Cap-Independent mRNA Translation in Germ Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Sorting Roles for CD and CI Translation in Differentiating Meiotic Cells
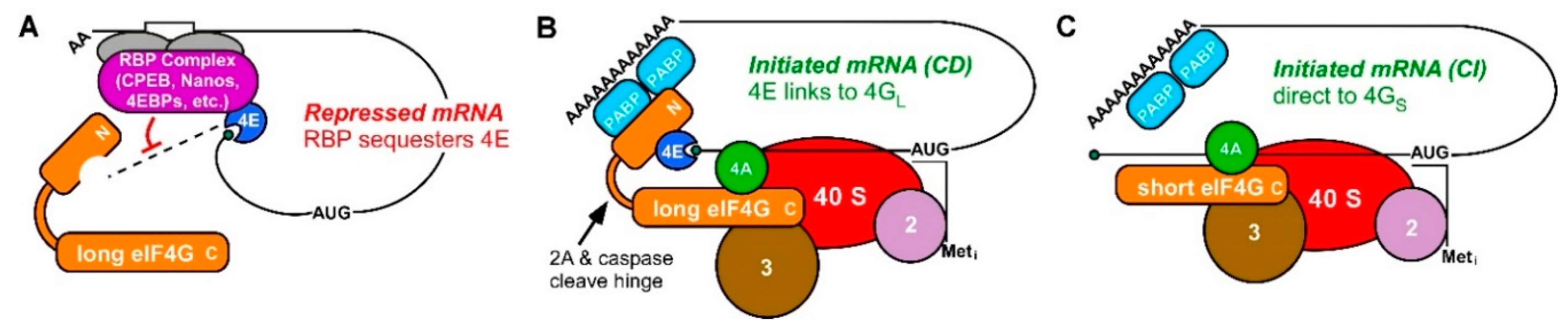
2.1. Translational Control in Development Has Focused on Repression
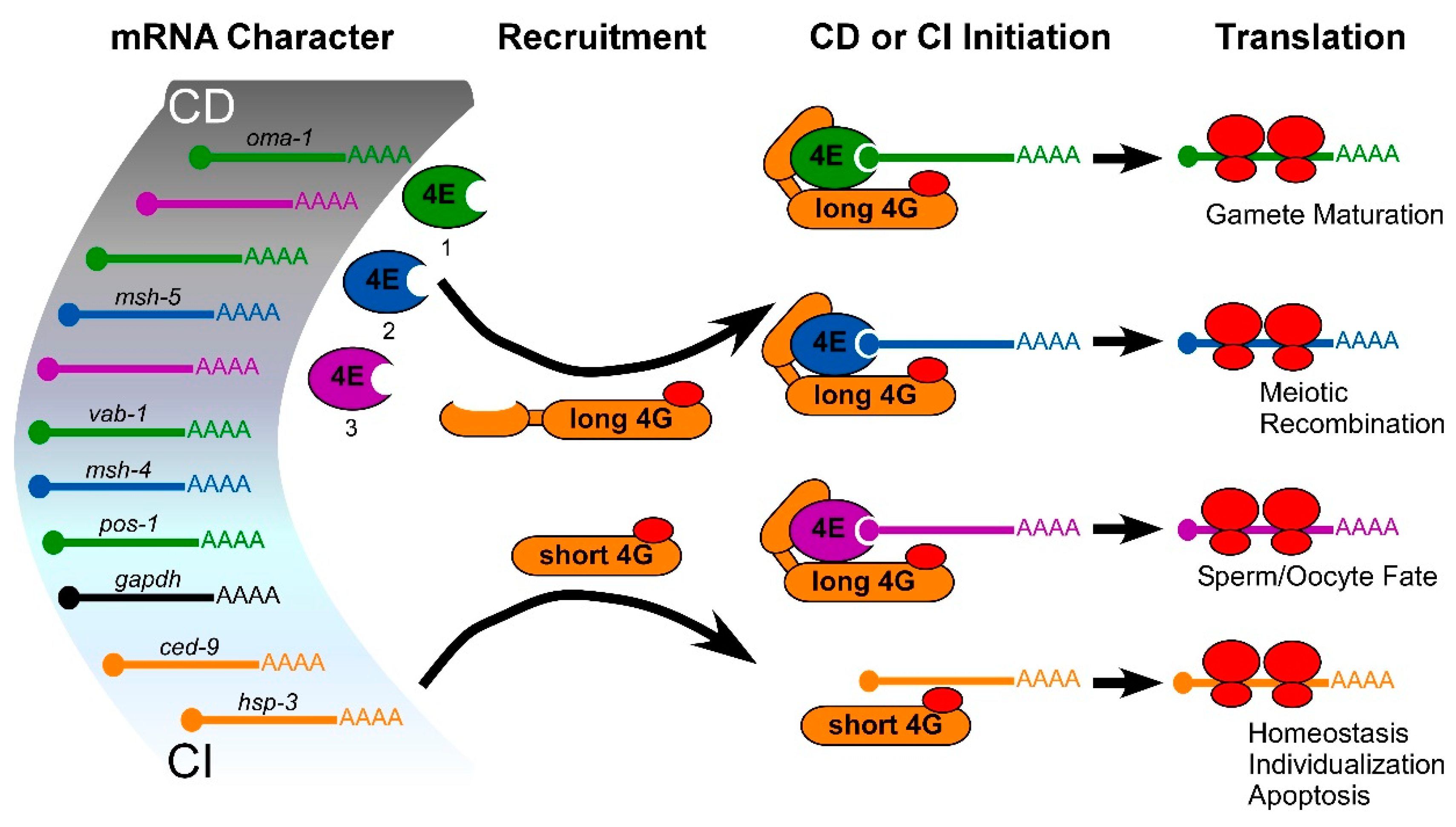
2.2. Germ Cell Translation Does Not Follow the Rules; the Prevalence of CI Translation in Frog Oocytes
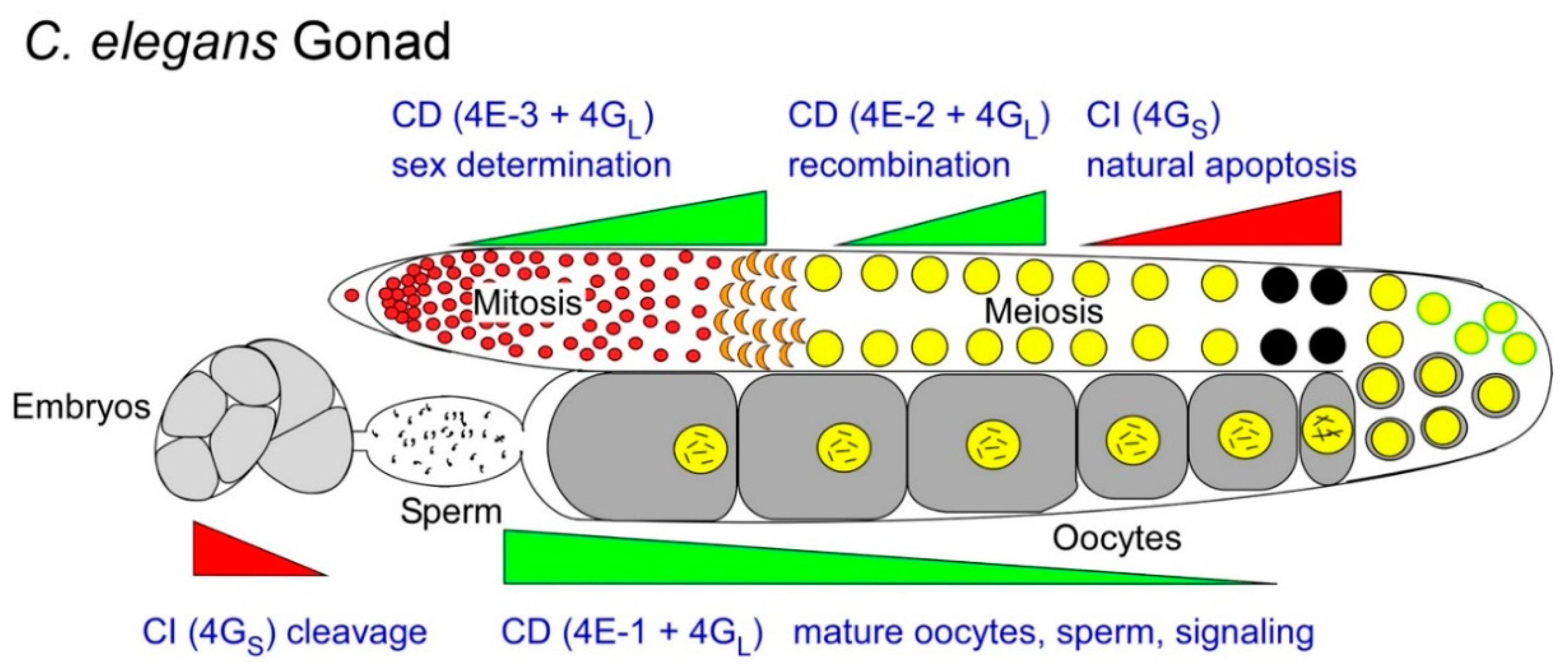
2.3. A Use for CI Translation in Germ Cell Homeostasis; Evidence from Worm Oocytes
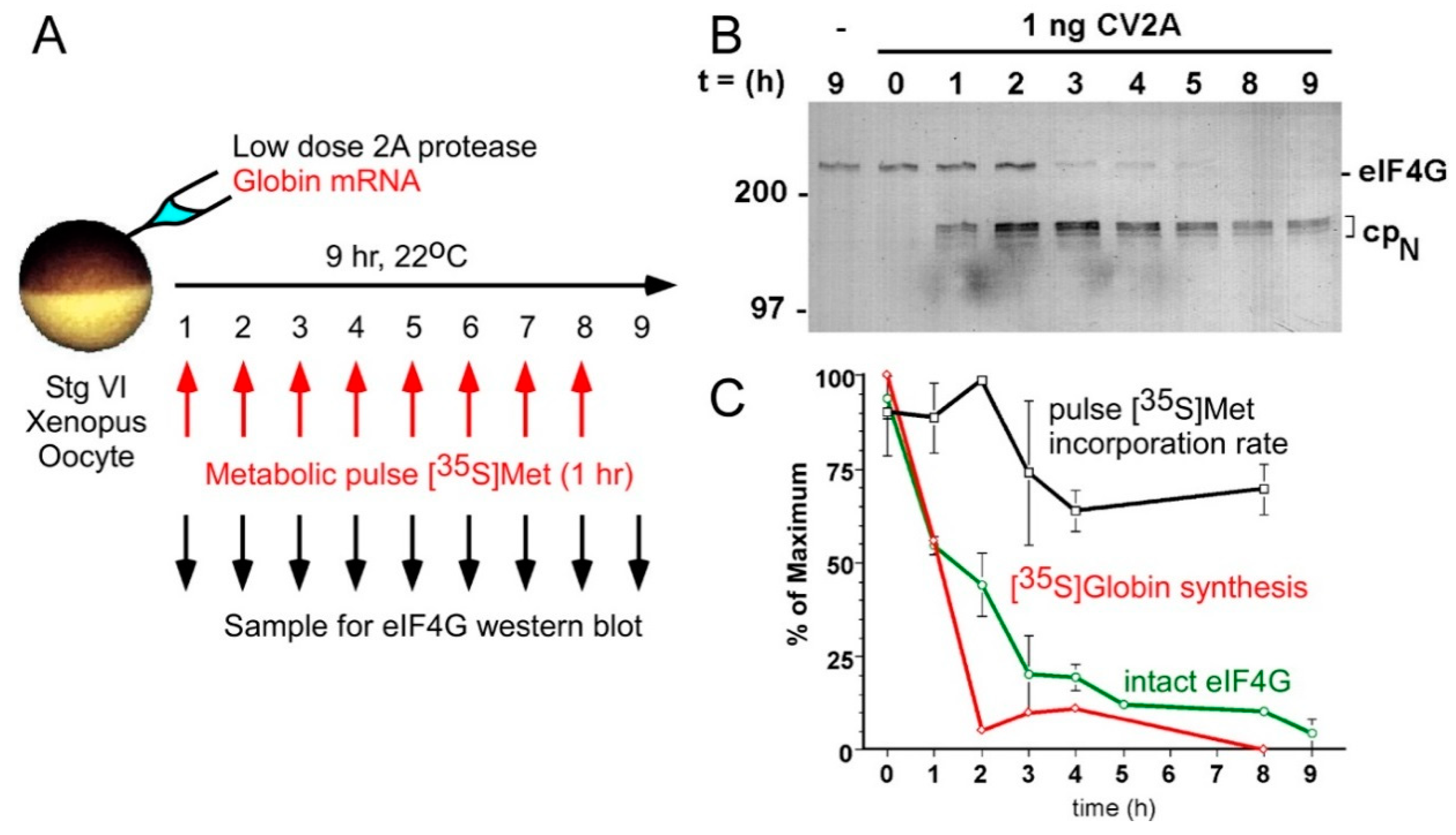
2.4. Germ Cells also Use CI Translation of “Death-Promoting” mRNAs to Differentiate
2.5. Even CD Translation in Germ Cells Comes in Multiple Flavors
2.6. Potential Relevance of CI Translation to RNA Viruses That Cause Birth Defects
3. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| UTR | Untranslated region |
| eIF | Eukaryotic (translation) initiation factor |
| 4GS | Short form of eIF4G |
| 4GL | Long form of eIF4G |
| PABP | Poly(A) binding protein |
| IRES | Internal ribosome entry site |
| RBP | RNA binding protein |
| EMCV | Encephalomyocarditis virus |
| mRNP | Messenger ribonuclear protein |
| CI | Cap-independent |
| CD | Cap-dependent |
| CITE | Cap-independent translational enhancer |
| CPEB | Cytoplasmic polyadenylation element binding protein |
| MPF | Maturation promoting factor (cyclin B/CDK2) |
References
- Jackson, R.J.; Kaminski, A. Internal Initiation of translation in eukaryotes: The picornavirus paradigm and beyond. RNA 1995, 1, 985–1000. [Google Scholar] [PubMed]
- Ehrenfeld, E. Initiation of translation by picornavirus RNAs. In Translational Control; Hershey, J.W.B., Mathews, M.B., Sonenberg, N., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1996; pp. 549–573. [Google Scholar]
- Hernandez, G. Was the initiation of translation in early eukaryotes IRES-driven? Trends Biochem. Sci. 2008, 33, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.M.; Kirchweger, R.; Ziegler, E.; Rhoads, R.E.; Skern, T.; Kean, K.M. eIF4G and its proteolytic cleavage products: Effects on initiation of protein synthesis from capped, uncapped, and IRES-containing mRNAs. RNA 1997, 3, 186–196. [Google Scholar] [PubMed]
- Ziegler, E.; Borman, A.M.; Deliat, F.G.; Liebig, H.-D.; Jugovic, D.; Kean, K.M.; Skern, T.; Kuechler, E. Picornavirus 2A protease-mediated stimulation of internal initiation of translation is dependent on enzymatic activity and the cleavage of products of cellular proteins. Virology 1995, 213, 549–557. [Google Scholar] [CrossRef]
- Shatkin, A.J. mRNA caps-old and newer hats. Bioessays 1987, 7, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, H.; Li, H.; McGuire, A.M.; Fletcher, C.M.; Gingras, A.C.; Sonenberg, N.; Wagner, G. Structure of translation factor eIF4E bound to m7GDP and interaction with 4E-binding protein. Nat. Struct. Biol. 1997, 4, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, Y.; LaFiandra, A.; Shatkin, A.J. 5’-Terminal structure and mRNA stability. Nature 1977, 266, 235–239. [Google Scholar] [CrossRef]
- Rhoads, R.E. Cap recognition and the entry of mRNA into the protein synthesis initiation cycle. Trends Biochem. Sci. 1988, 13, 52–56. [Google Scholar] [CrossRef]
- Hambidge, S.J.; Sarnow, P. Translational enhancement of the poliovirus 5’ noncoding region mediated by virus-encoded polypeptide 2A. Proc. Natl. Acad. Sci. USA 1992, 89, 10272–10276. [Google Scholar] [CrossRef]
- Borman, A.M.; Bailly, J.-L.; Girard, M.; Kean, K.M. Picornavirus internal ribosome entry segments: Comparison of translation efficiency and the requirements for optimal internal initiation of translation in vitro. Nucleic Acids Res. 1995, 23, 3656–3663. [Google Scholar] [CrossRef]
- Graber, T.E.; Holcik, M. Cap-independent regulation of gene expression in apoptosis. Mol. Biosyst. 2007, 3, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Sonenberg, N. Translational control in stress and apoptosis. Nat. Rev. Mol. Cell Biol. 2005, 6, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, R.E.; Lamphear, B.J. Cap-independent translation of heat shock messenger RNAs. Curr. Top. Microbiol. Immunol. 1995, 203, 131–153. [Google Scholar] [PubMed]
- Schneider, R.J. Cap-independent translation in adenovirus infected cells. Curr. Top. Microbiol. Immunol. 1995, 203, 117–129. [Google Scholar] [PubMed]
- Lopez-Lastra, M.; Rivas, A.; Barria, M.I. Protein synthesis in eukaryotes: The growing biological relevance of cap-independent translation initiation. Biol. Res. 2005, 38, 121–146. [Google Scholar] [CrossRef] [PubMed]
