In Situ Monitoring of Bacteria under Antimicrobial Stress Using 31P Solid-State NMR

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
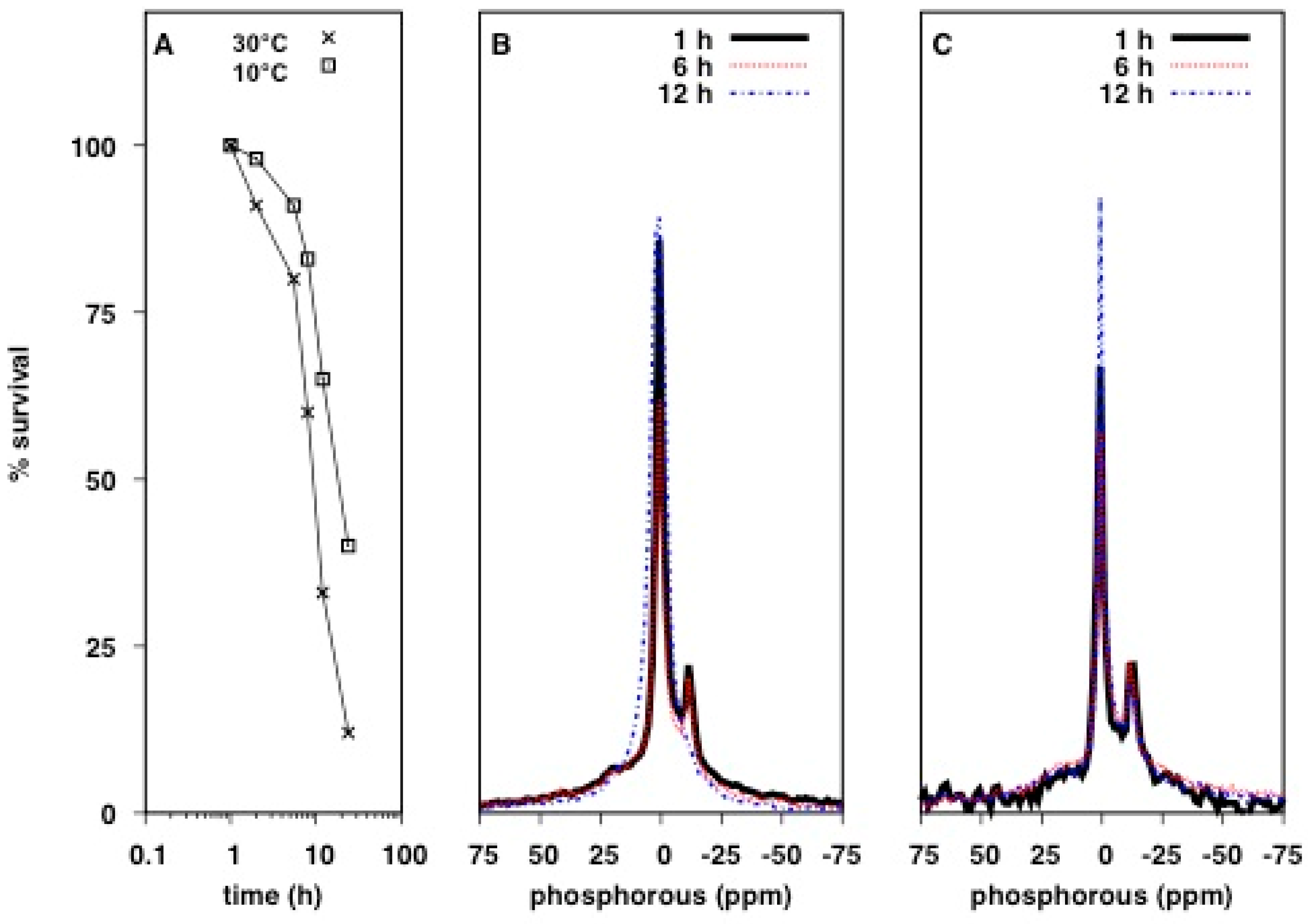
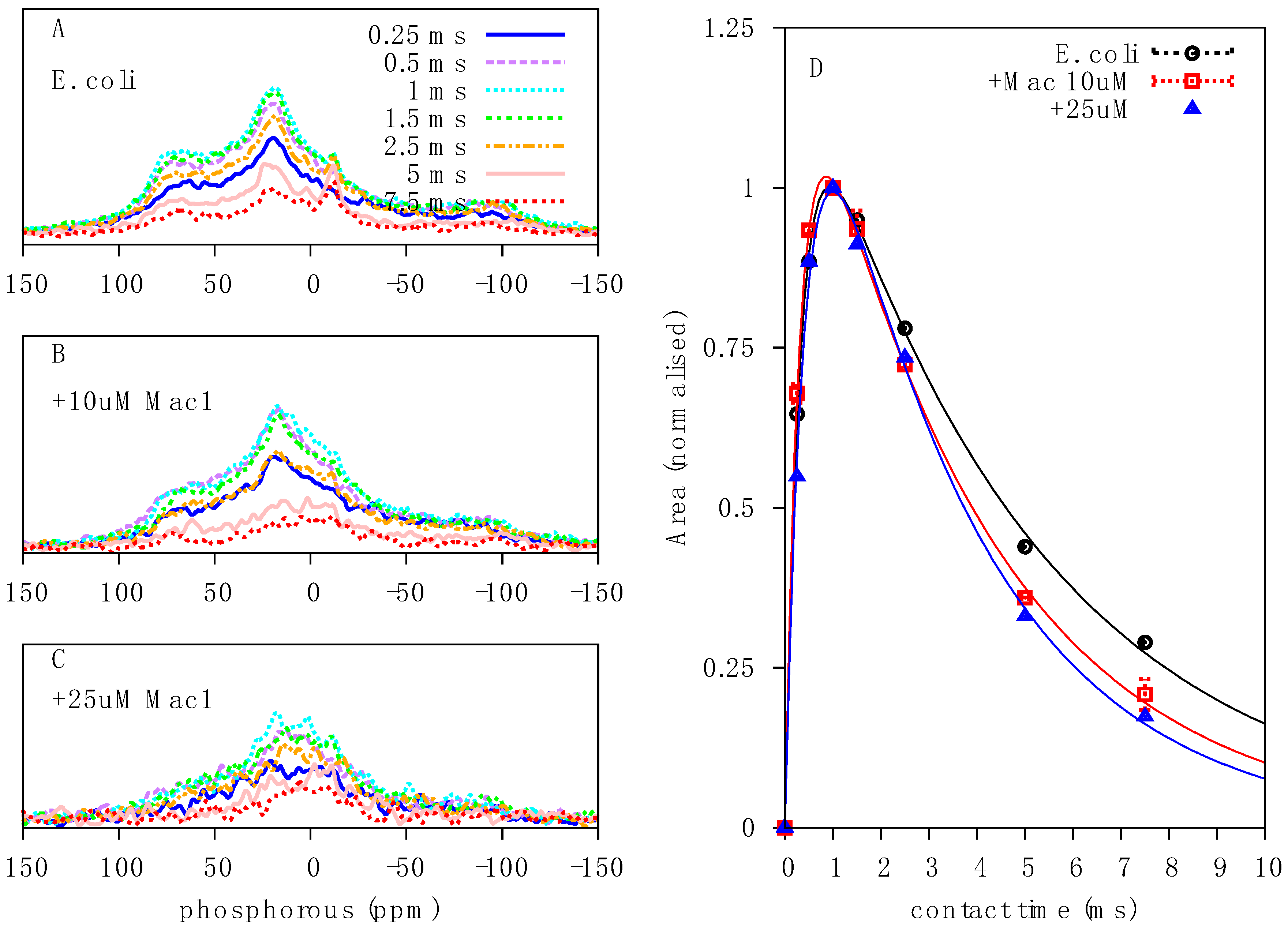
2.1. Solid-State 31P NMR Experiments on Intact E. coli Bacteria
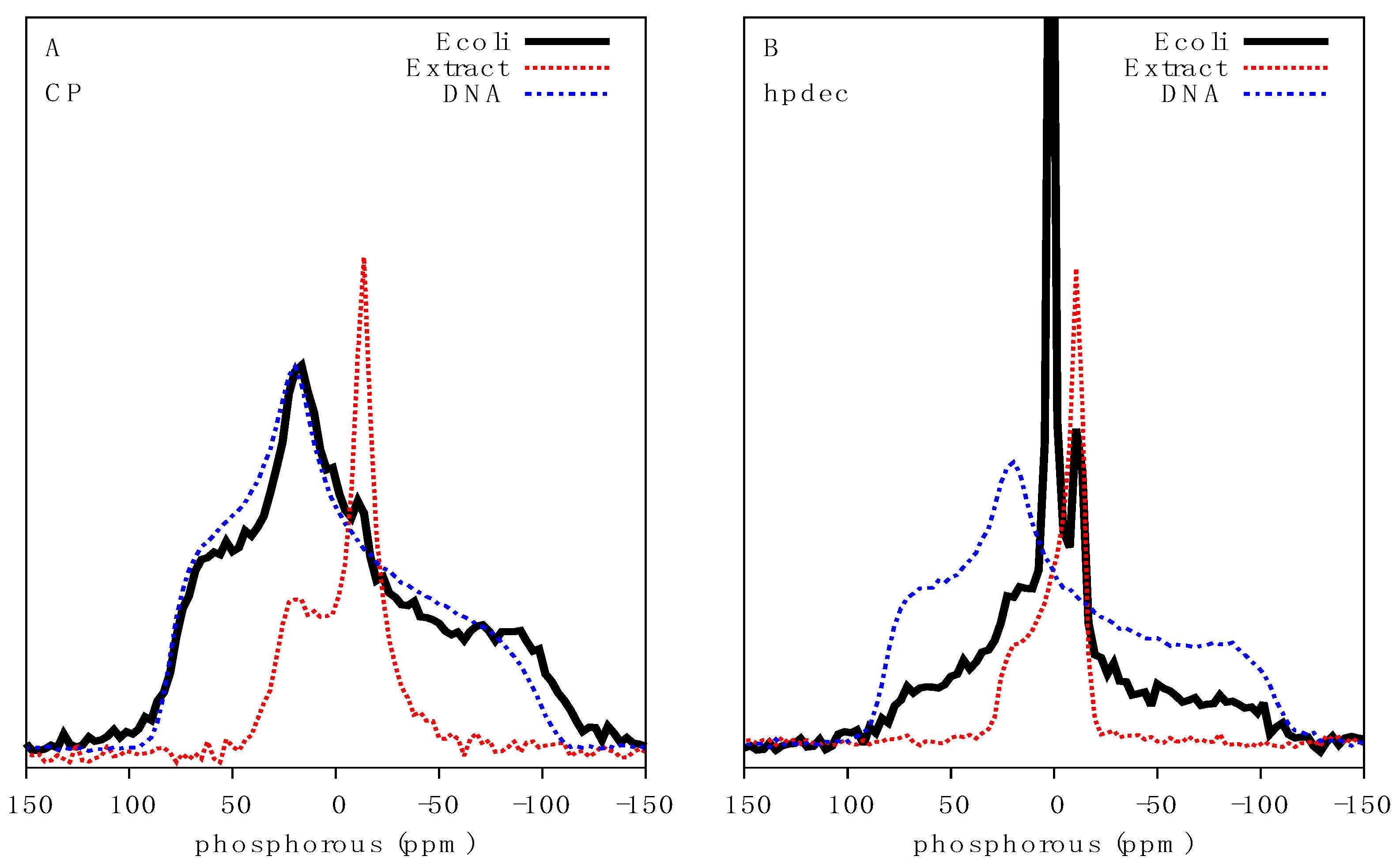
2.2. Qualitative Analysis of 31P Lineshapes in Intact Bacteria
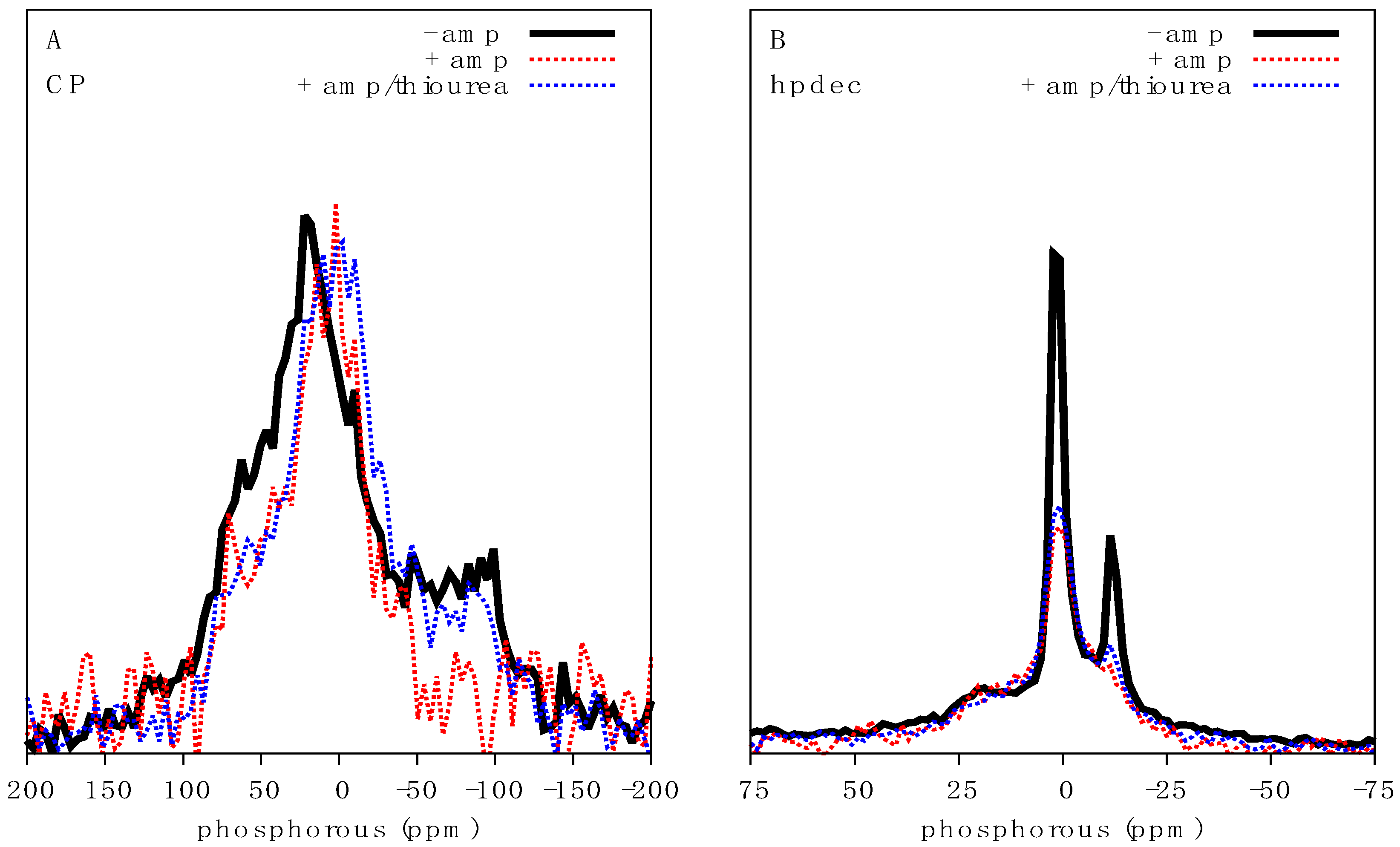
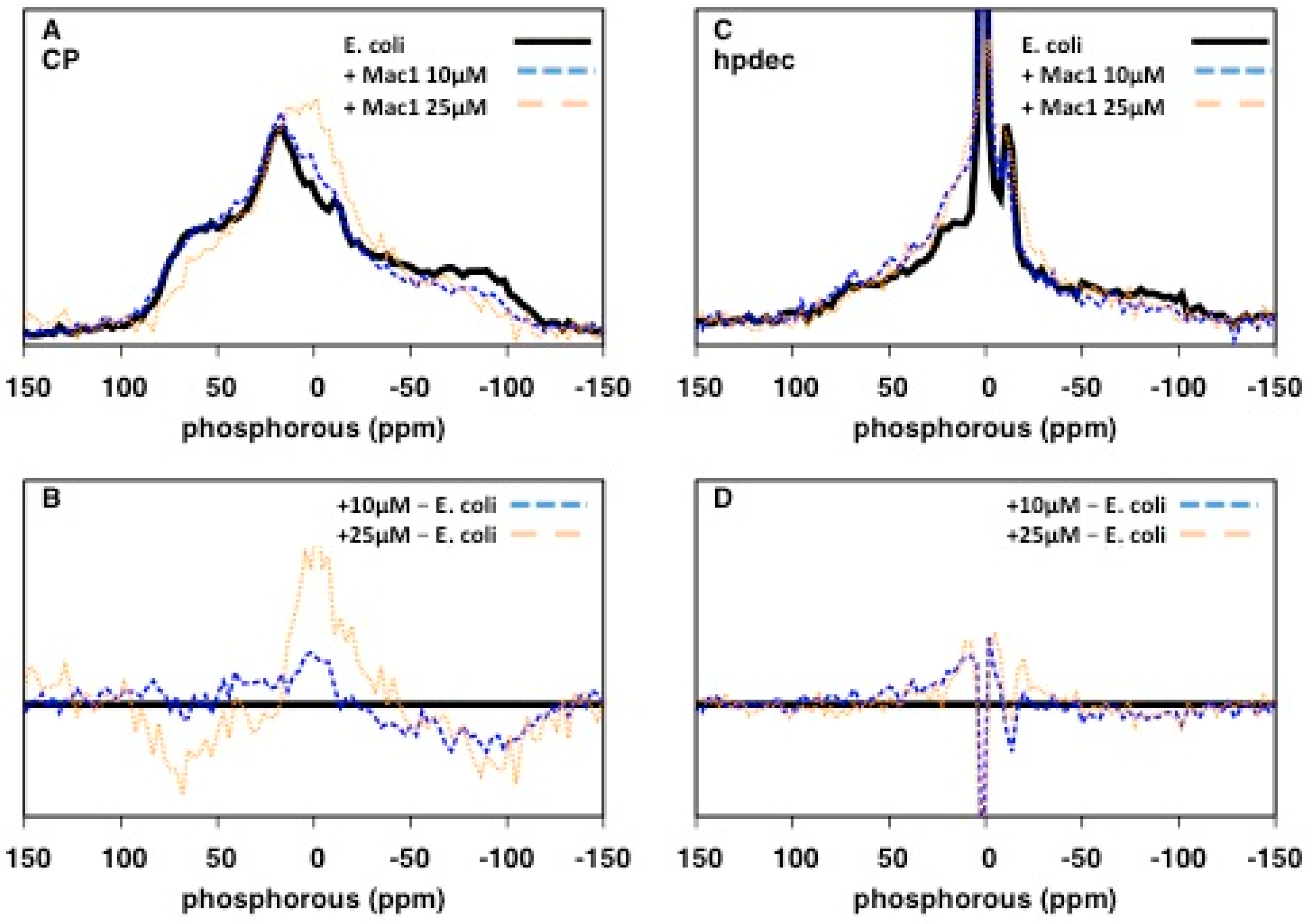
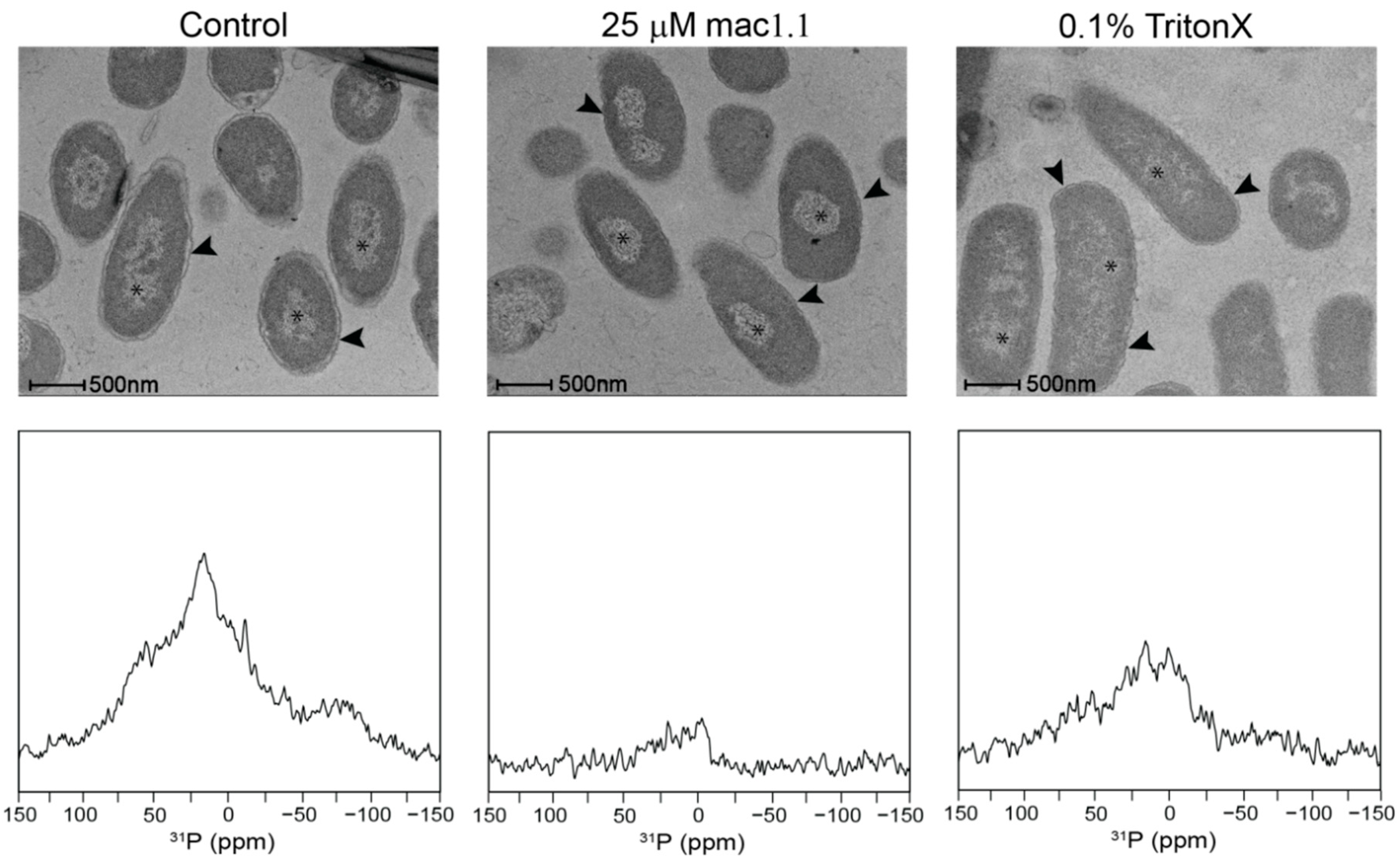
2.3. Characterisation of the Effects of Antimicrobial Agents on Bacteria
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Bacterial Growth
4.3. Bacterial Survival
4.4. E. coli Total Lipid Extract Sample Preparation
4.5. Solid-State NMR Spectroscopy
4.6. Electron Microscopy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NMR | Nuclear Magnetic Resonance |
