A PTPmu Biomarker is Associated with Increased Survival in Gliomas


Abstract
:1. Introduction
2. Results
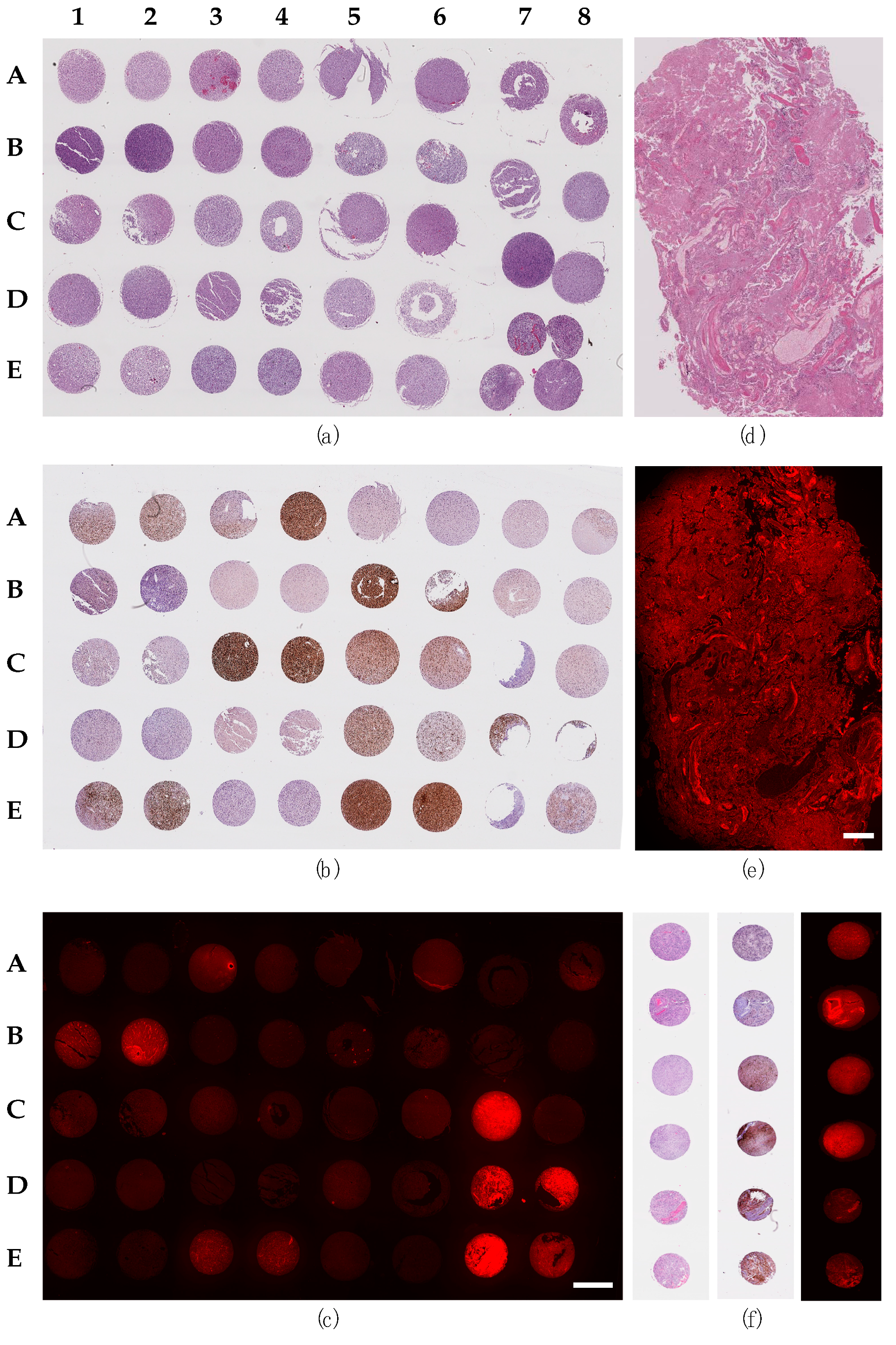
2.1. Staining of Glioma Sections with the PTPµ Biomarker
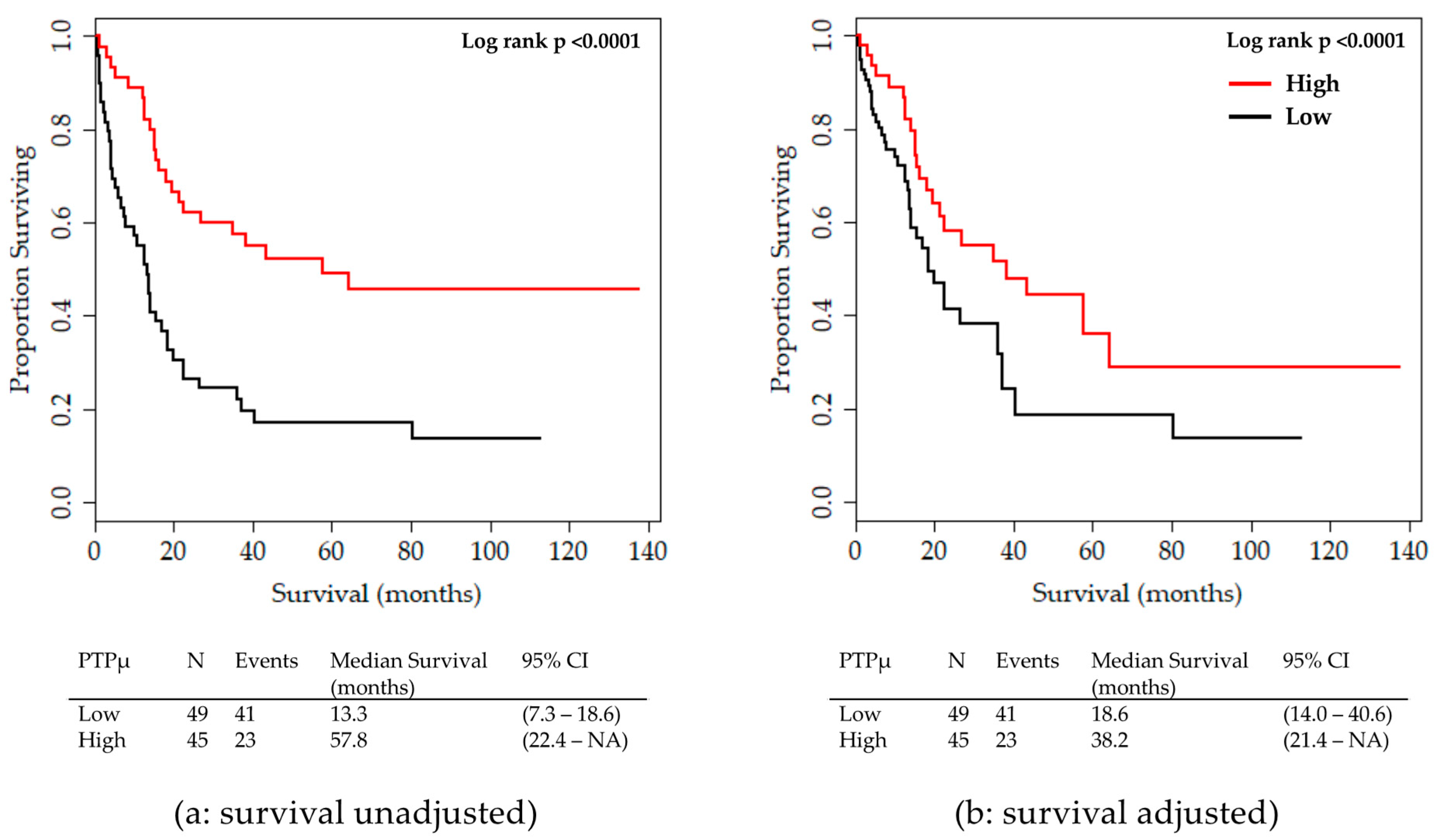
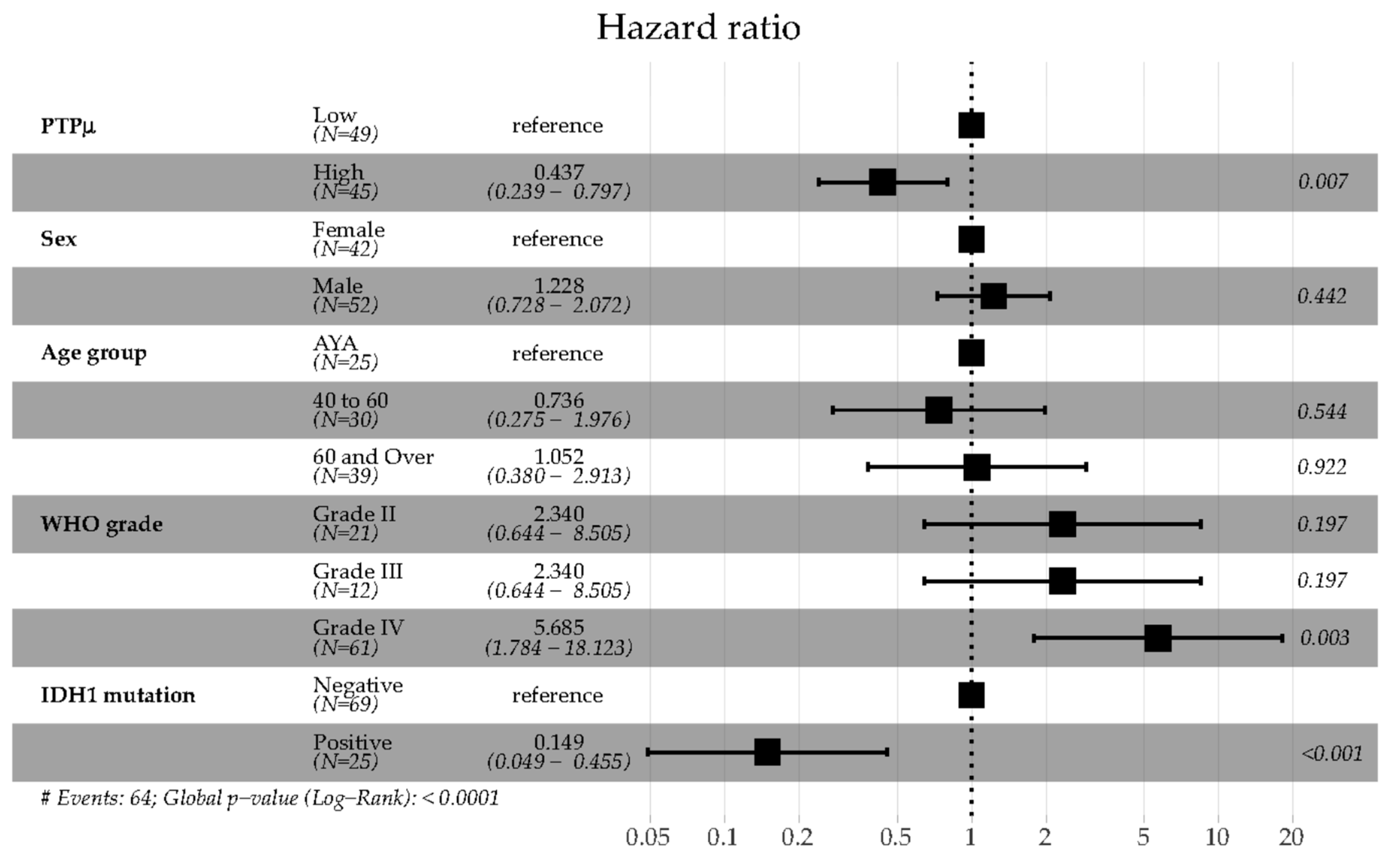
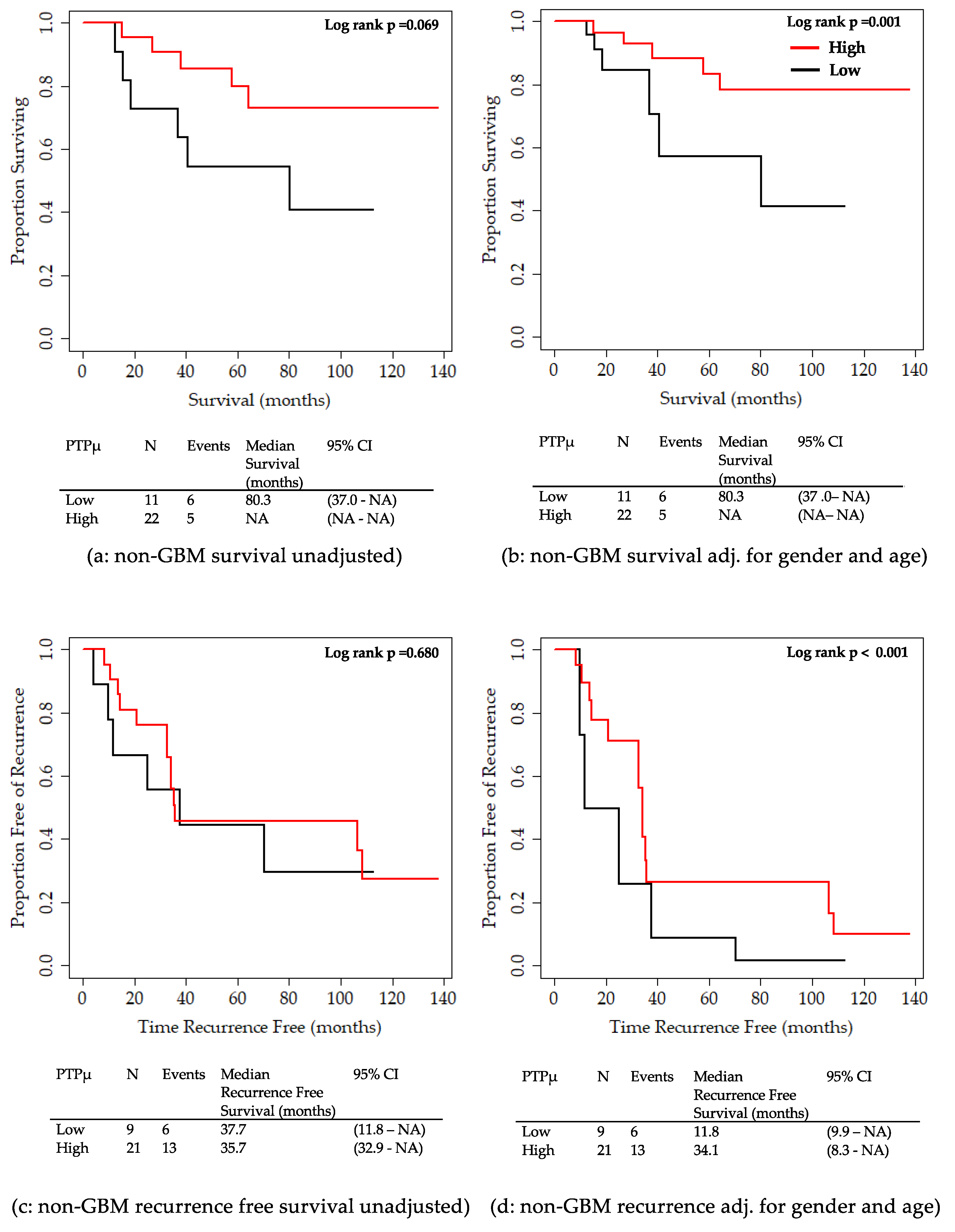
2.2. Analysis of Clinical Variables in Comparison to the PTPµ Biomarker
3. Discussion
4. Materials and Methods
4.1. Study Ethics and Patient Information
4.2. Reagents
4.3. Biomarker Labeling of Human Glioma Tissue
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-ALA | 5-aminolevulinic acid |
| ATRX | Transcriptional regulator ATRX |
| AYA | Adolescent and Young Adult |
| C-GIMP | CpG island Methylator Phenotype |
| CLIA | Clinical Laboratory Improvements Amendments |
| DOTA | 1,4,7,10-Tetraazacyclododecane-1,4,7,10-tetraacetic acid |
| H&E | Hematoxylin and Eosin |
| IDH | Isocitrate Dehydrogenase |
| mAbs | monoclonal Antibodies |
| MRI | Magnetic Resonance Imaging |
| GBM | Glioblastoma |
| RT | Room Temperature |
| TMA | Tissue Microarray |
| TR | Texas Red |
| WHO | World Health Organization |
References
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Vecchione-Koval, T.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and other central nervous system tumors diagnosed in the united states in 2010–2014. Neuro. Oncol. 2017, 19, v1–v88. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.R.; Slone, S.A.; Pittman, T.; St Clair, W.H.; Lightner, D.D.; Villano, J.L. Comprehensive evaluation of treatment and outcomes of low-grade diffuse gliomas. PLoS ONE 2018, 13, e0203639. