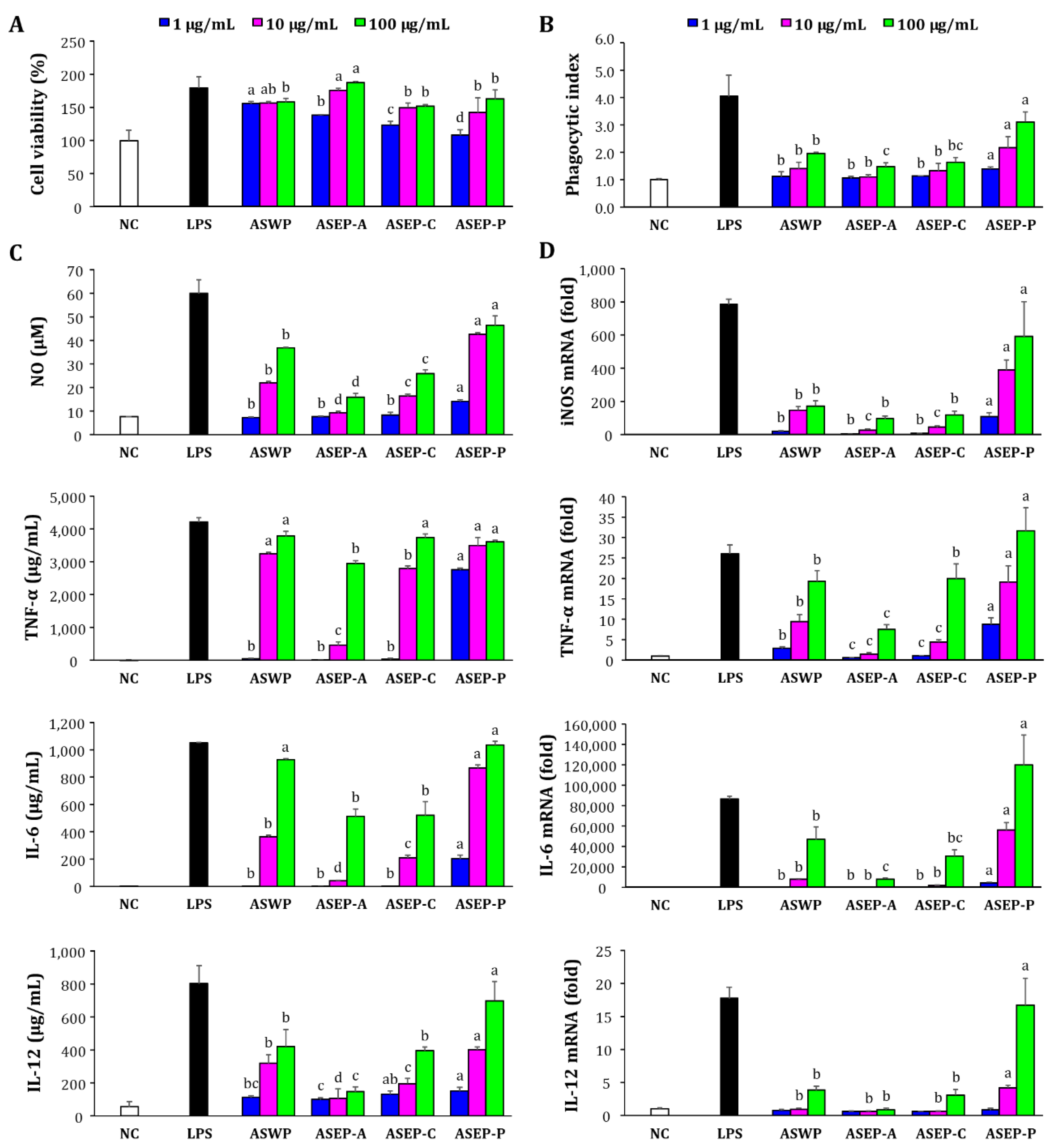
2.3. Comparison of Macrophage Immunostimulatory Activity of ASWP and Three ASEPs
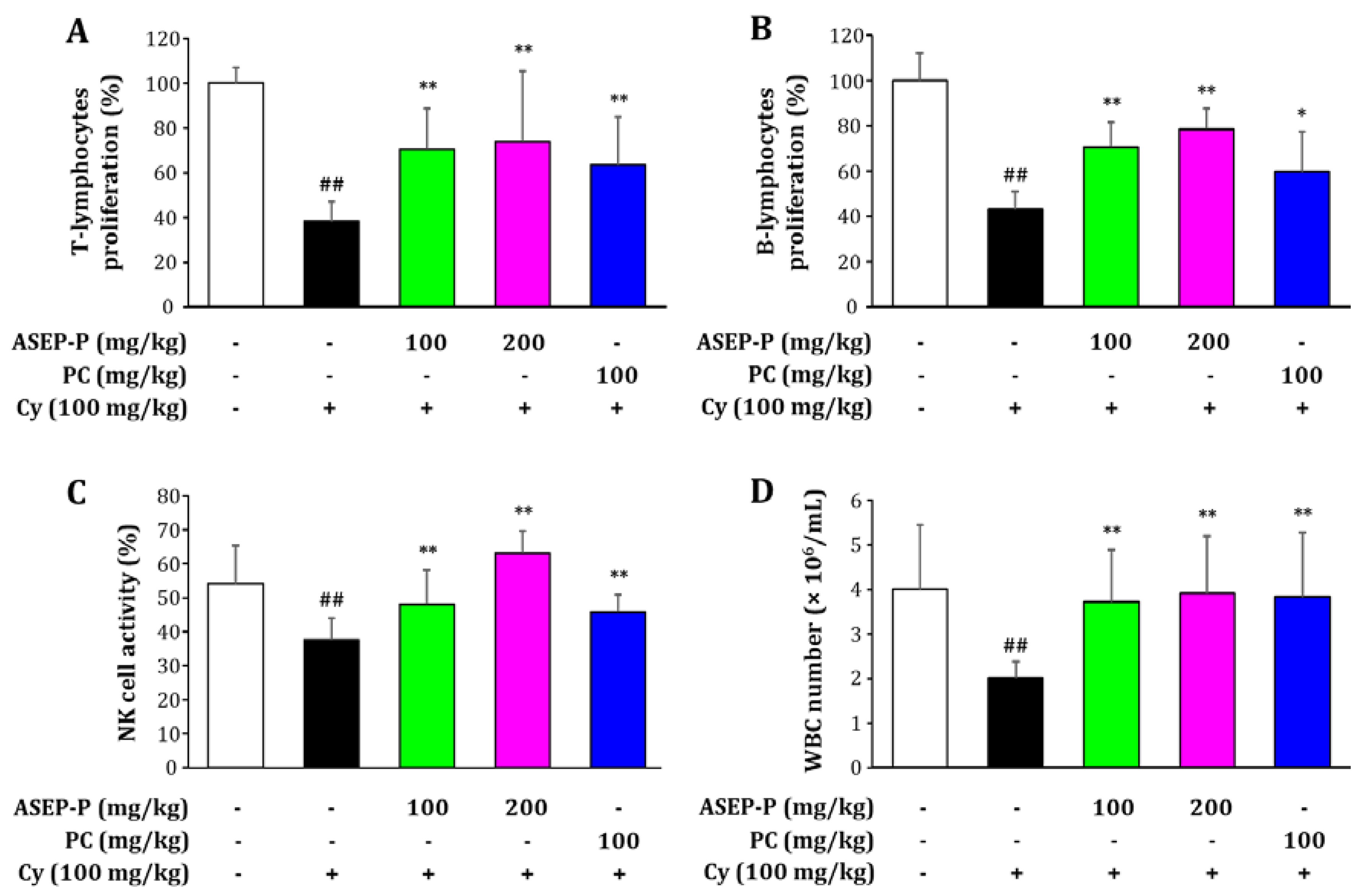
2.4. Immuno-Enhancement Effects of ASEP-P In Vivo
2.4.1. Effects on Immune Organ Indices and Splenic Lymphocyte Proliferation in Normal Mice
2.4.2. Effects on Splenic Lymphocyte Proliferation, Natural Killer Cell Activity, and White Blood Cell Count in Cyclophosphamide-Treated Mice
3. Materials and Methods
3.1. Materials
3.2. Extraction of Polysaccharides from Aster scaber
3.2.1. Polysaccharides Obtained Using Enzyme-Assisted Extraction
3.2.2. Polysaccharides Obtained Using Hot Water Extraction
3.3. Physicochemical Properties of Polysaccharides
3.3.1. Chemical Component Analysis
3.3.2. Monosaccharide Composition Analysis
3.3.3. Molecular Weight Determination
3.3.4. Infrared Analysis
3.4. Immunostimulatory Activities of Polysaccharides In Vitro
3.4.1. Cell Culture
3.4.2. Cell Viability Assay
3.4.3. Phagocytic Activity Assay
3.4.4. Measurement of NO and Cytokine Production
3.4.5. Real-Time Quantitative PCR
3.5. Immuno-Enhancement Effects of ASEP-P In Vivo
3.5.1. Experimental Animals
3.5.2. Animal Treatments
3.5.3. Measurement of Organ Indexes
3.5.4. Splenocyte Proliferation Assay
3.5.5. NK-Cell Activity Assay and WBC Counting
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASEP-A | amylase-assisted extracted polysaccharide |
| ASEP-P | pectinase-assisted extracted polysaccharide |
| ASEP-C | cellulase-assisted extracted polysaccharide |
| ASWP | hot water extracted polysaccharide |
| RG-II | rhamnogalacturonan-II |
| TNF-α | tumor necrosis factor-alpha |
| IL | interleukin |
| NO | nitric oxide |
| LPS | lipopolysaccharide |
| PBS | phosphate-buffered saline |
| ELISA | enzyme-linked immunosorbent assay |
| qRT-PCR | quantitative reverse transcription–polymerase chain reaction |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| KM | Kunming |
| NK | natural killer |
| WBC | white blood cell |
| KDO | 2-keto-3-deoxy-mannooctanoic acid |
| HPSEC | high-performance size-exclusion chromatography |
| RI | refractive index |
| FT-IR | fourier-transform infrared spectroscopy |
References
- Di, T.; Chen, G.; Sun, Y.; Ou, S.; Zeng, X.; Ye, H. Antioxidant and immunostimulating activities in vitro of sulfated polysaccharides isolated from Gracilaria rubra. J. Funct. Foods 2017, 28, 64–75. [Google Scholar] [CrossRef]