- Bushell, M.; McKendrick, L.; Janicke, R.U.; Clemens, M.J.; Morley, S.J. Caspase-3 is necessary and sufficient for cleavage of protein synthesis eukaryotic initiation factor 4G during apoptosis. FEBS Lett. 1999, 451, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Marissen, W.E.; Gradi, A.; Sonenberg, N.; Lloyd, R.E. Cleavage of eukaryotic translation initiation factor 4GII correlates with translation inhibition during apoptosis. Cell Death Differ. 2000, 7, 1234–1243. [Google Scholar] [CrossRef] [Green Version]
- Marissen, W.E.; Lloyd, R.E. Eukaryotic translation initiation factor 4G is targeted for proteolytic cleavage by caspase 3 during inhibition of translation in apoptotic cells. Mol. Cell. Biol. 1998, 18, 7565–7574. [Google Scholar] [CrossRef]
- Nevins, T.A.; Harder, Z.M.; Korneluk, R.G.; Holcik, M. Distinct regulation of internal ribosome entry site-mediated translation following cellular stress is mediated by apoptotic fragments of eIF4G translation initiation factor family members eIF4GI and p97/DAP5/NAT1. J. Biol. Chem. 2003, 278, 3572–3579. [Google Scholar] [CrossRef]
- Prevot, D.; Darlix, J.L.; Ohlmann, T. Conducting the initiation of protein synthesis: The role of eIF4G. Biol. Cell 2003, 95, 141–156. [Google Scholar] [CrossRef]
- Hosszu Ungureanu, N.; Cloutier, M.; Lewis, S.M.; de Silva, N.; Blais, J.D.; Bell, J.C.; Holcik, M. IRES-mediated translation of Apaf-1, but not XIAP, is regulated during UV-induced cell death. J. Biol. Chem. 2006, 281, 15155–15163. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.K.; Friday, A.J.; Henderson, M.A.; Hao, E.; Keiper, B.D. Induction of cap-independent BiP (hsp-3) and Bcl-2 (ced-9) translation in response to eIF4G (IFG-1) depletion in C. elegans. Translation 2014, 2, e28935. [Google Scholar] [CrossRef]
- Malys, N.; McCarthy, J.E. Translation initiation: Variations in the mechanism can be anticipated. Cell. Mol. Life Sci. 2011, 68, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.S.; St George, J.P.; McCall, K. Programmed cell death in the germline. Semin. Cell Dev. Biol. 2005, 16, 245–259. [Google Scholar] [CrossRef]
- Kinchen, J.M.; Hengartner, M.O. Tales of cannibalism, suicide, and murder: Programmed cell death in C. elegans. Curr. Top. Dev. Biol. 2005, 65, 1–45. [Google Scholar] [PubMed]
- Shatsky, I.N.; Terenin, I.M.; Smirnova, V.V.; Andreev, D.E. Cap-Independent Translation: What’s in a Name? Trends Biochem. Sci. 2018, 43, 882–895. [Google Scholar] [CrossRef] [PubMed]
- Johannes, G.; Carter, M.S.; Eisen, M.B.; Brown, P.O.; Sarnow, P. Identification of eukaryotic mRNAs that are translated at reduced cap binding complex eIF4F concentrations using a cDNA microarray. Proc. Natl. Acad. Sci. USA 1999, 96, 13118–13123. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Sarnow, P. Location of the internal ribosome entry site in the 5’ non-coding region of the immunoglobulin heavy-chain binding protein (BiP) mRNA: Evidence for specific RNA-protein interactions. Nucleic Acids Res. 1997, 25, 2800–2807. [Google Scholar] [CrossRef]
- Macejak, D.G.; Sarnow, P. Translational regulation of the immunoglobulin heavy-chain binding protein mRNA. Enzyme 1990, 44, 310–319. [Google Scholar] [CrossRef]
- Philippe, C.; Dubrac, A.; Quelen, C.; Desquesnes, A.; Van Den Berghe, L.; Segura, C.; Filleron, T.; Pyronnet, S.; Prats, H.; Brousset, P.; et al. PERK mediates the IRES-dependent translational activation of mRNAs encoding angiogenic growth factors after ischemic stress. Sci. Signal. 2016, 9, ra44. [Google Scholar] [CrossRef]
- Vagner, S.; Gensac, M.C.; Mare, A.; Baynard, F.; Amalric, F.; Prats, H.; Prats, A.C. Alternative translation of human fibroblast growth factor 2 mRNA occurs by internal entry of ribosomes. Mol. Cell. Biol. 1995, 15, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Silvera, D.; Schneider, R.J. Inflammatory breast cancer cells are constitutively adapted to hypoxia. Cell Cycle 2009, 8, 3091–3096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundsdoerfer, P.; Thoma, C.; Hentze, M.W. Eukaryotic translation initiation factor 4GI and p97 promote cellular internal ribosome entry sequence-driven translation. Proc. Natl. Acad. Sci. USA 2005, 102, 13421–13426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coldwell, M.J.; Mitchell, S.A.; Stoneley, M.; MacFarlane, M.; Willis, A.E. Initiation of Apaf-1 translation by internal ribosome entry. Oncogene 2000, 19, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holcik, M.; Gordon, B.W.; Korneluk, R.G. The internal ribosome entry site-mediated translation of antiapoptotic protein XIAP is modulated by the heterogeneous nuclear ribonucleoproteins C1 and C2. Mol. Cell. Biol. 2003, 23, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Sherrill, K.W.; Byrd, M.P.; Van Eden, M.E.; Lloyd, R.E. BCL-2 translation is mediated via internal ribosome entry during cell stress. J. Biol. Chem. 2004, 279, 29066–29074. [Google Scholar] [CrossRef] [PubMed]