| CP | Cross-Polarization |
| CSA | Chemical Shift Anisotropy |
| AMP | Antimicrobial Peptide |
| Mac1.1 | Maculatin 1.1 |
| DNA | Deoxyribonucleic Acid |
| CFU | Colony Forming Unit |
References
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.A.; Gagne, E.; Gehman, J.D.; Whitwell, T.C.; Separovic, F. Dye-release assay for investigation of antimicrobial peptide activity in a competitive lipid environment. Eur. Biophys. J. EBJ 2014, 43, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.A.; Separovic, F. Progression of NMR studies of membrane-active peptides from lipid bilayers to live cells. J. Magn. Reson. 2015, 253, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.A.; Separovic, F. How Membrane-Active Peptides Get into Lipid Membranes. Acc. Chem. Res. 2016, 4, 1130–1138. [Google Scholar] [CrossRef]
- Melo, M.N.; Ferre, R.; Castanho, M.A.R.B. Antimicrobial peptides: Linking partition, activity and high membrane-bound concentrations. Nat. Rev. Microbiol. 2009, 7, 245–250. [Google Scholar] [CrossRef]
- Chia, B.C.S.; Carver, J.A.; Mulhern, T.D.; Bowie, J.H. Maculatin 1.1, an anti-microbial peptide from the Australian tree frog, Litoria genimaculata—Solution structure and biological activity. Eur. J. Biochem. 2000, 267, 1894–1908. [Google Scholar] [CrossRef]
- Rozek, T.; Waugh, R.J.; Steinborner, S.T.; Bowie, J.H.; Tyler, M.J.; Wallace, J.C. The maculatin peptides from the skin glands of the tree frog Litoria genimaculata: A comparison of the structures and antibacterial activities of maculatin 1.1 and caerin 1.1. J. Pept. Sci. 1998, 4, 111–115. [Google Scholar] [CrossRef]
- Sani, M.A.; Whitwell, T.C.; Gehman, J.D.; Robins-Browne, R.M.; Pantarat, N.; Attard, T.J.; Reynolds, E.C.; O’Brien-Simpson, N.M.; Separovic, F. Maculatin 1.1 disrupts Staphylococcus aureus lipid membranes via a pore mechanism. Antimicrob. Agents Chemother. 