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Gittleman, H.; Lim, D.; Kattan, M.W.; Chakravarti, A.; Gilbert, M.R.; Lassman, A.B.; Lo, S.S.; Machtay, M.; Sloan, A.E.; Sulman, E.P.; et al. An independently validated nomogram for individualized estimation of survival among patients with newly diagnosed glioblastoma: NRG oncology RTOG 0525 and 0825. Neuro Oncol. 2017, 19, 669–677. [Google Scholar] [PubMed]
- Ostrom, Q.T.; Gittleman, H.; Stetson, L.; Virk, S.M.; Barnholtz-Sloan, J.S. Epidemiology of gliomas. Cancer Treat. Res. 2015, 163, 1–14. [Google Scholar]
- Ostrom, Q.T.; Bauchet, L.; Davis, F.G.; Deltour, I.; Fisher, J.L.; Langer, C.E.; Pekmezci, M.; Schwartzbaum, J.A.; Turner, M.C.; Walsh, K.M.; et al. The epidemiology of glioma in adults: A “state of the science” review. Neuro Oncol. 2014, 16, 896–913. [Google Scholar] [CrossRef]
- Semmel, D.; Ware, C.; Kim, J.Y.; Peters, K.B. Evidence-based treatment for low-grade glioma. Semin. Oncol. Nurs. 2018, 34, 465–471. [Google Scholar] [CrossRef]
- D’Amico, R.S.; Kennedy, B.C.; Bruce, J.N. Neurosurgical oncology: Advances in operative technologies and adjuncts. J. Neurooncol. 2014, 119, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.E.L.; Wright, J.; Sloan, A.E.; Brady-Kalnay, S.M. Fluorescent-guided surgical resection of glioma with targeted molecular imaging agents: A literature review. World Neurosurg. 2016, 90, 154–163. [Google Scholar] [CrossRef]