- Komar, A.A.; Hatzoglou, M. Internal ribosome entry sites in cellular mRNAs: Mystery of their existence. J. Biol. Chem. 2005, 280, 23425–23428. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.D.; Zhou, F.; Harris, M.S.; Ingolia, N.T.; Hinnebusch, A.G. eIF4B stimulates translation of long mRNAs with structured 5’ UTRs and low closed-loop potential but weak dependence on eIF4G. Proc. Natl. Acad. Sci. USA 2016, 113, 10464–10472. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Park, S.M.; Kim, T.D.; Kim, J.H.; Kim, K.T.; Jang, S.K. BiP internal ribosomal entry site activity is controlled by heat-induced interaction of NSAP1. Mol. Cell. Biol. 2007, 27, 368–383. [Google Scholar] [CrossRef] [PubMed]
- Dinkova, T.D.; Zepeda, H.; Martinez-Salas, E.; Martinez, L.M.; Nieto-Sotelo, J.; de Jimenez, E.S. Cap-independent translation of maize Hsp101. Plant J. 2005, 41, 722–731. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, W.V. Alternative ways to think about cellular internal ribosome entry. J. Biol. Chem. 2010, 285, 29033–29038. [Google Scholar] [CrossRef] [PubMed]
- Joshi-Barve, S.; DeBenedetti, A.; Rhoads, R.E. Preferential translation of heat shock mRNAs in HeLa cells deficient in protein synthesis initiation factors eIF-4E and eIF-4g. J. Biol. Chem. 1992, 267, 21038–21043. [Google Scholar] [PubMed]
- Kraft, J.J.; Treder, K.; Peterson, M.S.; Miller, W.A. Cation-dependent folding of 3’ cap-independent translation elements facilitates interaction of a 17-nucleotide conserved sequence with eIF4G. Nucleic Acids Res. 2013, 41, 3398–3413. [Google Scholar] [CrossRef] [PubMed]
- Miras, M.; Truniger, V.; Querol-Audi, J.; Aranda, M.A. Analysis of the interacting partners eIF4F and 3’-CITE required for Melon necrotic spot virus cap-independent translation. Mol. Plant Pathol. 2017, 18, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5’ UTR m(6)A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef]
- Sweeney, T.R.; Abaeva, I.S.; Pestova, T.V.; Hellen, C.U. The mechanism of translation initiation on Type 1 picornavirus IRESs. EMBO J. 2014, 33, 76–92. [Google Scholar] [CrossRef]
- Mitchell, S.A.; Brown, E.C.; Coldwell, M.J.; Jackson, R.J.; Willis, A.E. Protein factor requirements of the Apaf-1 internal ribosome entry segment: Roles of polypyrimidine tract binding protein and upstream of N-ras. Mol. Cell. Biol. 2001, 21, 3364–3374. [Google Scholar] [CrossRef]
- Gradi, A.; Imataka, H.; Svitkin, Y.V.; Rom, E.; Raught, B.; Morino, S.; Sonenberg, N. A novel functional human eukaryotic translation initiation factor 4G. Mol. Cell. Biol. 1998, 18, 334–342. [Google Scholar] [CrossRef]
- Yoffe, Y.; David, M.; Kalaora, R.; Povodovski, L.; Friedlander, G.; Feldmesser, E.; Ainbinder, E.; Saada, A.; Bialik, S.; Kimchi, A. Cap-independent translation by DAP5 controls cell fate decisions in human embryonic stem cells. Genes Dev. 2016, 30, 1991–2004. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, A.-M.; Sonenberg, N. Proteolysis of the p220 component of the cap-binding protein complex is not sufficient for complete inhibition of host cell protein synthesis after poliovirus infection. J. Virol. 1987, 61, 986–991. [Google Scholar]
- Aldabe, R.; Feduchi, E.; Novoa, I.; Carrasco, L. Expression of poliovirus 2Apro in mammalian cells: Effects on translation. FEBS Lett. 1995, 377, 1–5. [Google Scholar] [CrossRef]
- IIzuka, N.; Najita, L.; Franzusoff, A.; Sarnow, P. Cap-dependent and cap-independent translation by internal initiation of mRNAs in cell extracts prepared from Saccharomyces cerevisiae. Mol. Cell. Biol. 1994, 14, 7322–7330. [Google Scholar] [CrossRef] [PubMed]
- Ohlmann, T.; Rau, M.; Pain, V.M.; Morley, S.J. The C-terminal domain of eukaryotic protein synthesis initiation factor (eIF) 4G is sufficient to support cap-independent translation in the absence of eIF4E. EMBO J. 1996, 15, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Hentze, M.W. eIF4G: A multipurpose ribosome adaptor? Science 1997, 275, 500–501. [Google Scholar] [CrossRef] [PubMed]
- Keiper, B.D.; Gan, W.; Rhoads, R.E. Protein synthesis initiation factor 4G. Int. J. Biochem. Cell Biol. 1999, 31, 37–41. [Google Scholar] [CrossRef]
- Dworkin, M.B.; Dworkin-Rastl, E. Functions of maternal mRNA in early development. Mo.l Reprod. Dev. 1990, 26, 261–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nousch, M.; Eckmann, C.R. Translational control in the Caenorhabditis elegans germ line. Adv. Exp. Med. Biol. 2013, 757, 205–247. [Google Scholar]
