Ribosome Structure, Function, and Early Evolution



Abstract
:1. Introduction
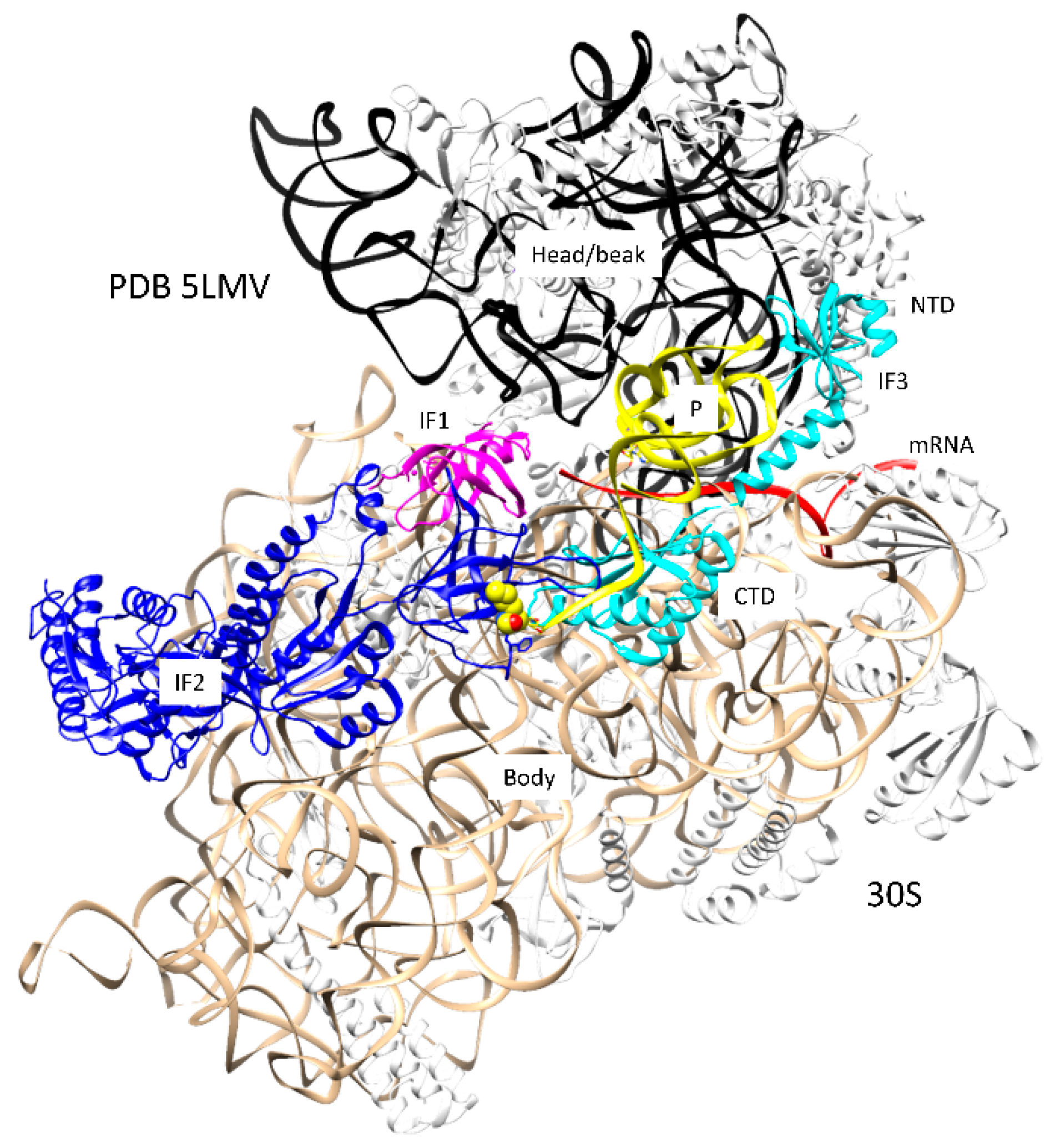
2. Initiation of Translation
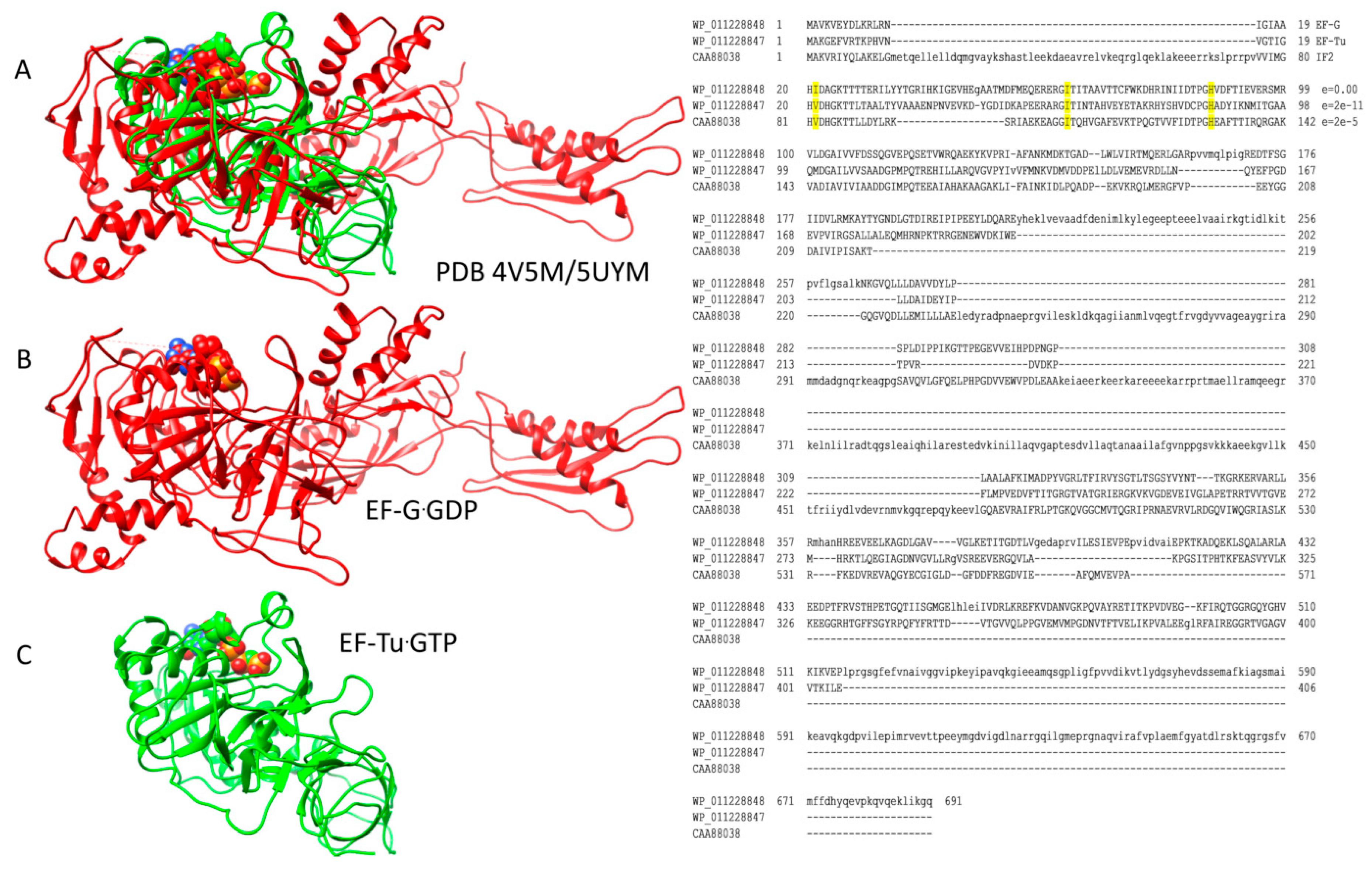
2.1. Homologous GTPases in Initiation and Elongation of Translation
2.2. Mechanism of Initiation
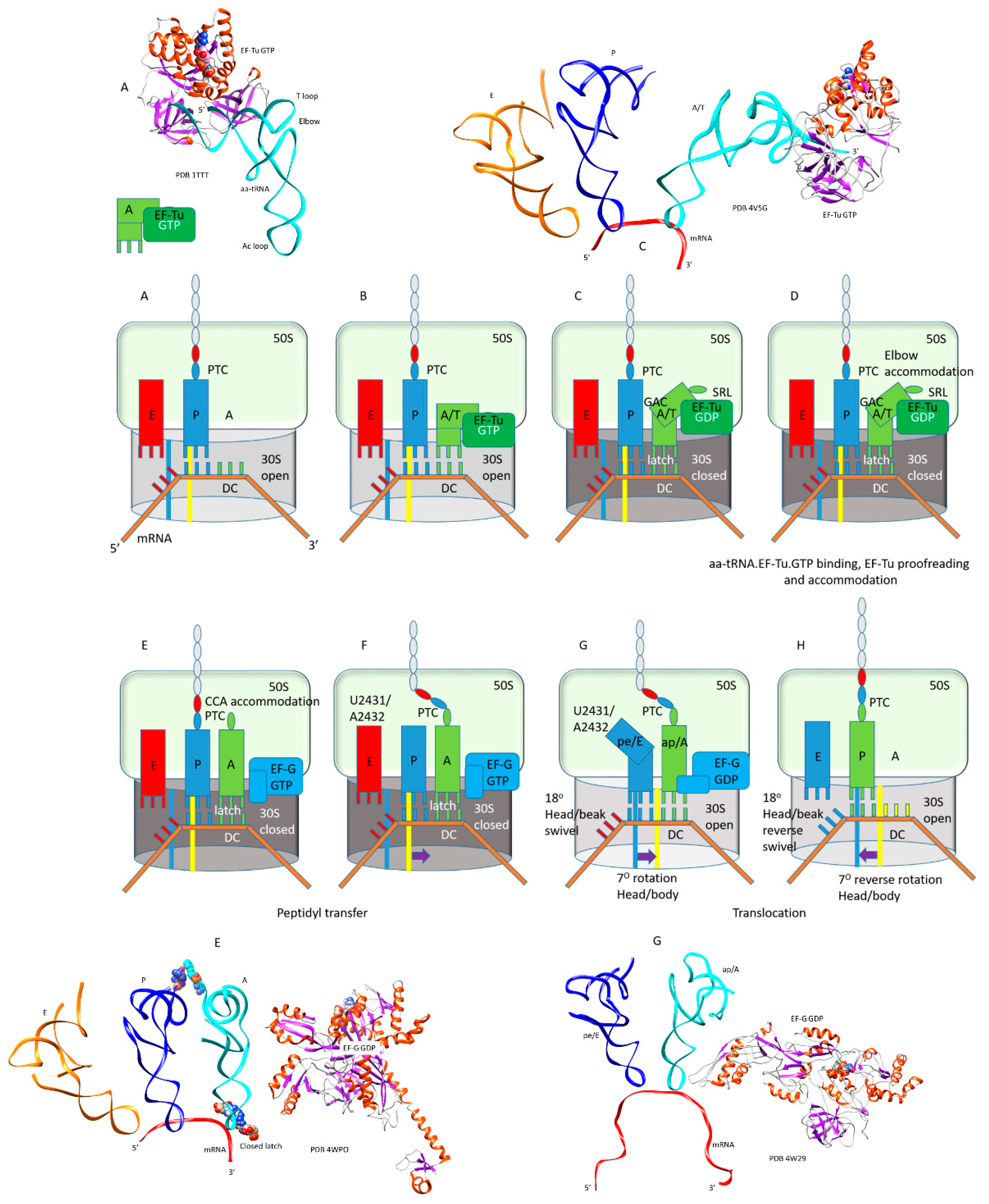
3. Elongation of Translation
3.1. Molecular Motor
3.2. tRNA as a Relatively Stiff Adapter
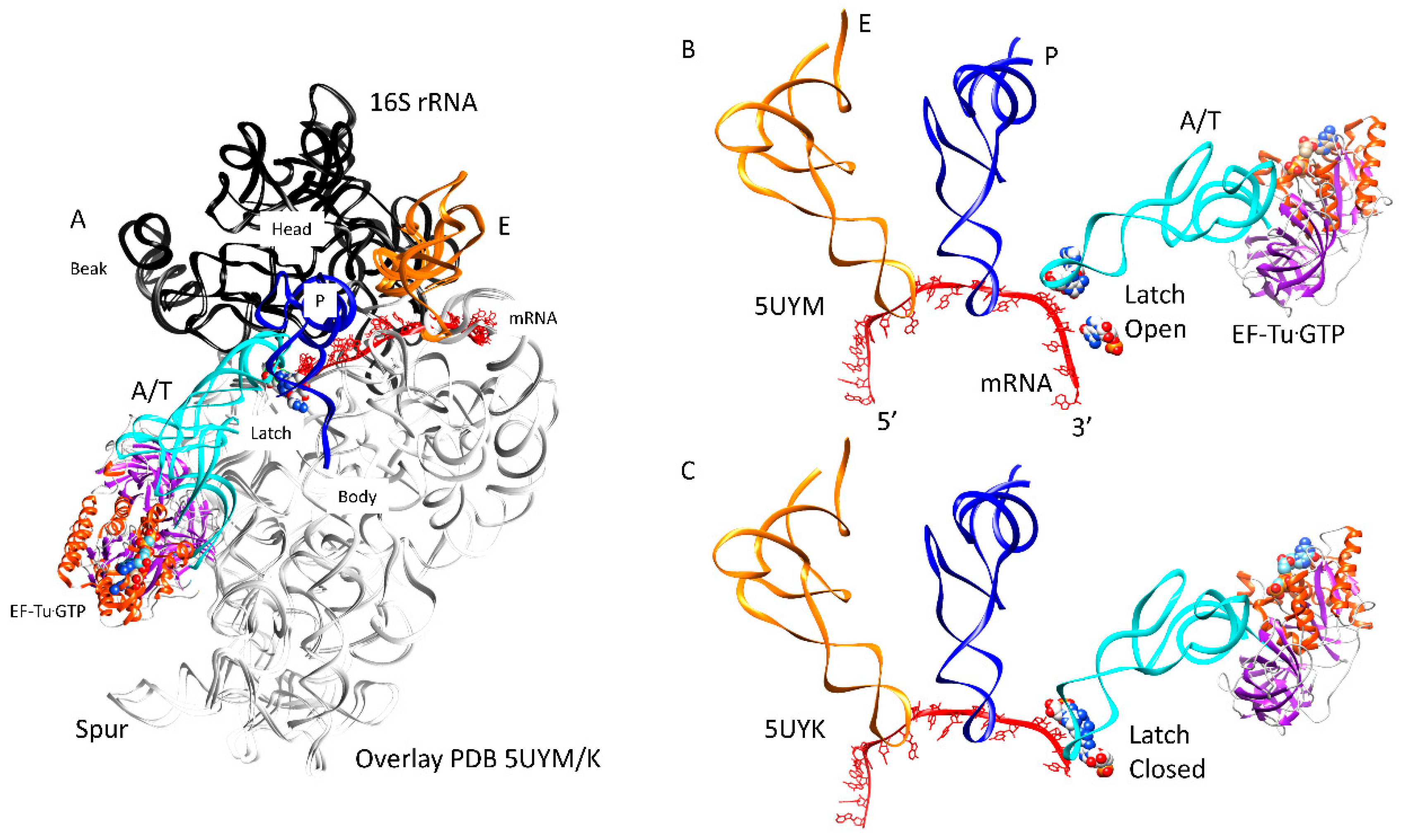
3.3. tRNA Entry
3.4. Forming the Accurate Codon-Anticodon Latch and Closing the 30S Subunit Conformation
3.5. Accommodation
3.6. Peptide Bond Formation
3.7. Translocation
3.8. EF-G·GTP/GDP in Translocation
3.9. 30S-50S Intersubunit Bridges in Translocation
3.10. Ratchet Pawls
3.11. Kink-Turns and Micro-Motions
4. Evolution of Translation
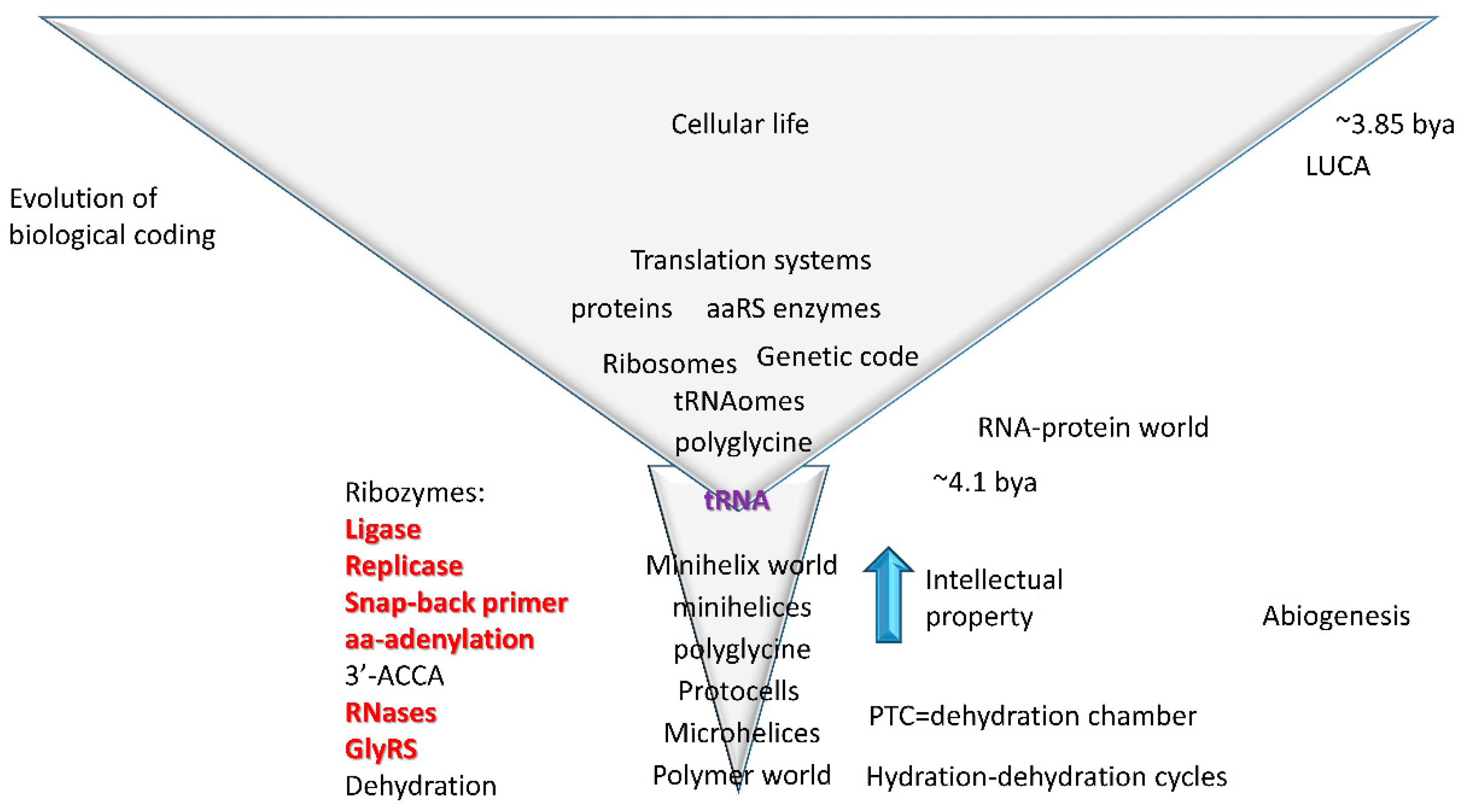
4.1. tRNA Evolution
4.2. tRNA as Core Evolutionary Intellectual Property
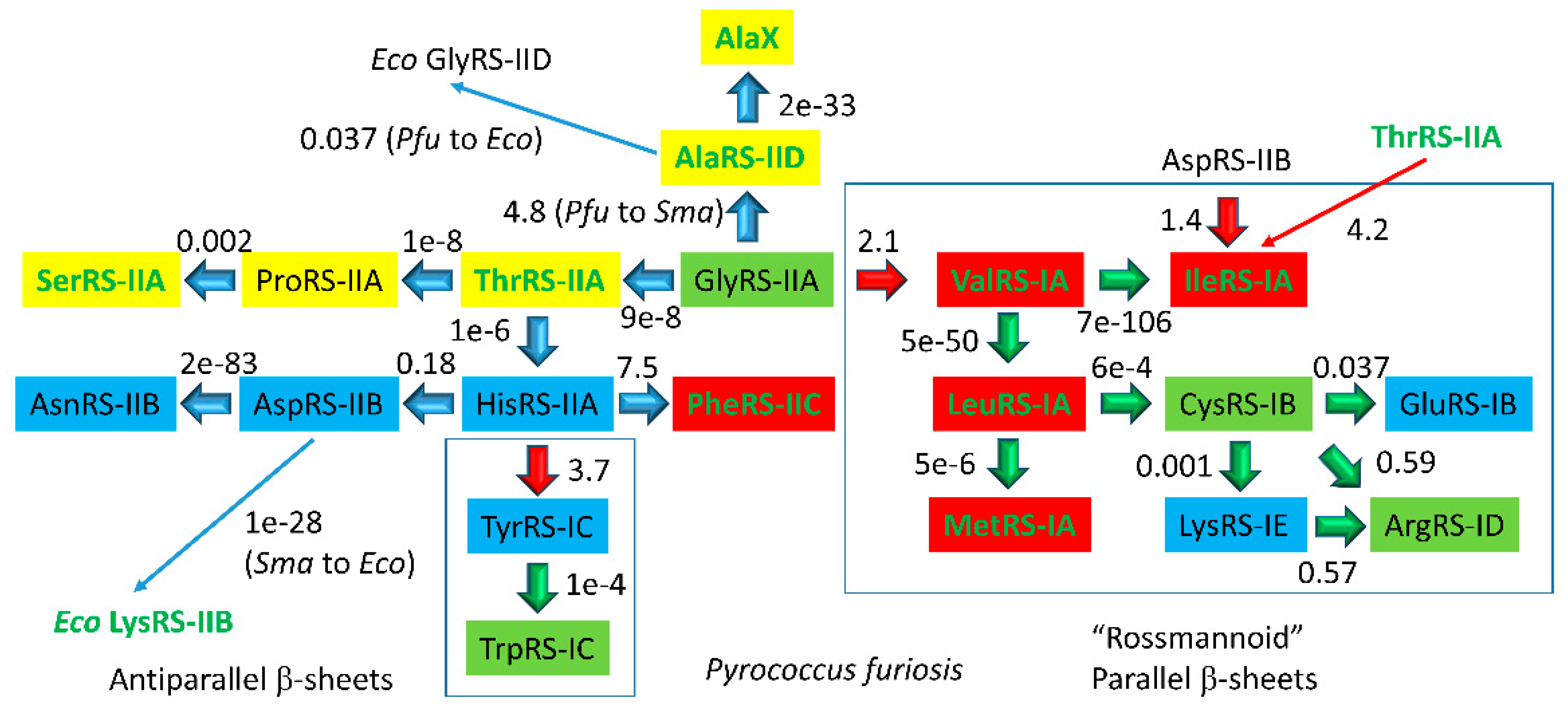
4.3. Aminoacyl-tRNA Synthetase Evolution
4.4. rRNA Evolution
4.5. Evolution of the Genetic Code
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| aaRS | Aminoacyl-tRNA synthetase (i.e., GlyRS) |
| A-site | Aminoacyl site |
| As | Acceptor stems |
| ASL | Anticodon stem loop |
| Ac loop | Anticodon loop |
| EF | Elongation Factor |
| E-site | Exit site |
| IF | Initiation Factor |
| LUCA | Last universal common (cellular) ancestor |
| P-site | Peptidyl site |
| PTC | Peptidyl Transferase Center |
| SRL | Sarcin-ricin loop |
| T loop | T loop or TΨC loop |
| V loop | Variable loop |
References
- Laursen, B.S.; Sorensen, H.P.; Mortensen, K.K.; Sperling-Petersen, H.U. Initiation of protein synthesis in bacteria. Microbiol. Mol. Biol. Rev. 2005, 69, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V.; Fischer, N.; Maracci, C.; Stark, H. Ribosome dynamics during decoding. Philos. Trans. R Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Maracci, C.; Rodnina, M.V. Review: Translational GTPases. Biopolymers 2016, 105, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorhees, R.M.; Ramakrishnan, V. Structural basis of the translational elongation cycle. Annu. Rev. Biochem. 2013, 82, 203–236. [Google Scholar] [CrossRef]
- Agirrezabala, X.; Frank, J. Elongation in translation as a dynamic interaction among the ribosome, tRNA, and elongation factors EF-G and EF-Tu. Q. Rev. Biophys. 2009, 42, 159–200. [Google Scholar] [CrossRef]
- Sergiev, P.V.; Bogdanov, A.A.; Dontsova, O.A. How can elongation factors EF-G and EF-Tu discriminate the functional state of the ribosome using the same binding site? FEBS Lett. 2005, 579, 5439–5442. [Google Scholar] [CrossRef] [Green Version]