- Cordova, J.S.; Gurbani, S.S.; Holder, C.A.; Olson, J.J.; Schreibmann, E.; Shi, R.; Guo, Y.; Shu, H.K.; Shim, H.; Hadjipanayis, C.G. Semi-automated volumetric and morphological assessment of glioblastoma resection with fluorescence-guided surgery. Mol. Imaging Biol. 2016, 18, 454–462. [Google Scholar] [CrossRef]
- Stummer, W.; Pichlmeier, U.; Meinel, T.; Wiestler, O.D.; Zanella, F.; Reulen, H.J.; Group, A.L.-G.S. Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: A randomised controlled multicentre phase III trial. Lancet Oncol. 2006, 7, 392–401. [Google Scholar] [CrossRef]
- Colditz, M.J.; Jeffree, R.L. Aminolevulinic acid (ALA)-protoporphyrin IX fluorescence guided tumour resection. Part 1: Clinical, radiological and pathological studies. J. Clin. Neurosci. Off. J. Neurosurg. Soc. Australas. 2012, 19, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Diez Valle, R.; Tejada Solis, S.; Idoate Gastearena, M.A.; Garcia de Eulate, R.; Dominguez Echavarri, P.; Aristu Mendiroz, J. Surgery guided by 5-aminolevulinic fluorescence in glioblastoma: Volumetric analysis of extent of resection in single-center experience. J. Neurooncol. 2011, 102, 105–113. [Google Scholar] [CrossRef]
- Brady-Kalnay, S.M.; Flint, A.J.; Tonks, N.K. Homophilic binding of PTPmu, a receptor-type protein tyrosine phosphatase, can mediate cell-cell aggregation. J. Cell. Biol. 1993, 122, 961–972. [Google Scholar] [CrossRef]
- Burgoyne, A.M.; Phillips-Mason, P.J.; Burden-Gulley, S.M.; Robinson, S.; Sloan, A.E.; Miller, R.H.; Brady-Kalnay, S.M. Proteolytic cleavage of protein tyrosine phosphatase mu regulates glioblastoma cell migration. Cancer Res. 2009, 69, 6960–6968. [Google Scholar] [CrossRef]