- Tadros, W.; Lipshitz, H.D. Setting the stage for development: mRNA translation and stability during oocyte maturation and egg activation in Drosophila. Dev. Dyn. 2005, 232, 593–608. [Google Scholar] [CrossRef]
- Friday, A.J.; Keiper, B.D. Positive mRNA Translational Control in Germ Cells by Initiation Factor Selectivity. Biomed Res. Int. 2015, 2015, e327963. [Google Scholar] [CrossRef]
- Mendez, R.; Richter, J.D. Translational control by CPEB: A means to the end. Nat. Rev. Mol. Cell Biol. 2001, 2, 521–529. [Google Scholar] [CrossRef]
- Cao, Q.; Richter, J.D. Dissolution of the maskin-eIF4E complex by cytoplasmic polyadenylation and poly(A)-binding protein controls cyclin B1 mRNA translation and oocyte maturation. EMBO J. 2002, 21, 3852–3862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, P.M.; Smibert, C.A. Translational regulation of maternal mRNAs. Curr. Opin. Genet. Dev. 1996, 6, 403–407. [Google Scholar] [CrossRef]
- Varnum, S.M.; Wormington, W.M. Deadenylation of maternal mRNAs during Xenopus oocyte maturation does not require specific cis-sequences: A default mechanism for translational control. Genes Dev. 1990, 4, 2278–2286. [Google Scholar] [CrossRef] [PubMed]
- Wormington, M.; Searfoss, A.M.; Hurney, C.A. Overexpression of poly(A) binding protein prevents maturation-specific deadenylation and translational inactivation in Xenopus oocytes. EMBO J. 1996, 15, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Tarun, S.Z.J.; Sachs, A.B. A common function for mRNA 5’ and 3’ ends in translation initiation in yeast. Genes Dev. 1995, 9, 2997–3007. [Google Scholar] [CrossRef] [PubMed]
- Wells, S.E.; Hillner, P.E.; Vale, R.D.; Sachs, A.B. Circularization of mRNA by eukaryotic translation initiation factors. Mol. Cell 1998, 2, 135–140. [Google Scholar] [CrossRef]
- Mancera-Martinez, E.; Brito Querido, J.; Valasek, L.S.; Simonetti, A.; Hashem, Y. ABCE1: A special factor that orchestrates translation at the crossroad between recycling and initiation. RNA Biol. 2017, 14, 1279–1285. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Mamillapalli, S.S.; Keiper, B.D.; Cha, D.S. A Systematic mRNA Control Mechanism for Germline Stem Cell Homeostasis and Cell Fate Specification. BMB Rep. 2015, 2015, 3259. [Google Scholar] [CrossRef]
- Parisi, M.; Lin, H. Translational repression: A duet of Nanos and Pumilio. Curr. Biol. 2000, 10, R81–R83. [Google Scholar] [CrossRef]
- Keiper, B.D. Translation of mRNAs in Xenopus oocytes. In Encyclopedia of Life Sciences; Nature Publishing Company: London, UK, 2003; pp. 1–6. [Google Scholar]
- Richter, J.D.; Smith, L.D. Differential capacity for translation and lack of competition between mRNAs that segregate to free and membrane-bound polysomes. Cell 1981, 27, 183–191. [Google Scholar] [CrossRef]
- Richter, J.D.; Wasserman, W.J.; Smith, L.D. The mechanism for increased protein synthesis during oocyte maturation. Dev. Biol. 1982, 89, 159–167. [Google Scholar] [CrossRef]
- Asselbergs, F.A.M.; Peters, W.H.M.; van Venrooij, W.J.; Bloemendal, H. Cap analogues do not inhibit mRNA translation in Xenopus laevis oocyte. FEBS Lett. 1978, 94, 195–198. [Google Scholar] [CrossRef]
- Keiper, B.D.; Rhoads, R.E. Cap-independent translation initiation in Xenopus oocytes. Nucleic Acids Res. 1997, 25, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Skern, T.; Liebig, H.-D. Picornains 2A and 3C. Methods Enzymol. 1994, 244, 583–595. [Google Scholar]
- Kerekatte, V.; Keiper, B.D.; Badorff, C.; Cai, A.; Knowlton, K.U.; Rhoads, R.E. Cleavage of poly(A)-binding protein by coxsackievirus 2A protease in vitro and in vivo: Another mechanism for host protein synthesis shutoff? J. Virol. 1999, 73, 709–717. [Google Scholar] [PubMed]
- De Gregorio, E.; Preiss, T.; Hentze, M.W. Translation driven by an eIF4G core domain in vivo. EMBO J. 1999, 18, 4865–4874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrew, L.L.; Dworkin-Rastl, E.; Dworkin, M.B.; Richter, J.D. Poly(A) elongation during Xenopus oocyte maturation is required for translational recruitment and is mediated by a short sequence element. Genes Dev. 1989, 3, 803–815. [Google Scholar] [CrossRef]
- Dworkin, M.B.; Dworkin-Rastl, E. Changes in RNA titers and polyadenylation during oogenesis and oocyte maturation in Xenopus laevis. Dev. Biol. 1985, 112, 451–457. [Google Scholar] [CrossRef]
- Keiper, B.D.; Rhoads, R.E. Translational recruitment of Xenopus maternal mRNAs in response to poly(A) elongation requires initiation factor eIF4G-1. Dev. Biol. 1999, 206, 1–14. [Google Scholar] [CrossRef]