- Rodnina, M.V.; Gromadski, K.B.; Kothe, U.; Wieden, H.J. Recognition and selection of tRNA in translation. FEBS Lett. 2005, 579, 938–942. [Google Scholar] [CrossRef]
- Holtkamp, W.; Wintermeyer, W.; Rodnina, M.V. Synchronous tRNA movements during translocation on the ribosome are orchestrated by elongation factor G and GTP hydrolysis. Bioessays 2014, 36, 908–918. [Google Scholar] [CrossRef] [Green Version]
- Rodnina, M.V.; Wintermeyer, W. The ribosome as a molecular machine: The mechanism of tRNA-mRNA movement in translocation. Biochem. Soc. Trans. 2011, 39, 658–662. [Google Scholar] [CrossRef]
- Ling, C.; Ermolenko, D.N. Structural insights into ribosome translocation. Wiley Int. Rev. RNA 2016, 7, 620–636. [Google Scholar] [CrossRef] [Green Version]
- Frank, J. Intermediate states during mRNA-tRNA translocation. Curr. Opin. Struct. Biol. 2012, 22, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Kavaliauskas, D.; Chen, C.; Liu, W.; Cooperman, B.S.; Goldman, Y.E.; Knudsen, C.R. Structural dynamics of translation elongation factor Tu during aa-tRNA delivery to the ribosome. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mondal, D.; Warshel, A. EF-Tu and EF-G are activated by allosteric effects. Proc. Natl. Acad. Sci. USA 2018, 115, 3386–3391. [Google Scholar] [CrossRef] [PubMed]
- Salsi, E.; Farah, E.; Netter, Z.; Dann, J.; Ermolenko, D.N. Movement of elongation factor G between compact and extended conformations. J. Mol. Biol. 2015, 427, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.G.; Lin, J.; Steitz, T.A. Elongation factor 4 remodels the A-site tRNA on the ribosome. Proc. Natl. Acad. Sci. USA 2016, 113, 4994–4999. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Alonso, J.P.; Fabbretti, A.; Kaminishi, T.; Iturrioz, I.; Brandi, L.; Gil-Carton, D.; Gualerzi, C.O.; Fucini, P.; Connell, S.R. Structure of a 30S pre-initiation complex stalled by GE81112 reveals structural parallels in bacterial and eukaryotic protein synthesis initiation pathways. Nucleic Acids Res. 2017, 45, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Llacer, J.L.; Wimberly, B.T.; Kieft, J.S.; Ramakrishnan, V. Large-Scale Movements of IF3 and tRNA during Bacterial Translation Initiation. Cell 2016, 167, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Yusupova, G.Z.; Yusupov, M.M.; Cate, J.H.; Noller, H.F. The path of messenger RNA through the ribosome. Cell 2001, 106, 233–241. [Google Scholar] [CrossRef]
- Yusupova, G.; Jenner, L.; Rees, B.; Moras, D.; Yusupov, M. Structural basis for messenger RNA movement on the ribosome. Nature 2006, 444, 391–394. [Google Scholar] [CrossRef]
- Ling, C.; Ermolenko, D.N. Initiation factor 2 stabilizes the ribosome in a semirotated conformation. Proc. Natl. Acad. Sci. USA 2015, 112, 15874–15879. [Google Scholar] [CrossRef]