- Phillips-Mason, P.J.; Craig, S.E.; Brady-Kalnay, S.M. A protease storm cleaves a cell-cell adhesion molecule in cancer: Multiple proteases converge to regulate PTPmu in glioma cells. J. Cell. Biochem. 2014, 115, 1609–1623. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, A.M.; Palomo, J.M.; Phillips-Mason, P.J.; Burden-Gulley, S.M.; Major, D.L.; Zaremba, A.; Robinson, S.; Sloan, A.E.; Vogelbaum, M.A.; Miller, R.H.; et al. PTPmu suppresses glioma cell migration and dispersal. Neuro Oncol. 2009, 11, 767–778. [Google Scholar] [CrossRef]
- Burden-Gulley, S.M.; Gates, T.J.; Burgoyne, A.M.; Cutter, J.L.; Lodowski, D.T.; Robinson, S.; Sloan, A.E.; Miller, R.H.; Basilion, J.P.; Brady-Kalnay, S.M. A novel molecular diagnostic of glioblastomas: Detection of an extracellular fragment of protein tyrosine phosphatase mu. Neoplasia 2010, 12, 305–316. [Google Scholar] [CrossRef]
- Burden-Gulley, S.M.; Qutaish, M.Q.; Sullivant, K.E.; Tan, M.; Craig, S.E.; Basilion, J.P.; Lu, Z.R.; Wilson, D.L.; Brady-Kalnay, S.M. Single cell molecular recognition of migrating and invading tumor cells using a targeted fluorescent probe to receptor PTPmu. Int. J. Cancer 2013, 132, 1624–1632. [Google Scholar] [CrossRef]
- Herrmann, K.; Johansen, M.L.; Craig, S.E.; Vincent, J.; Howell, M.; Gao, Y.; Lu, L.; Erokwu, B.; Agnes, R.S.; Lu, Z.R.; et al. Molecular imaging of tumors using a quantitative T1 mapping technique via magnetic resonance imaging. Diagn 2015, 5, 318–332. [Google Scholar] [CrossRef]