- Laskey, R.A.; Mills, A.D.; Gurdon, J.B.; Partington, G.A. Protein synthesis in oocytes of Xenopus laevis is not regulated by the supply of messenger RNA. Cell 1977, 11, 345–351. [Google Scholar] [CrossRef]
- Gamarnik, A.V.; Andino, R. Replication of poliovirus in Xenopus oocytes requires two human factors. EMBO J. 1996, 15, 5988–5998. [Google Scholar] [CrossRef]
- Contreras, V.; Richardson, M.A.; Hao, E.; Keiper, B.D. Depletion of the cap-associated isoform of translation factor eIF4G induces germline apoptosis in C. elegans. Cell Death Differ. 2008, 15, 1232–1242. [Google Scholar] [CrossRef]
- Hu, J.; Sun, F.; Handel, M.A. Nuclear localization of EIF4G3 suggests a role for the XY body in translational regulation during spermatogenesis in mice. Biol. Reprod. 2018, 98, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Palmer, K.; Handel, M.A. Mutation of Eif4g3, encoding a eukaryotic translation initiation factor, causes male infertility and meiotic arrest of mouse spermatocytes. Development 2010, 137, 1699–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Domenico, E.; Owens, N.D.; Grant, I.M.; Gomes-Faria, R.; Gilchrist, M.J. Molecular asymmetry in the 8-cell stage Xenopus tropicalis embryo described by single blastomere transcript sequencing. Dev. Biol. 2017, 408, 252–268. [Google Scholar] [CrossRef] [PubMed]
- Wharton, R.P.; Sonoda, J.; Lee, T.; Patterson, M.; Murata, Y. The Pumilio RNA-binding domain is also a translational regulator. Mol. Cell 1998, 1, 863–872. [Google Scholar] [CrossRef]
- Contreras, V.; Friday, A.J.; Morrison, J.K.; Hao, E.; Keiper, B.D. Cap-Independent translation promotes C. elegans germ cell apoptosis through Apaf-1/CED-4 in a caspase-dependent mechanism. PLoS ONE 2011, 6, e24444. [Google Scholar]
- Stebbins-Boaz, B.; Cao, Q.; de Moor, C.H.; Mendez, R.; Richter, J.D. Maskin is a CPEB-associated factor that transiently interacts with elF-4E. Mol. Cell 1999, 4, 1017–1027. [Google Scholar] [CrossRef]
- Mendez, R.; Barnard, D.; Richter, J.D. Differential mRNA translation and meiotic progression require Cdc2-mediated CPEB destruction. EMBO J. 2002, 21, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Minshall, N.; Reiter, M.H.; Weil, D.; Standart, N. CPEB interacts with an ovary-specific eIF4E and 4E-T in early Xenopus oocytes. J. Biol. Chem. 2007, 282, 37389–37401. [Google Scholar] [CrossRef]
- Pique, M.; Lopez, J.M.; Foissac, S.; Guigo, R.; Mendez, R. A combinatorial code for CPE-mediated translational control. Cell 2008, 132, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, G.; Miron, M.; Han, H.; Liu, N.; Magescas, J.; Tettweiler, G.; Frank, F.; Siddiqui, N.; Sonenberg, N.; Lasko, P. Mextli is a novel eukaryotic translation initiation factor 4E-binding protein that promotes translation in Drosophila melanogaster. Mol. Cell. Biol. 2013, 33, 2854–2864. [Google Scholar] [CrossRef] [PubMed]
- Cho, P.F.; Gamberi, C.; Cho-Park, Y.A.; Cho-Park, I.B.; Lasko, P.; Sonenberg, N. Cap-dependent translational inhibition establishes two opposing morphogen gradients in Drosophila embryos. Curr. Biol. 2006, 16, 2035–2041. [Google Scholar] [CrossRef] [PubMed]
- Hengartner, M.O.; Horvitz, H.R. Programmed cell death in Caenorhabditis elegans. Curr. Opin. Genet. Dev. 1994, 4, 581–586. [Google Scholar] [CrossRef]
- Schedl, T. Developmental genetics of the germ line. In C Elegans II.; Riddle, D.L., Blumenthal, T., Meyer, B.J., Priess, J.R., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997; pp. 241–269. [Google Scholar]
- Gumienny, T.L.; Lambie, E.; Hartwieg, E.; Horvitz, H.R.; Hengartner, M.O. Genetic control of programmed cell death in the Caenorhabditis elegans hermaphrodite germline. Development 1999, 126, 1011–1022. [Google Scholar]
- Metzstein, M.M.; Stanfield, G.M.; Horvitz, H.R. Genetics of programmed cell death in C. elegans: Past, present and future. Trends Genet. 1998, 14, 410–416. [Google Scholar] [CrossRef]
- Lee, M.H.; Hook, B.; Pan, G.; Kershner, A.M.; Merritt, C.; Seydoux, G.; Thomson, J.A.; Wickens, M.; Kimble, J. Conserved regulation of MAP kinase expression by PUF RNA-binding proteins. PLoS Genet. 2007, 3, e233. [Google Scholar] [CrossRef]
- Min, H.; Shim, Y.H.; Kawasaki, I. Loss of PGL-1 and PGL-3, members of a family of constitutive germ-granule components, promotes germline apoptosis in C. elegans. J. Cell Sci. 2016, 129, 341–353. [Google Scholar] [CrossRef]
- Subramaniam, K.; Seydoux, G. nos-1 and nos-2, two genes related to Drosophila nanos, regulate primordial germ cell development and survival in Caenorhabditis elegans. Development 1999, 126, 4861–4871. [Google Scholar]