- Liu, T.; Kaplan, A.; Alexander, L.; Yan, S.; Wen, J.D.; Lancaster, L.; Wickersham, C.E.; Fredrick, K.; Noller, H.; Tinoco, I.; et al. Direct measurement of the mechanical work during translocation by the ribosome. Elife 2014, 3, e03406. [Google Scholar] [CrossRef] [PubMed]
- Ratje, A.H.; Loerke, J.; Mikolajka, A.; Brunner, M.; Hildebrand, P.W.; Starosta, A.L.; Donhofer, A.; Connell, S.R.; Fucini, P.; Mielke, T.; et al. Head swivel on the ribosome facilitates translocation by means of intra-subunit tRNA hybrid sites. Nature 2010, 468, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Gao, H.; Sengupta, J.; Gao, N.; Taylor, D.J. The process of mRNA-tRNA translocation. Proc. Natl. Acad. Sci. USA 2007, 104, 19671–19678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlgemuth, I.; Pohl, C.; Rodnina, M.V. Optimization of speed and accuracy of decoding in translation. EMBO J. 2010, 29, 3701–3709. [Google Scholar] [CrossRef] [Green Version]
- Gromadski, K.B.; Rodnina, M.V. Kinetic determinants of high-fidelity tRNA discrimination on the ribosome. Mol. Cell. 2004, 13, 191–200. [Google Scholar] [CrossRef]
- Rodnina, M.V.; Wintermeyer, W. Fidelity of aminoacyl-tRNA selection on the ribosome: Kinetic and structural mechanisms. Annu. Rev. Biochem. 2001, 70, 415–435. [Google Scholar] [CrossRef]
- Pak, D.; Root-Bernstein, R.; Burton, Z.F. tRNA structure and evolution and standardization to the three nucleotide genetic code. Transcription 2017, 8, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Agirrezabala, X.; Lei, J.; Bouakaz, L.; Brunelle, J.L.; Ortiz-Meoz, R.F.; Green, R.; Sanyal, S.; Ehrenberg, M.; Frank, J. Recognition of aminoacyl-tRNA: A common molecular mechanism revealed by cryo-EM. EMBO J. 2008, 27, 3322–3331. [Google Scholar] [CrossRef]
- Zhou, J.; Lancaster, L.; Donohue, J.P.; Noller, H.F. Crystal structures of EF-G-ribosome complexes trapped in intermediate states of translocation. Science 2013, 340, 1236086. [Google Scholar] [CrossRef]
- Noel, J.K.; Whitford, P.C. How EF-Tu can contribute to efficient proofreading of aa-tRNA by the ribosome. Nat. Commun. 2016, 7, 13314. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.; Yang, H.; Whitford, P.C. How the Ribosomal A-Site Finger Can Lead to tRNA Species-Dependent Dynamics. J. Phys. Chem. B 2017, 121, 2767–2775. [Google Scholar] [CrossRef]