- Johansen, M.L.; Gao, Y.; Hutnick, M.A.; Craig, S.E.L.; Pokorski, J.K.; Flask, C.A.; Brady-Kalnay, S.M. Quantitative molecular imaging with a single Gd-based contrast agent reveals specific tumor binding and retention in vivo. Anal. Chem. 2017, 89, 5932–5939. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An integrated genomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef]
- Park, S.H.; Won, J.; Kim, S.I.; Lee, Y.; Park, C.K.; Kim, S.K.; Choi, S.H. Molecular testing of brain tumor. J. Pathol. Transl. Med. 2017, 51, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Reuss, D.E.; Sahm, F.; Schrimpf, D.; Wiestler, B.; Capper, D.; Koelsche, C.; Schweizer, L.; Korshunov, A.; Jones, D.T.; Hovestadt, V.; et al. ATRX and Idh1-R132h immunohistochemistry with subsequent copy number analysis and IDH sequencing as a basis for an “integrated” diagnostic approach for adult astrocytoma, oligodendroglioma and glioblastoma. Acta Neuropathol. 2015, 129, 133–146. [Google Scholar] [CrossRef]
- Weller, M.; Weber, R.G.; Willscher, E.; Riehmer, V.; Hentschel, B.; Kreuz, M.; Felsberg, J.; Beyer, U.; Loffler-Wirth, H.; Kaulich, K.; et al. Molecular classification of diffuse cerebral WHO grade II/III gliomas using genome- and transcriptome-wide profiling improves stratification of prognostically distinct patient groups. Acta Neuropathol. 2015, 129, 679–693. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, C.; Hentschel, B.; Wick, W.; Capper, D.; Felsberg, J.; Simon, M.; Westphal, M.; Schackert, G.; Meyermann, R.; Pietsch, T.; et al. Patients with IDH1 wild type anaplastic astrocytomas exhibit worse prognosis than IDH1-mutated glioblastomas, and IDH1 mutation status accounts for the unfavorable prognostic effect of higher age: Implications for classification of gliomas. Acta Neuropathol. 