- Liu, J.; Willför, S.; Xu, C. A review of bioactive plant polysaccharides: Biological activities, functionalization, and biomedical applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 31–61. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Wu, Z.; Yang, J.; Li, W.; Shen, L. Extraction, purification and anti-proliferative activities of polysaccharides from Lentinus edodes. Int. J. Boil. Macromol. 2016, 93, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Rostami, H.; Gharibzahedi, S.M.T. Cellulase-assisted extraction of polysaccharides from Malva sylvestris: Process optimization and potential functionalities. Int. J. Boil. Macromol. 2017, 101, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, C.; Zhou, D.; Ou, S.; Huang, H. Structural characterization of a novel polysaccharide fraction from Hericium erinaceus and its signaling pathways involved in macrophage immunomodulatory activity. J. Funct. Foods 2017, 37, 574–585. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.-J. Chemistry and pharmacological action of caffeoylquinic acid derivatives and pharmaceutical utilization of chwinamul (Korean Mountainous vegetable). Arch. Pharmacal. Res. 2010, 33, 1703–1720. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.C.; Jung, C.M.; Shin, C.G.; Lee, J.K.; Choi, S.U.; Kim, S.Y.; Lee, K.R. A new caffeoly quinic acid from aster scaber and its inhibitory activity against human immunodeficiency virus-1 (HIV-1) integrase. Chem. Pharm. Bull. 2000, 48, 1796–1798. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, H.-S. The immunomodulating effects of Aster scaber Thunb extracts in mice. Nutr. Sci. 2002, 5, 203–210. [Google Scholar]
- Kim, J.; Park, C.-S.; Lim, Y.; Kim, H.-S. Paeonia japonica, Houttuynia cordata, and Aster scaber water extracts induce nitric oxide and cytokine production by lipopolysaccharide-activated macrophages. J. Med. Food 2009, 12, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Lee, S.; Park, Y.I.; Lee, J.; Kwon, K.H. Neuroprotective effects of phytosterols and flavonoids from Cirsium setidens and Aster scaber in human brain neuroblastoma SK-N-SH cells. Life Sci. 2016, 148, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-X.; Kim, H.-Y.; Kim, S.-N.; Lee, J.-Y.; Seo, D.; Choi, Y. Comparison of nutritional compositions of green vegetables. J. Korean Soc. Food Sci. Nutr. 2017, 46, 592–599. [Google Scholar] [CrossRef]
- Sung, S.-K.; Rhee, Y.-K.; Cho, C.-W.; Kim, E.-Y.; Kang, D.-Z.; Hong, H.-D. Immunostimulating activites of polysaccharide fractions isolated from Aster scaber Thunb. Korean J. Food Nutr. 2015, 28, 821–828. [Google Scholar] [CrossRef]