- Whitford, P.C.; Sanbonmatsu, K.Y. Simulating movement of tRNA through the ribosome during hybrid-state formation. J. Chem. Phys. 2013, 139, 121919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Perrier, J.; Whitford, P.C. Disorder guides domain rearrangement in elongation factor Tu. Proteins 2018, 86, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.; Kjeldgaard, M.; Thirup, S.; Polekhina, G.; Reshetnikova, L.; Clark, B.F.; Nyborg, J. Crystal structure of the ternary complex of Phe-tRNAPhe, EF-Tu, and a GTP analog. Science 1995, 270, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu, K.Y.; Joseph, S.; Tung, C.S. Simulating movement of tRNA into the ribosome during decoding. Proc. Natl. Acad. Sci. USA 2005, 102, 15854–15859. [Google Scholar] [CrossRef] [Green Version]
- Whitford, P.C.; Onuchic, J.N.; Sanbonmatsu, K.Y. Connecting energy landscapes with experimental rates for aminoacyl-tRNA accommodation in the ribosome. J. Am. Chem. Soc. 2010, 132, 13170–13171. [Google Scholar] [CrossRef]
- Whitford, P.C.; Geggier, P.; Altman, R.B.; Blanchard, S.C.; Onuchic, J.N.; Sanbonmatsu, K.Y. Accommodation of aminoacyl-tRNA into the ribosome involves reversible excursions along multiple pathways. RNA 2010, 16, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ieong, K.W.; Uzun, U.; Selmer, M.; Ehrenberg, M. Two proofreading steps amplify the accuracy of genetic code translation. Proc. Natl. Acad. Sci. USA 2016, 113, 13744–13749. [Google Scholar] [CrossRef]
- Aqvist, J.; Kamerlin, S.C. The conformation of a catalytic loop is central to GTPase activity on the ribosome. Biochemistry 2015, 54, 546–556. [Google Scholar] [CrossRef]
- Shi, X.; Khade, P.K.; Sanbonmatsu, K.Y.; Joseph, S. Functional role of the sarcin-ricin loop of the 23S rRNA in the elongation cycle of protein synthesis. J. Mol. Biol. 2012, 419, 125–138. [Google Scholar] [CrossRef]
- Sanbonmatsu, K.Y. Alignment/misalignment hypothesis for tRNA selection by the ribosome. Biochimie 2006, 88, 1075–1089. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.; Zavialov, A.; Li, W.; Stagg, S.M.; Sengupta, J.; Nielsen, R.C.; Nissen, P.; Harvey, S.C.; Ehrenberg, M.; Frank, J. Incorporation of aminoacyl-tRNA into the ribosome as seen by cryo-electron microscopy. Nat. Struct. Biol. 2003, 10, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Loveland, A.B.; Demo, G.; Grigorieff, N.; Korostelev, A.A. Ensemble cryo-EM elucidates the mechanism of translation fidelity. Nature 2017, 546, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Rozov, A.; Wolff, P.; Grosjean, H.; Yusupov, M.; Yusupova, G.; Westhof, E. Tautomeric G*U pairs within the molecular ribosomal grip and fidelity of decoding in bacteria. Nucleic Acids Res. 2018, 46, 7425–7435. [Google Scholar] [CrossRef] [PubMed]