2013, 57, 3593–3600. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.A.; Henriques, S.T.; Weber, D.; Separovic, F. Bacteria May Cope Differently from Similar Membrane Damage Caused by the Australian Tree Frog Antimicrobial Peptide Maculatin 1.1. J. Biol. Chem. 2015, 290, 19853–19862. [Google Scholar] [CrossRef] [PubMed]
- Scocchi, M.; Mardirossian, M.; Runti, G.; Benincasa, M. Non-Membrane Permeabilizing Modes of Action of Antimicrobial Peptides on Bacteria. Curr. Top. Med. Chem. 2016, 16, 76–88. [Google Scholar] [CrossRef]
- Freedberg, D.I.; Selenko, P. Live Cell NMR. Annu. Rev. Biophys. 2014, 43, 171–192. [Google Scholar] [CrossRef] [PubMed]
- Burz, D.S.; Dutta, K.; Cowburn, D.; Shekhtman, A. In-cell NMR for protein-protein interactions (STINT-NMR). Nat. Protoc. 2006, 1, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, S.; DeMott, C.M.; Reverdatto, S.; Burz, D.S.; Shekhtman, A. Total Cellular RNA Modulates Protein Activity. Biochemistry 2016, 55, 4568–4573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahab, N.; Flett, F.; Oliver, S.G.; Butler, P.R. Growth rate control of protein and nucleic acid content in Streptomyces coelicolor A3(2) and Escherichia coli B/r. Microbiology 1996, 142 Pt 8, 1927–1935. [Google Scholar] [CrossRef]
- Smirnova, G.; Muzyka, N.; Lepekhina, E.; Oktyabrsky, O. Roles of the glutathione- and thioredoxin-dependent systems in the Escherichia coli responses to ciprofloxacin and ampicillin. Arch. Microbiol. 2016, 198, 913–921. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, S5–S16. [Google Scholar] [CrossRef]