- Baik, J.H.; Shin, K.S.; Park, Y.; Yu, K.W.; Suh, H.J.; Choi, H.S. Biotransformation of catechin and extraction of active polysaccharide from green tea leaves via simultaneous treatment with tannase and pectinase. J. Sci. Food Agric. 2015, 95, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, D.; Ning, X.; Yang, G.; Lin, Z.; Tian, M.; Zhou, Y. α-Amylase-assisted extraction of polysaccharides from Panax ginseng. Int. J. Boil. Macromol. 2015, 75, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kwak, B.-S.; Hong, H.-D.; Suh, H.-J.; Shin, K.-S. Structural features of immunostimulatory polysaccharide purified from pectinase hydrolysate of barley leaf. Int. J. Boil. Macromol. 2016, 87, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Nie, C.; Zhu, P.; Ma, S.; Wang, M.; Hu, Y. Purification, characterization and immunomodulatory activity of polysaccharides from stem lettuce. Carbohydr. Polym. 2018, 188, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shang, H.; Yang, J.; Li, R.; Wu, H. Effects of different extraction techniques on physicochemical properties and activities of polysaccharides from comfrey (Symphytum officinale L.) root. Ind. Crop. Prod. 2018, 121, 18–25. [Google Scholar] [CrossRef]
- Li, X.; Wang, L. Effect of extraction method on structure and antioxidant activity of Hohenbuehelia serotina polysaccharides. Int. J. Boil. Macromol. 2016, 83, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Devshri, U.; Janne, H.; Rekha, S.; Salem, S. Enzymatic extraction and characterization of polysaccharide from Tuber aestivum. Bioact. Carbohydr. Diet. Fibre 2017, 10, 1–9. [Google Scholar]
- Yin, C.; Fan, X.; Fan, Z.; Shi, D.; Gao, H. Optimization of enzymes-microwave-ultrasound assisted extraction of Lentinus edodes polysaccharides and determination of its antioxidant activity. Int. J. Boil. Macromol. 2018, 111, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Karaki, N.; Aljawish, A.; Humeau, C.; Muniglia, L.; Jasniewski, J. Enzymatic modification of polysaccharides: Mechanisms, properties, and potential applications: A review. Enzym. Microb. Technol. 2016, 90, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-Y.; Zhang, W.-L.; Jiang, J.-G. Immune-enhancing activity of polysaccharides from Hibiscus sabdariffa Linn. via MAPK and NF-κB signaling pathways in RAW264. 7 cells. J. Funct. Foods 2017, 34, 118–129. [Google Scholar] [CrossRef]
- Shen, C.-Y.; Jiang, J.-G.; Li, M.-Q.; Zheng, C.-Y.; Zhu, W. Structural characterization and immunomodulatory activity of novel polysaccharides from Citrus aurantium Linn. variant amara Engl. J. Funct. Foods 2017, 35, 352–362. [Google Scholar] [CrossRef]