- Pak, D.; Du, N.; Kim, Y.; Sun, Y.; Burton, Z.F. Rooted tRNAomes and evolution of the genetic code. Transcription 2018, 9, 137–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pak, D.; Kim, Y.; Burton, Z.F. Aminoacyl-tRNA synthetase evolution and sectoring of the genetic code. Transcription 2018, 9, 205–224. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; Whitford, P.C. Capturing Transition States for tRNA Hybrid-State Formation in the Ribosome. J. Phys. Chem. B 2016, 120, 8768–8775. [Google Scholar] [CrossRef]
- Zhou, J.; Lancaster, L.; Donohue, J.P.; Noller, H.F. How the ribosome hands the A-site tRNA to the P site during EF-G-catalyzed translocation. Science 2014, 345, 1188–1191. [Google Scholar] [CrossRef] [Green Version]
- Bowman, J.C.; Hud, N.V.; Williams, L.D. The ribosome challenge to the RNA world. J. Mol. Evol. 2015, 80, 143–161. [Google Scholar] [CrossRef]
- Wallin, G.; Aqvist, J. The transition state for peptide bond formation reveals the ribosome as a water trap. Proc. Natl. Acad. Sci. USA 2010, 107, 1888–1893. [Google Scholar] [CrossRef] [Green Version]
- Sanbonmatsu, K.Y. Flipping through the genetic code: New developments in discrimination between cognate and near-cognate tRNAs and the effect of antibiotics. J. Mol. Biol. 2014, 426, 3197–3200. [Google Scholar] [CrossRef]
- Wohlgemuth, I.; Brenner, S.; Beringer, M.; Rodnina, M.V. Modulation of the rate of peptidyl transfer on the ribosome by the nature of substrates. J. Biol. Chem. 2008, 283, 32229–32235. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M.; Socan, J.; Himo, F.; Aqvist, J. Mechanistic alternatives for peptide bond formation on the ribosome. Nucleic Acids Res. 2018, 46, 5345–5354. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V. The ribosome as a versatile catalyst: Reactions at the peptidyl transferase center. Curr. Opin. Struct. Biol. 2013, 23, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.J.; Trobro, S.; He, S.L.; Aqvist, J.; Green, R. A Role for the 2′ OH of peptidyl-tRNA substrate in peptide release on the ribosome revealed through RF-mediated rescue. Chem. Biol. 2012, 19, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Sund, J.; Ander, M.; Aqvist, J. Principles of stop-codon reading on the ribosome. Nature 2010, 465, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; Whitford, P.C. Steric interactions lead to collective tilting motion in the ribosome during mRNA-tRNA translocation. Nat. Commun. 2016, 7, 10586. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Cui, X.; Beausang, J.F.; Zhang, H.; Farrell, I.; Cooperman, B.S.; Goldman, Y.E. Elongation factor G initiates translocation through a power stroke. Proc. Natl. Acad. Sci. USA 2016, 113, 7515–7520. [Google Scholar] [CrossRef]