- Luchinat, E.; Banci, L. In-cell NMR: A topical review. IUCrJ 2017, 4, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Diverdi, J.A.; Opella, S.J. Phosphorus-31 Nuclear Magnetic-Resonance of fd Virus. Biochemistry 1981, 20, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Amani, J.; Barjini, K.A.; Moghaddam, M.M.; Asadi, A. In vitro synergistic effect of the CM11 antimicrobial peptide in combination with common antibiotics against clinical isolates of six species of multidrug-resistant pathogenic bacteria. Protein Pept. Lett. 2015, 22, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Z.; Li, Z.; Li, X.L.; Tian, Y.M.; Fan, Y.Z.; Yu, C.H.; Zhou, B.L.; Liu, Y.; Xiang, R.; Yang, L. Synergistic effects of antimicrobial peptide DP7 combined with antibiotics against multidrug-resistant bacteria. Drug Des. Dev. Ther. 2017, 11, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Laadhari, M.; Arnold, A.A.; Gravel, A.E.; Separovic, F.; Marcotte, I. Interaction of the antimicrobial peptides caerin 1.1 and aurein 1.2 with intact bacteria by 2H solid-state NMR. BBA-Biomembranes 2016, 1858, 2959–2964. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.A.; Separovic, F.; Gehman, J.D. Disentanglement of Heterogeneous Dynamics in Mixed Lipid Systems. Biophys. J. 2011, 100, L40–L42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sani, M.A.; Dufourc, E.J.; Grobner, G. How does the Bax-α1 targeting sequence interact with mitochondrial membranes? The role of cardiolipin. Biochim. Biophys. Acta 2009, 1788, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejski, W.; Klinowski, J. Kinetics of cross-polarization in solid-state NMR: A guide for chemists. Chem. Rev. 2002, 102, 613–628. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Overall, S.A.; Zhu, S.; Hanssen, E.; Separovic, F.; Sani, M.-A. In Situ Monitoring of Bacteria under Antimicrobial Stress Using 31P Solid-State NMR. Int. J. Mol. Sci. 2019, 20, 181. https://doi.org/10.3390/ijms20010181
Overall SA, Zhu S, Hanssen E, Separovic F, Sani M-A. In Situ Monitoring of Bacteria under Antimicrobial Stress Using 31P Solid-State NMR. International Journal of Molecular Sciences. 2019; 20(1):181. https://doi.org/10.3390/ijms20010181
Chicago/Turabian StyleOverall, Sarah A., Shiying Zhu, Eric Hanssen, Frances Separovic, and Marc-Antoine Sani. 2019. "In Situ Monitoring of Bacteria under Antimicrobial Stress Using 31P Solid-State NMR" International Journal of Molecular Sciences 20, no. 1: 181. https://doi.org/10.3390/ijms20010181
APA StyleOverall, S. A., Zhu, S., Hanssen, E., Separovic, F., & Sani, M. -A. (2019). In Situ Monitoring of Bacteria under Antimicrobial Stress Using 31P Solid-State NMR. International Journal of Molecular Sciences, 20(1), 181. https://doi.org/10.3390/ijms20010181