- Černá, M.; Barros, A.S.; Nunes, A.; Rocha, S.M.; Delgadillo, I.; Čopíková, J.; Coimbra, M.A. Use of FT-IR spectroscopy as a tool for the analysis of polysaccharide food additives. Carbohydr. Polym. 2003, 51, 383–389. [Google Scholar] [CrossRef]
- Song, Y.-R.; Sung, S.-K.; Jang, M.; Lim, T.-G.; Cho, C.-W.; Han, C.-J.; Hong, H.-D. Enzyme-assisted extraction, chemical characteristics, and immunostimulatory activity of polysaccharides from Korean ginseng (Panax ginseng Meyer). Int. J. Boil. Macromol. 2018, 116, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qi, Q.; Li, A.; Yang, M.; Huang, W.; Xu, H.; Zhao, Z.; Li, S. Immuno-enhancement effects of Yifei Tongluo Granules on cyclophosphamide-induced immunosuppression in Balb/c mice. J. Ethnopharmacol. 2016, 194, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.-D.; Kim, J.-C.; Lim, T.-G.; Song, Y.-R.; Cho, C.-W.; Jang, M. Mixing ratio optimization for functional complex extracts of Rhodiola crenulata, Panax quinquefolius, and Astragalus membranaceus using mixture design and verification of immune functional efficacy in animal models. J. Funct. Foods 2018, 40, 447–454. [Google Scholar] [CrossRef]
- Cho, C.-W.; Han, C.-J.; Rhee, Y.K.; Lee, Y.-C.; Shin, K.-S.; Shin, J.-S.; Lee, K.-T.; Hong, H.-D. Cheonggukjang polysaccharides enhance immune activities and prevent cyclophosphamide-induced immunosuppression. Int. J. Boil. Macromol. 2015, 72, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, M.; Sun, R.; Pan, L. Extraction, characterization of a Ginseng fruits polysaccharide and its immune modulating activities in rats with Lewis lung carcinoma. Carbohydr. Polym. 2015, 127, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Karkhanis, Y.D.; Zeltner, J.Y.; Jackson, J.J.; Carlo, D.J. A new and improved microassay to determine 2-keto-3-deoxyoctonate in lipopolysaccharide of Gram-negative bacteria. Anal. Biochem. 1978, 85, 595–601. [Google Scholar] [CrossRef]
- Wang, M.; Guilbert, L.J.; Ling, L.; Li, J.; Wu, Y.; Xu, S.; Pang, P.; Shan, J.J. Immunomodulating activity of CVT-E002, a proprietary extract from North American ginseng (Panax quinquefolium). J. Pharm. Pharmacol. 2001, 53, 1515–1523. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | ASWP | ASEP-A | ASEP-C | ASEP-P |
|---|---|---|---|---|
| Yield (%) | 3.3 ab | 3.4 ab | 3.1 b | 3.8 a |
| Chemical composition (%) (1) | ||||
| Neutral sugar | 45.7 b | 59.9 a | 44.6 b | 58.5 a |