- Mace, K.; Giudice, E.; Chat, S.; Gillet, R. The structure of an elongation factor G-ribosome complex captured in the absence of inhibitors. Nucleic Acids Res. 2018, 46, 3211–3217. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Gagnon, M.G.; Bulkley, D.; Steitz, T.A. Conformational changes of elongation factor G on the ribosome during tRNA translocation. Cell 2015, 160, 219–227. [Google Scholar] [CrossRef]
- Liu, Q.; Fredrick, K. Intersubunit Bridges of the Bacterial Ribosome. J. Mol. Biol. 2016, 428, 2146–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Fredrick, K. Contribution of intersubunit bridges to the energy barrier of ribosomal translocation. Nucleic Acids Res. 2013, 41, 565–574. [Google Scholar] [CrossRef]
- Sergiev, P.V.; Lesnyak, D.V.; Kiparisov, S.V.; Burakovsky, D.E.; Leonov, A.A.; Bogdanov, A.A.; Brimacombe, R.; Dontsova, O.A. Function of the ribosomal E-site: A mutagenesis study. Nucleic Acids Res. 2005, 33, 6048–6056. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kowiatek, B.; Opron, K.; Burton, Z.F. Type-II tRNAs and Evolution of Translation Systems and the Genetic Code. Int. J. Mol. Sci 2018, 19, 3275. [Google Scholar] [CrossRef] [PubMed]
- Root-Bernstein, R.; Kim, Y.; Sanjay, A.; Burton, Z.F. tRNA evolution from the proto-tRNA minihelix world. Transcription 2016, 7, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaswamy, U.; Fox, G.E. RNA ligation and the origin of tRNA. Orig. Life Evol. Biosph. 2003, 33, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, M. The origin of the tRNA molecule: Independent data favor a specific model of its evolution. Biochimie 2012, 94, 1464–1466. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, M. A comparison among the models proposed to explain the origin of the tRNA molecule: A synthesis. J. Mol. Evol. 2009, 69, 1–9. [Google Scholar] [CrossRef]
- Widmann, J.; Di Giulio, M.; Yarus, M.; Knight, R. tRNA creation by hairpin duplication. J. Mol. Evol. 2005, 61, 524–530. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, M.; Surman, A.J.; Cooper, G.J.; Suarez-Marina, I.; Hosni, Z.; Lee, M.P.; Cronin, L. Formation of oligopeptides in high yield under simple programmable conditions. Nat. Commun. 2015, 6, 8385. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Schneider, C.; Okamura, H.; Crisp, A.; Amatov, T.; Dejmek, M.; Carell, T. Wet-dry cycles enable the parallel origin of canonical and non-canonical nucleosides by continuous synthesis. Nat. Commun. 2018, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.J.; Gruic-Sovulj, I. Synthetic and editing mechanisms of aminoacyl-tRNA synthetases. Top Curr. Chem. 2014, 344, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Rodin, S.N.; Ohno, S. Two types of aminoacyl-tRNA synthetases could be originally encoded by complementary strands of the same nucleic acid. Orig. Life Evol. Biosph. 1995, 25, 565–589. [Google Scholar] [CrossRef]