- Boag, P.R.; Nakamura, A.; Blackwell, T.K. A conserved RNA-protein complex component involved in physiological germline apoptosis regulation in C. elegans. Development 2005, 132, 4975–4986. [Google Scholar] [CrossRef]
- Navarro, R.E.; Blackwell, T.K. Requirement for P granules and meiosis for accumulation of the germline RNA helicase CGH-1. Genesis 2005, 42, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Navarro, R.E.; Shim, E.Y.; Kohara, Y.; Singson, A.; Blackwell, T.K. cgh-1, a conserved predicted RNA helicase required for gametogenesis and protection from physiological germline apoptosis in C. elegans. Development 2001, 128, 3221–3232. [Google Scholar]
- Salinas, L.S.; Franco-Cea, A.; Lascarez-Lagunas, L.I.; Villanueva-Chimal, E.; Maldonado, E.; Navarro, R.E. Germ cell survival in C. elegans and C. remanei is affected when the DEAD box RNA helicases VBH-1 or Cre-VBH-1 are silenced. Genesis 2012, 50, 801–818. [Google Scholar] [CrossRef] [PubMed]
- Kritikou, E.A.; Milstein, S.; Vidalain, P.O.; Lettre, G.; Bogan, E.; Doukoumetzidis, K.; Gray, P.; Chappell, T.G.; Vidal, M.; Hengartner, M.O. C. elegans GLA-3 is a novel component of the MAP kinase MPK-1 signaling pathway required for germ cell survival. Genes Dev. 2006, 20, 2279–2292. [Google Scholar] [CrossRef] [PubMed]
- Gavis, E.R.; Lehmann, R. Localization of nanos RNA controls embryonic polarity. Cell 1992, 71, 301–313. [Google Scholar] [CrossRef]
- Forbes, A.; Lehmann, R. Nanos and Pumilio have critical roles in the development and function of Drosophila germline stem cells. Development 1998, 125, 679–690. [Google Scholar] [PubMed]
- Lai, F.; Zhou, Y.; Luo, X.; Fox, J.; King, M.L. Nanos1 functions as a translational repressor in the Xenopus germline. Mech. Dev. 2011, 128, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Chagnovich, D.; Lehmann, R. Poly(A)-independent regulation of maternal hunchback translation in the Drosophila embryo. Proc. Natl. Acad. Sci. USA 2001, 98, 11359–11364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Joza, N.; Tasdemir, E.; Maiuri, M.C.; Hengartner, M.; Abrams, J.M.; Tavernarakis, N.; Penninger, J.; Madeo, F.; Kroemer, G. No death without life: Vital functions of apoptotic effectors. Cell Death Differ. 2008, 15, 1113–1123. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.R.; Vernooy, S.Y.; Yu, H.; Yan, N.; Shi, Y.; Guo, M.; Hay, B.A. Multiple apoptotic caspase cascades are required in nonapoptotic roles for Drosophila spermatid individualization. PLoS Biol. 2004, 2, E15. [Google Scholar] [CrossRef] [PubMed]
- Henderson, M.A.; Cronland, E.; Dunkelbarger, S.; Contreras, V.; Strome, S.; Keiper, B.D. A germ line-specific isoform of eIF4E (IFE-1) is required for efficient translation of stored mRNAs and maturation of both oocytes and sperm. J. Cell Sci. 2009, 122, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Honarpour, N.; Du, C.; Richardson, J.A.; Hammer, R.E.; Wang, X.; Herz, J. Adult Apaf-1-deficient mice exhibit male infertility. Dev. Biol. 2000, 218, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Arama, E.; Agapite, J.; Steller, H. Caspase activity and a specific cytochrome C are required for sperm differentiation in Drosophila. Dev. Cell 2003, 4, 687–697. [Google Scholar] [CrossRef]
- Zermati, Y.; Mouhamad, S.; Stergiou, L.; Besse, B.; Galluzzi, L.; Boehrer, S.; Pauleau, A.L.; Rosselli, F.; D’Amelio, M.; Amendola, R.; et al. Nonapoptotic role for Apaf-1 in the DNA damage checkpoint. Mol. Cell 2007, 28, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Medvedev, S.; Yu, J.; Tang, L.C.; Agno, J.E.; Matzuk, M.M.; Schultz, R.M.; Hecht, N.B. Absence of the DNA-/RNA-binding protein MSY2 results in male and female infertility. Proc. Natl. Acad. Sci. USA 2005, 102, 5755–5760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evdokimova, V.; Tognon, C.; Ng, T.; Ruzanov, P.; Melnyk, N.; Fink, D.; Sorokin, A.; Ovchinnikov, L.P.; Davicioni, E.; Triche, T.J.; et al. Translational activation of snail1 and other developmentally regulated transcription factors by YB-1 promotes an epithelial-mesenchymal transition. Cancer Cell 2009, 15, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Clemens, M.J. Targets and mechanisms for the regulation of translation in malignant transformation. Oncogene 2004, 23, 3180–3188. [Google Scholar] [CrossRef] [Green Version]
- De Benedetti, A.; Graff, J.R. eIF-4E expression and its role in malignancies and metastases. Oncogene 2004, 23, 3189–3199. [Google Scholar] [CrossRef]