2010, 120, 707–718. [Google Scholar] [CrossRef]
- Craig, S.E.; Brady-Kalnay, S.M. Cancer cells cut homophilic cell adhesion molecules and run. Cancer Res. 2011, 71, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Pogue, B.W.; Gibbs-Strauss, S.; Valdes, P.A.; Samkoe, K.; Roberts, D.W.; Paulsen, K.D. Review of neurosurgical fluorescence imaging methodologies. IEEE J. Sel. Top. Quantum Electron. 2010, 16, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Idoate, M.A.; Diez Valle, R.; Echeveste, J.; Tejada, S. Pathological characterization of the glioblastoma border as shown during surgery using 5-aminolevulinic acid-induced fluorescence. Neuropathology 2011, 31, 575–582. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; de Blank, P.M.; Finlay, J.L.; Gurney, J.G.; McKean-Cowdin, R.; Stearns, D.S.; Wolff, J.E.; Liu, M.; Wolinsky, Y.; et al. American Brain Tumor Association adolescent and young adult primary brain and central nervous system tumors diagnosed in the United States in 2008–2012. Neuro Oncol. 2016, 18, i1–i50. [Google Scholar] [CrossRef] [PubMed]
- Zapotocky, M.; Ramaswamy, V.; Lassaletta, A.; Bouffet, E. Adolescents and young adults with brain tumors in the context of molecular advances in neuro-oncology. Pediatr Blood Cancer 2018, 65. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Suri, V.; Singh, G.; Jha, P.; Purkait, S.; Pathak, P.; Sharma, V.; Sharma, M.C.; Suri, A.; Gupta, D.; et al. Characterization of molecular genetic alterations in GBMs highlights a distinctive molecular profile in young adults. Diagn. Mol. Pathol. 2011, 20, 225–232. [Google Scholar] [CrossRef]
- Noushmehr, H.; Weisenberger, D.J.; Diefes, K.; Phillips, H.S.; Pujara, K.; Berman, B.P.; Pan, F.; Pelloski, C.E.; Sulman, E.P.; Bhat, K.P.; et al. Identification of a CpG island methylator phenotype that defines a distinct subgroup of glioma. Cancer Cell. 2010, 17, 510–522. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Category | PTPµ Low | PTPµ High | p Test |
|---|---|---|---|---|
| Number | 49 | 45 | ||
| Sex (%) | Female | 17 (34.7) | 25 (55.6) | 0.068 1 |
| Male | 32 (65.3) | 20 (44.4) | ||
| Race (%) | Asian | 1 (2.0) | 0 (0.0) | 0.546 1 |
| Black | 2 (4.1) | 1 (2.2) | ||
| White | 46 (93.9) | 44 (97.8) | ||
| Age at Diagnosis [mean (sd)] | 62.2 (14.4) | 46.3 (15.5) | <0.001 2 | |
| Histologic Type (%) | Astrocytoma | 3 (6.1) | 9 (20.0) | 0.044 1 |
| Glioblastoma | 38 (77.6) | 23 (51.1) | ||
| Oligoastrocytoma | 2 (4.1) | 5 (11.1) | ||
| Oligodendroglioma | 6 (12.2) | 8 (17.8) | ||
| WHO Grade (%) | Grade II | 5 (10.2) | 16 (35.6) | 0.010 1 |
| Grade III | 6 (12.2) | 6 (13.3) | ||
| Grade IV | 38 (77.6) | 23 (51.1) | ||
| Recurrence Status (%) | No | 22 (47.8) | 16 (36.4) | 0.375 1 |
| Yes | 24 (52.2) | 28 (63.6) | ||
| Recurrence time in months [mean (sd)] | 12.8 (14.9) | 22.6 (26.4) | 0.114 2 | |
| Survival Status (%) | Alive | 8 (16.3) | 22 (48.9) | 0.002 1 |
| Deceased | 41 (83.7) | 23 (51.1) | ||