- Carter, C.W., Jr. Coding of Class I and II Aminoacyl-tRNA Synthetases. Adv. Exp. Med. Biol. 2017, 966, 103–148. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rodriguez, L.; Erdogan, O.; Jimenez-Rodriguez, M.; Gonzalez-Rivera, K.; Williams, T.; Li, L.; Weinreb, V.; Collier, M.; Chandrasekaran, S.N.; Ambroggio, X.; et al. Functional Class I and II Amino Acid-activating Enzymes Can Be Coded by Opposite Strands of the Same Gene. J. Biol. Chem. 2015, 290, 19710–19725. [Google Scholar] [CrossRef]
- Carter, C.W., Jr.; Li, L.; Weinreb, V.; Collier, M.; Gonzalez-Rivera, K.; Jimenez-Rodriguez, M.; Erdogan, O.; Kuhlman, B.; Ambroggio, X.; Williams, T.; et al. The Rodin-Ohno hypothesis that two enzyme superfamilies descended from one ancestral gene: An unlikely scenario for the origins of translation that will not be dismissed. Biol. Direct. 2014, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, S.N.; Yardimci, G.G.; Erdogan, O.; Roach, J.; Carter, C.W., Jr. Statistical evaluation of the Rodin-Ohno hypothesis: Sense/antisense coding of ancestral class I and II aminoacyl-tRNA synthetases. Mol. Biol. Evol. 2013, 30, 1588–1604. [Google Scholar] [CrossRef]
- Rodin, A.S.; Rodin, S.N.; Carter, C.W., Jr. On primordial sense-antisense coding. J. Mol. Evol. 2009, 69, 555–567. [Google Scholar] [CrossRef]
- Root-Bernstein, R.; Root-Bernstein, M. The ribosome as a missing link in prebiotic evolution II: Ribosomes encode ribosomal proteins that bind to common regions of their own mRNAs and rRNAs. J. Theor. Biol. 2016, 397, 115–127. [Google Scholar] [CrossRef]
- Root-Bernstein, M.; Root-Bernstein, R. The ribosome as a missing link in the evolution of life. J. Theor. Biol. 2015, 367, 130–158. [Google Scholar] [CrossRef]
- Bernhardt, H.S.; Patrick, W.M. Genetic code evolution started with the incorporation of glycine, followed by other small hydrophilic amino acids. J. Mol. Evol. 2014, 78, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, H.S.; Tate, W.P. Evidence from glycine transfer RNA of a frozen accident at the dawn of the genetic code. Biol. Direct. 2008, 3, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Novozhilov, A.S. Origin and Evolution of the Universal Genetic Code. Annu. Rev. Genet. 2017, 51, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Novozhilov, A.S. Origin and evolution of the genetic code: The universal enigma. IUBMB Life 2009, 61, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
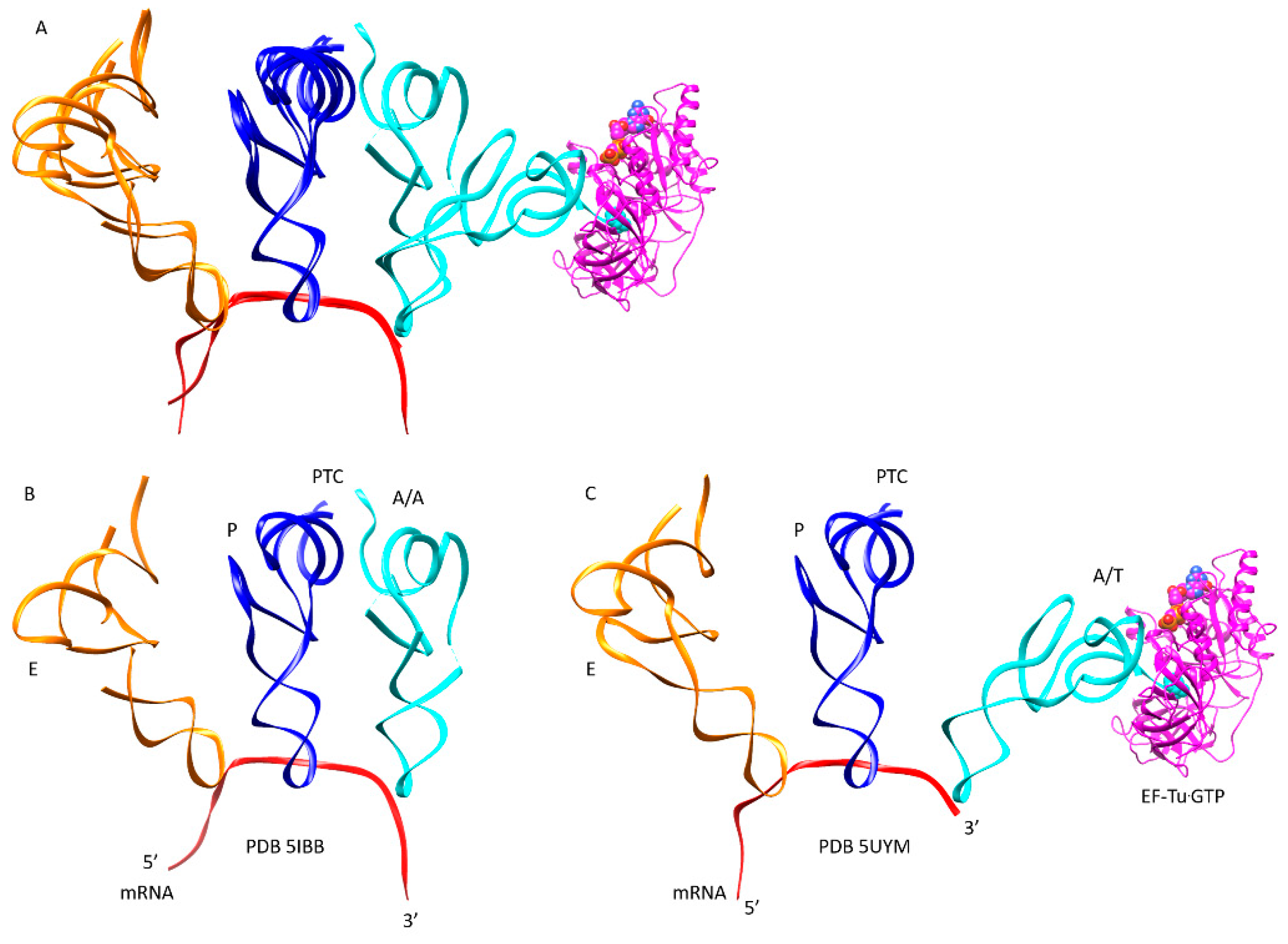