- Li, L.; Luo, Q.; Xie, Z.; Li, G.; Mao, C.; Liu, Y.; Wen, X.; Yin, N.; Cao, J.; Wang, J.; et al. Characterization of the Expression of the RNA Binding Protein eIF4G1 and Its Clinicopathological Correlation with Serous Ovarian Cancer. PLoS ONE 2016, 11, e0163447. [Google Scholar] [CrossRef]
- Silvera, D.; Arju, R.; Darvishian, F.; Levine, P.H.; Zolfaghari, L.; Goldberg, J.; Hochman, T.; Formenti, S.C.; Schneider, R.J. Essential role for eIF4GI overexpression in the pathogenesis of inflammatory breast cancer. Nat. Cell Biol. 2009, 11, 903–908. [Google Scholar] [CrossRef]
- Baker, C.C.; Fuller, M.T. Translational control of meiotic cell cycle progression and spermatid differentiation in male germ cells by a novel eIF4G homolog. Development 2007, 134, 2863–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrera, P.; Johnstone, O.; Nakamura, A.; Casanova, J.; Jackle, H.; Lasko, P. VASA mediates translation through interaction with a Drosophila yIF2 homolog. Mol. Cell 2000, 5, 181–187. [Google Scholar] [CrossRef]
- Ghosh, S.; Lasko, P. Loss-of-function analysis reveals distinct requirements of the translation initiation factors eIF4E, eIF4E-3, eIF4G and eIF4G2 in Drosophila spermatogenesis. PLoS ONE 2015, 10, e0122519. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Spycher, C.; Han, Z.S.; Rose, A.M.; Muller, F.; Avruch, J. TOR deficiency in C. elegans causes developmental arrest and intestinal atrophy by inhibition of mRNA translation. Curr. Biol. 2002, 12, 1448–1461. [Google Scholar] [CrossRef]
- Shen, R.; Weng, C.; Yu, J.; Xie, T. eIF4A controls germline stem cell self-renewal by directly inhibiting BAM function in the Drosophila ovary. Proc. Natl. Acad. Sci. USA 2009, 106, 11623–11628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri, A.; Keiper, B.D.; Kawasaki, I.; Fan, Y.; Kohara, Y.; Rhoads, R.E.; Strome, S. An isoform of eIF4E is a component of germ granules and is required for spermatogenesis in C. elegans. Development 2001, 128, 3899–3912. [Google Scholar]
- Keiper, B.D.; Lamphear, B.J.; Deshpande, A.M.; Jankowska-Anyszka, M.; Aamodt, E.J.; Blumenthal, T.; Rhoads, R.E. Functional characterization of five eIF4E isoforms in Caenorhabditis elegans. J. Biol. Chem. 2000, 275, 10590–10596. [Google Scholar] [CrossRef]
- Song, A.; Labella, S.; Korneeva, N.L.; Keiper, B.D.; Aamodt, E.J.; Zetka, M.; Rhoads, R.E. A C. elegans eIF4E-family member upregulates translation at elevated temperatures of mRNAs encoding MSH-5 and other meiotic crossover proteins. J. Cell Sci. 2010, 123, 2228–2237. [Google Scholar] [CrossRef]
- Cao, Q.; Padmanabhan, K.; Richter, J.D. Pumilio 2 controls translation by competing with eIF4E for 7-methyl guanosine cap recognition. RNA 2010, 16, 221–227. [Google Scholar] [CrossRef]
- Rodriguez, C.M.; Freire, M.A.; Camilleri, C.; Robaglia, C. The Arabidopsis thaliana cDNAs coding for eIF4E and eIF(iso)4E are not functionally equivalent for yeast complementation and are differentially expressed during plant development. Plant J. 1998, 13, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Ruffel, S.; Gallois, J.L.; Moury, B.; Robaglia, C.; Palloix, A.; Caranta, C. Simultaneous mutations in translation initiation factors eIF4E and eIF(iso)4E are required to prevent pepper veinal mottle virus infection of pepper. J. Gen. Virol. 2006, 87, 2089–2098. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, G.; Altmann, M.; Sierra, J.M.; Urlaub, H.; del Corral, R.D.; Schwartz, P.; Rivera-Pomar, R. Functional analysis of seven genes encoding eight translation initiation factor 4E (eIF4E) isoforms in Drosophila. Mech. Dev. 2005, 122, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.; Lee, K.; Maeder, D.L.; Jagus, R. Phylogenetic analysis of eIF4E-family members. BMC Evol. Biol. 2005, 5, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Patrick, R.M.; Browning, K.S. The eIF4F and eIFiso4F Complexes of Plants: An Evolutionary Perspective. Comp. Funct. Genom. 2012, 2012, 287814. [Google Scholar] [CrossRef] [PubMed]
- Patrick, R.M.; Mayberry, L.K.; Choy, G.; Woodard, L.E.; Liu, J.S.; White, A.; Mullen, R.A.; Tanavin, T.M.; Latz, C.A.; Browning, K.S. Two Arabidopsis loci encode novel eukaryotic initiation factor 4E isoforms that are functionally distinct from the conserved plant eukaryotic initiation factor 4E. Plant Physiol. 2014, 164, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Franklin-Dumont, T.M.; Chatterjee, C.; Wasserman, S.A.; Dinardo, S. A novel eIF4G homolog, Off-schedule, couples translational control to meiosis and differentiation in Drosophila spermatocytes. Development 2007, 134, 2851–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, A.; Sato, K.; Hanyu-Nakamura, K. Drosophila cup is an eIF4E binding protein that associates with Bruno and regulates oskar mRNA translation in oogenesis. Dev. Cell 2004, 6, 69–78. [Google Scholar] [CrossRef]