| Survival time in months [mean (sd)] | 22.4 (27.9) | 48 (38.1) | <0.001 2 | |
| Age Group (%) | AYA 3 | 6 (12.2) | 19 (42.2) | <0.001 1 |
| 40 to 60 | 12 (24.5) | 18 (40.0) | ||
| 60 and over | 31 (63.3) | 8 (17.8) | ||
| IDH1 mutation (%) | Negative | 41 (83.7) | 28 (62.2) | 0.034 1 |
| Positive | 8 (16.3) | 17 (37.8) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johansen, M.L.; Vincent, J.; Gittleman, H.; Craig, S.E.L.; Couce, M.; Sloan, A.E.; Barnholtz-Sloan, J.S.; Brady-Kalnay, S.M. A PTPmu Biomarker is Associated with Increased Survival in Gliomas. Int. J. Mol. Sci. 2019, 20, 2372. https://doi.org/10.3390/ijms20102372
Johansen ML, Vincent J, Gittleman H, Craig SEL, Couce M, Sloan AE, Barnholtz-Sloan JS, Brady-Kalnay SM. A PTPmu Biomarker is Associated with Increased Survival in Gliomas. International Journal of Molecular Sciences. 2019; 20(10):2372. https://doi.org/10.3390/ijms20102372
Chicago/Turabian StyleJohansen, Mette L., Jason Vincent, Haley Gittleman, Sonya E. L. Craig, Marta Couce, Andrew E. Sloan, Jill S. Barnholtz-Sloan, and Susann M. Brady-Kalnay. 2019. "A PTPmu Biomarker is Associated with Increased Survival in Gliomas" International Journal of Molecular Sciences 20, no. 10: 2372. https://doi.org/10.3390/ijms20102372
APA StyleJohansen, M. L., Vincent, J., Gittleman, H., Craig, S. E. L., Couce, M., Sloan, A. E., Barnholtz-Sloan, J. S., & Brady-Kalnay, S. M. (2019). A PTPmu Biomarker is Associated with Increased Survival in Gliomas. International Journal of Molecular Sciences, 20(10), 2372. https://doi.org/10.3390/ijms20102372