| Intermediate | Stage | Ternary Complex | EF-G | A Site | P Site | E Site | PDB | Proofreading | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (Figure 1) | ASL | CCA/PTC | ASL | CCA/PTC | ASL | CCA | |||||
| A | aa-tRNA.EF-Tu.GTP (free) | empty | empty | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA-peptide | E site | E site U2431, A2432 | 1TTT | |||
| B | Cplx 1 | aa-tRNA.EF-Tu.GTP (bound) | A/T, open | empty | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA-peptide | E site | E site U2431, A2432 | 5UYK | ||
| Cplx 2 | aa-tRNA.EF-Tu.GTP (bound) | A/T, latched | empty | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA-peptide | E site | E site U2431, A2432 | 5UYL | !! | ||
| C | Cplx 3 | aa-tRNA.EF-Tu.GTP (bound) | A/T, latched | empty | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA-peptide | E site | E site U2431, A2432 | 5UYM | !! | |
| D | Elbow | aa-tRNA.EF-Tu.GDP (bound) | EA, latched | empty | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA-peptide | E site | E site U2431, A2432 | !!!! | ||
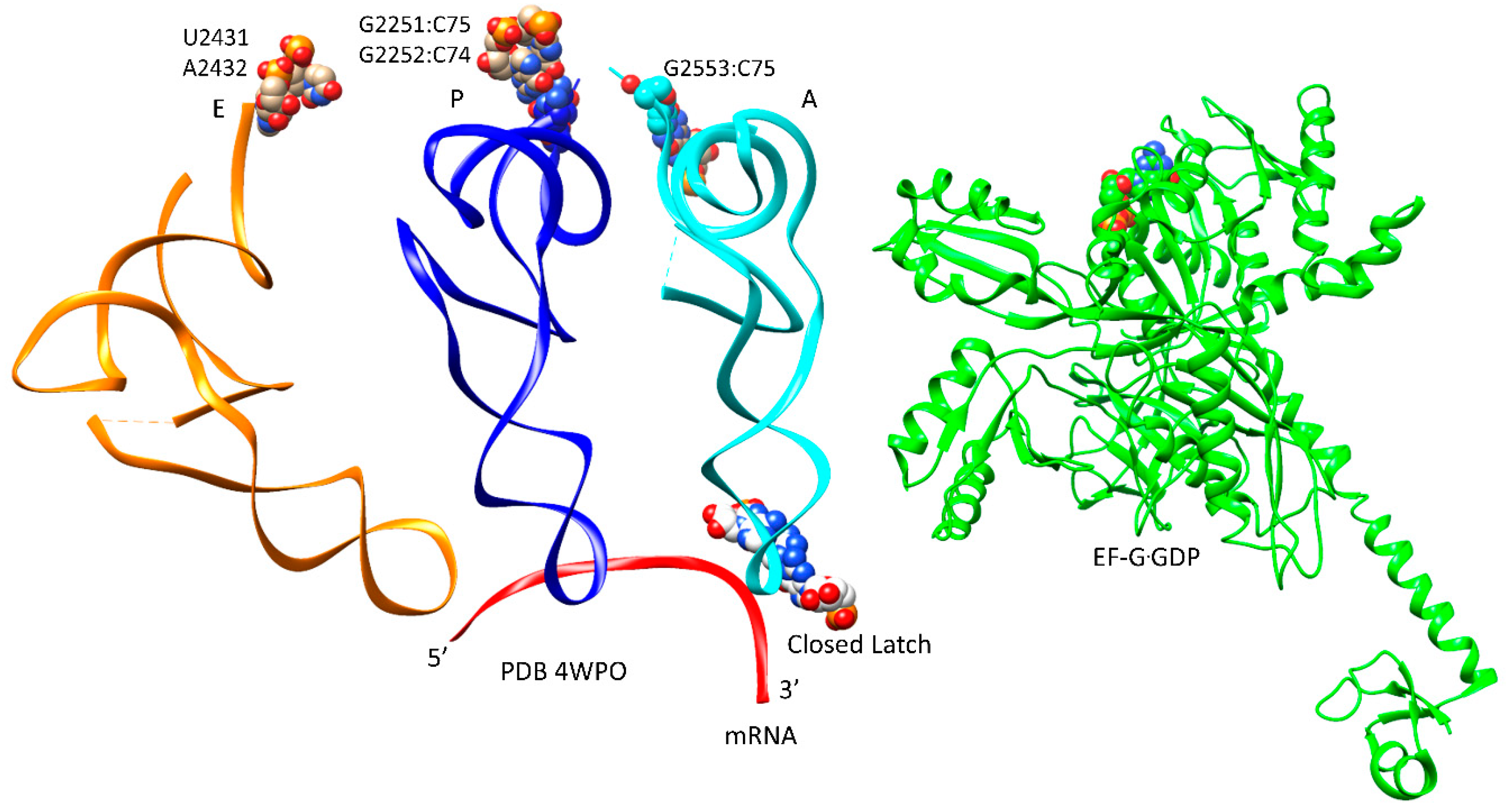
| E | CCA | EF-G.GTP (binds) | A, latched | CCA-aa, C75:G2553 | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA-peptide | E site | E site U2431, A2432 | 5IBB, 4WPO | ||
| F | EF-G.GTP | A, latched | CCA-peptide, C75:G2553 | P site | A2451, C2452, U2585, G2252:C74, G2251:C75, CCA | E site | E site U2431, A2432 | ||||
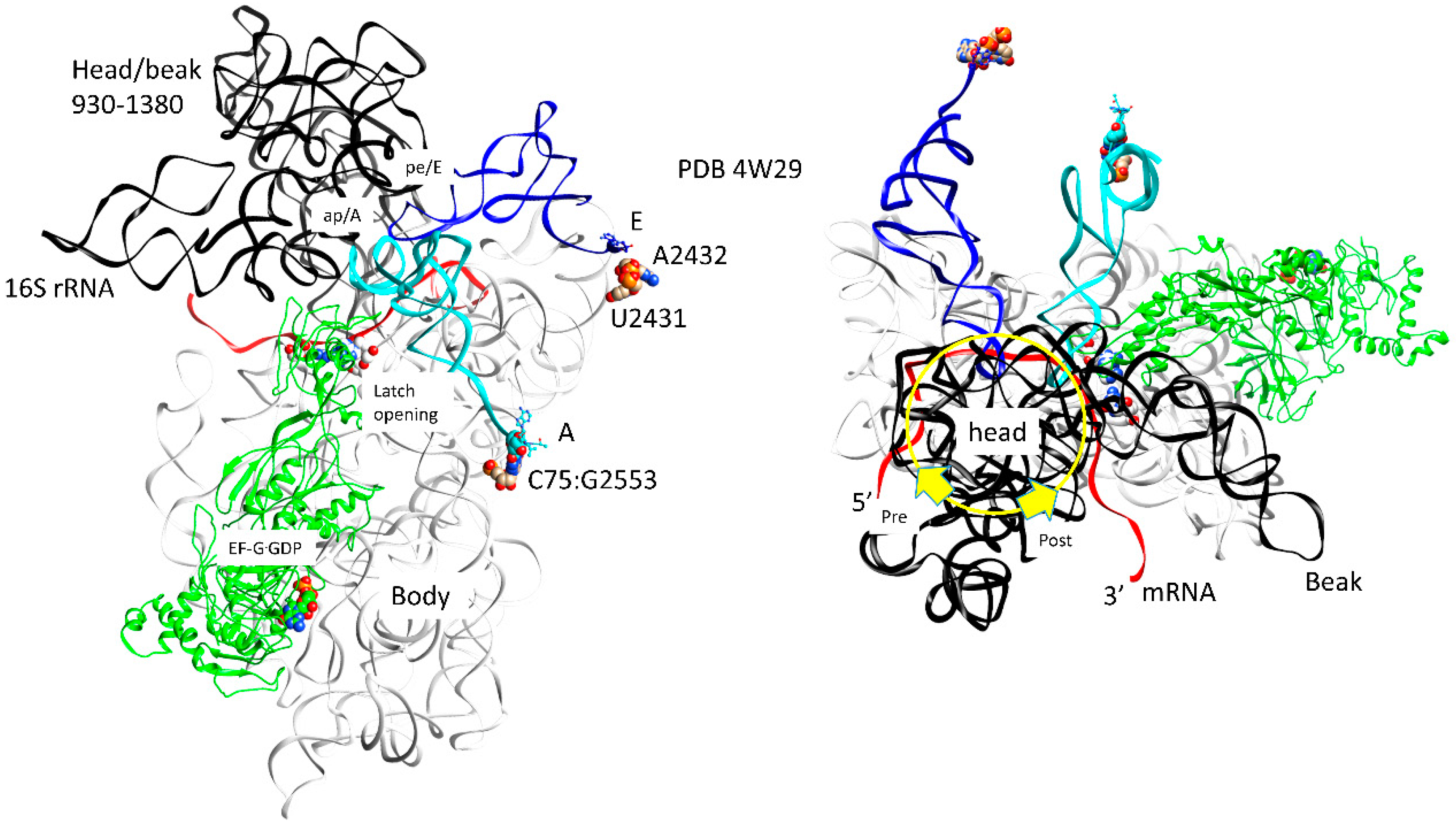
| G | pre | EF-G.GDP | ap, open | (CCA-peptide, C75:G2553) | empty | (ap/A<-->ap/ap tRNA-peptide) | pe | pe/E tRNA: U2431, A2432 | 4W29, 4V5M | ||
| pre/post | EF-G.GDP | ap, open | (CCA-peptide, C75:G2553) | empty | (ap/A<-->ap/ap tRNA-peptide) | pe | pe/E tRNA: U2431, A2432 | ||||
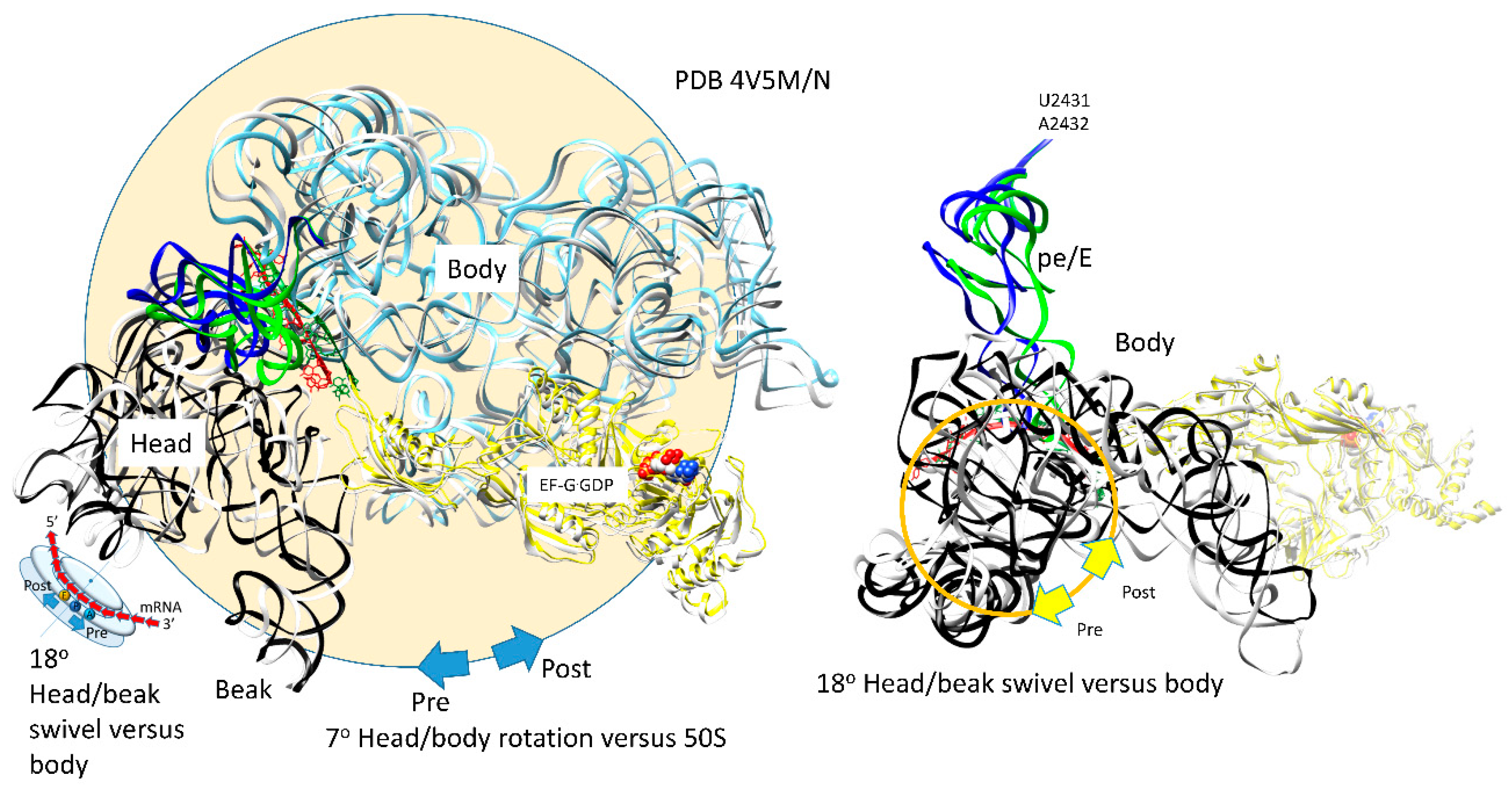
| H | post | EF-G.GDP | ap, open | (CCA-peptide, C75:G2553) | empty | (ap/A<-->ap/ap tRNA-peptide) | pe | pe/E tRNA: U2431, A2432 | 4V5N, 5OT7 | ||
| latch: 30S: S12, 16S: G530~A1492, A1493; 23S: A1913 | |||||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opron, K.; Burton, Z.F. Ribosome Structure, Function, and Early Evolution. Int. J. Mol. Sci. 2019, 20, 40. https://doi.org/10.3390/ijms20010040
Opron K, Burton ZF. Ribosome Structure, Function, and Early Evolution. International Journal of Molecular Sciences. 2019; 20(1):40. https://doi.org/10.3390/ijms20010040
Chicago/Turabian StyleOpron, Kristopher, and Zachary F. Burton. 2019. "Ribosome Structure, Function, and Early Evolution" International Journal of Molecular Sciences 20, no. 1: 40. https://doi.org/10.3390/ijms20010040
APA StyleOpron, K., & Burton, Z. F. (2019). Ribosome Structure, Function, and Early Evolution. International Journal of Molecular Sciences, 20(1), 40. https://doi.org/10.3390/ijms20010040