- Sengupta, M.S.; Low, W.Y.; Patterson, J.R.; Kim, H.M.; Traven, A.; Beilharz, T.H.; Colaiacovo, M.P.; Schisa, J.A.; Boag, P.R. ifet-1 is a broad-scale translational repressor required for normal P granule formation in C. elegans. J. Cell Sci. 2013, 126, 850–859. [Google Scholar] [CrossRef]
- Kawasaki, I.; Jeong, M.H.; Shim, Y.H. Regulation of sperm-specific proteins by IFE-1, a germline-specific homolog of eIF4E, in C. elegans. Mol. Cells 2011, 31, 191–197. [Google Scholar] [CrossRef]
- Mangio, R.S.; Votra, S.; Pruyne, D. The canonical eIF4E isoform of C. elegans regulates growth, embryogenesis, and germline sex-determination. Biol. Open 2015, 4, 843–851. [Google Scholar] [CrossRef]
- Friday, A.J.; Henderson, M.A.; Morrison, J.K.; Hoffman, J.L.; Keiper, B.D. Spatial and temporal translational control of germ cell mRNAs mediated by the eIF4E isoform IFE-1. J. Cell Sci. 2015, 128, 4487–4498. [Google Scholar] [CrossRef] [PubMed]
- Dinkova, T.D.; Keiper, B.D.; Korneeva, N.L.; Aamodt, E.J.; Rhoads, R.E. Translation of a small subset of Caenorhabditis elegans mRNAs is dependent on a specific eukaryotic translation initiation factor 4E isoform. Mol. Cell. Biol. 2005, 25, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Spike, C.A.; Coetzee, D.; Eichten, C.; Wang, X.; Hansen, D.; Greenstein, D.I. The TRIM-NHL Protein LIN-41 and the OMA RNA-Binding Proteins Antagonistically Control the Prophase-to-Metaphase Transition and Growth of Caenorhabditis elegans Oocytes. Genetics 2014, 198, 1535–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadler, T.W. Langman’s Essential Medical Embryology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2005; pp. 6–13, 24–26, 135–141. [Google Scholar]
- Larocca, R.A.; Abbink, P.; Peron, J.P.; Zanotto, P.M.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Nganga, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine protection against Zika virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, A.B.; Escribano-Romero, E.; Merino-Ramos, T.; Saiz, J.C.; Martin-Acebes, M.A. Stress responses in flavivirus-infected cells: Activation of unfolded protein response and autophagy. Front. Microbiol. 2014, 5, 266. [Google Scholar] [CrossRef] [PubMed]
- Elgner, F.; Sabino, C.; Basic, M.; Ploen, D.; Grunweller, A.; Hildt, E. Inhibition of Zika Virus Replication by Silvestrol. Viruses 2018, 10, 149. [Google Scholar] [CrossRef]
- Mao, H.; McMahon, J.J.; Tsai, Y.H.; Wang, Z.; Silver, D.L. Haploinsufficiency for Core Exon Junction Complex Components Disrupts Embryonic Neurogenesis and Causes p53-Mediated Microcephaly. PLoS Genet. 2016, 12, e1006282. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.J. Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses. Arch. Virol. 2007, 152, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Komar, A.A.; Mazumder, B.; Merrick, W.C. A new framework for understanding IRES-mediated translation. Gene 2010, 502, 75–86. [Google Scholar] [CrossRef]
- Hernandez, G.; Proud, C.G.; Preiss, T.; Parsyan, A. On the Diversification of the Translation Apparatus across Eukaryotes. Comp. Funct. Genom. 2012, 2012, 256848. [Google Scholar] [CrossRef]
- Sriram, A.; Bohlen, J.; Teleman, A.A. Translation acrobatics: How cancer cells exploit alternate modes of translational initiation. EMBO Rep. 2018, 19, 17. [Google Scholar] [CrossRef] [PubMed]
- Walters, B.; Thompson, S.R. Cap-Independent Translational Control of Carcinogenesis. Front. Oncol. 2016, 6, 128. [Google Scholar] [CrossRef] [PubMed]
- Braunstein, S.; Karpisheva, K.; Pola, C.; Goldberg, J.; Hochman, T.; Yee, H.; Cangiarella, J.; Arju, R.; Formenti, S.C.; Schneider, R.J. A hypoxia-controlled cap-dependent to cap-independent translation switch in breast cancer. Mol. Cell 2007, 28, 501–512. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keiper, B.D. Cap-Independent mRNA Translation in Germ Cells. Int. J. Mol. Sci. 2019, 20, 173. https://doi.org/10.3390/ijms20010173
Keiper BD. Cap-Independent mRNA Translation in Germ Cells. International Journal of Molecular Sciences. 2019; 20(1):173. https://doi.org/10.3390/ijms20010173
Chicago/Turabian StyleKeiper, Brett D. 2019. "Cap-Independent mRNA Translation in Germ Cells" International Journal of Molecular Sciences 20, no. 1: 173. https://doi.org/10.3390/ijms20010173
APA StyleKeiper, B. D. (2019). Cap-Independent mRNA Translation in Germ Cells. International Journal of Molecular Sciences, 20(1), 173. https://doi.org/10.3390/ijms20010173