| Uronic acid | 51.6 a | 39.1 b | 54.1 a | 40.1 b |
| Protein | 2.3 a | 0.6 bc | 0.7 b | 0.3 c |
| KDO (2)-like material | 0.4 b | 0.4 b | 0.6 b | 1.2 a |
| Component sugar (molar ratio) | ||||
| Fucose | 0.06 | 0.18 | 0.05 | 0.23 |
| Rhamnose | 0.44 | 1.59 | 0.62 | 2.08 |
| Arabinose | 1.00 | 0.75 | 1.25 | 1.11 |
| Galactose | 0.44 | 1.14 | 0.67 | 1.74 |
| Glucose | 1.00 | 1.00 | 1.00 | 1.00 |
| Mannose | 0.01 | 0.19 | 0.02 | 0.17 |
| Xylose | 0.03 | 0.13 | 0.05 | 0.14 |
| Galacturonic acid | 3.15 | 1.50 | 2.63 | 2.03 |
| Glucuronic acid | 0.05 | 0.13 | 0.04 | 0.18 |
| Group | Body Weight (g) | Immune Organ Indices (mg/g) | Splenocyte Proliferation (%) | |||
|---|---|---|---|---|---|---|
| Initial | Final | Spleen | Thymus | T-Lymphocytes | B-Lymphocytes | |
| Control | 22.00 ± 1.79 | 34.30 ± 3.46 | 0.62 ± 0.06 | 0.33 ± 0.13 | 100.0 ± 14.8 | 100.0 ± 11.8 |
| ASEP-P (100 mg/kg) | 22.13 ± 1.84 | 34.33 ± 3.28 | 0.70 ± 0.07 | 0.38 ± 0.09 | 113.1 ± 18.6 * | 115.7 ± 19.4 * |
| ASEP-P (200 mg/kg) | 21.75 ± 1.63 | 34.25 ± 3.93 | 0.74 ± 0.10 * | 0.41 ± 0.10 | 125.1 ± 11.7 ** | 124.2 ± 20.2 ** |
| PC (100 mg/kg) | 21.94 ± 1.75 | 34.21 ± 2.93 | 0.76 ± 0.07 * | 0.41 ± 0.13 | 124.4 ± 14.1 ** | 123.0 ± 13.8 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.-R.; Sung, S.-K.; Shin, E.-J.; Cho, C.-W.; Han, C.-J.; Hong, H.-D. The Effect of Pectinase-Assisted Extraction on the Physicochemical and Biological Properties of Polysaccharides from Aster scaber. Int. J. Mol. Sci. 2018, 19, 2839. https://doi.org/10.3390/ijms19092839
Song Y-R, Sung S-K, Shin E-J, Cho C-W, Han C-J, Hong H-D. The Effect of Pectinase-Assisted Extraction on the Physicochemical and Biological Properties of Polysaccharides from Aster scaber. International Journal of Molecular Sciences. 2018; 19(9):2839. https://doi.org/10.3390/ijms19092839
Chicago/Turabian StyleSong, Young-Ran, Su-Kyung Sung, Eun-Ju Shin, Chang-Won Cho, Chun-Ji Han, and Hee-Do Hong. 2018. "The Effect of Pectinase-Assisted Extraction on the Physicochemical and Biological Properties of Polysaccharides from Aster scaber" International Journal of Molecular Sciences 19, no. 9: 2839. https://doi.org/10.3390/ijms19092839
APA StyleSong, Y. -R., Sung, S. -K., Shin, E. -J., Cho, C. -W., Han, C. -J., & Hong, H. -D. (2018). The Effect of Pectinase-Assisted Extraction on the Physicochemical and Biological Properties of Polysaccharides from Aster scaber. International Journal of Molecular Sciences, 19(9), 2839. https://doi.org/10.3390/ijms19092839