Dysbiosis Disrupts Gut Immune Homeostasis and Promotes Gastric Diseases

 and
and 
Abstract
:1. Introduction
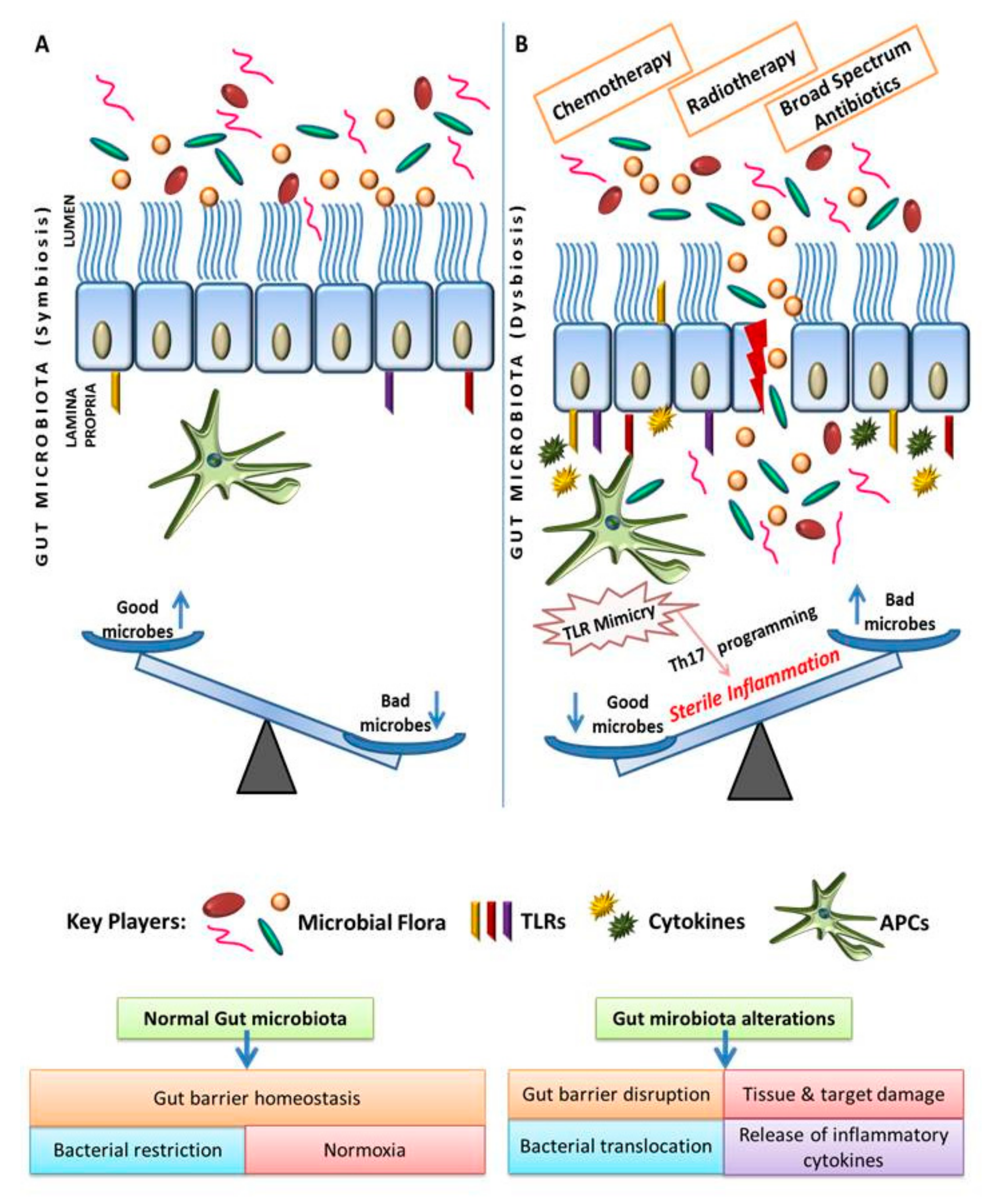
2. Eco-Physiological Balance of Gut Microbiota with Gut-Associated Tissues
3. Gut Microbiota Plays Critical Role in the Maintenance of Mucosal Immune Homeostasis
4. Change in the Gut Microbiome Triggers Sterile Inflammation and Promotes Gastric Inflammatory Disease
5. Change in the Gut Microbiome Triggers TLR Mimicry and Promotes Cancer-Related Inflammation
6. Immune Pharmaceutics as Next-Generation Modalities for Breaking Dysbiosis
7. Role of Bile Acids and Gut Microbiome
Regulation of Bile Acid Synthesis via FXR: Role of Gut Microbial Metabolites
8. Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eloe-Fadrosh, E.A.; Rasko, D.A. The human microbiome: From symbiosis to Pathogenesis. Annu. Rev. Med. 2014, 64, 145–163. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, J.G.; Milani, C.; Giori, G.S.; De Sesma, F.; Van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Ringel, Y.; Sanders, M.E.; Sartor, R.B.; Sherman, P.M.; Versalovic, J.; Young, V.; Finlay, B.B. Perspective Defining a Healthy Human Gut Microbiome: Current Concepts, Future Directions, and Clinical Applications. Cell Host Microbe. 2012, 12, 611–622. [Google Scholar]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Hugon, P.; Dufour, J.C.; Colson, P.; Fournier, P.E.; Sallah, K.; Raoult, D. A comprehensive repertoire of prokaryotic species identified in human beings. Lancet Infect. Dis. 2015, 15, 1211–1219. [Google Scholar] [CrossRef]
- Stanton, C.; Cusack, S.; O’Mahony, D.; O’Connor, K.; Henry, C.; Greene-Diniz, R.; Claesson, M.J.; Fitzgerald, A.P.; Fitzgerald, G.; de Weerd, H.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2010, 108, 4586–4591. [Google Scholar] [Green Version]
- Rodriguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Heal. Dis. 2015, 26, 26050. [Google Scholar] [CrossRef]
- Manichanh, C.; Bork, P.; Hansen, T.; Brunak, S.; Xu, X.; Zhong, H.; Prifti, E.; Chen, W.; Sunagawa, S.; Zhang, W.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar]
- Natividad, J.M.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef]
- Van Eunen, K.; den Besten, G.; Groen, A.K.; Reijngoud, D.; Venema, K.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [Green Version]
- Bäumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S.; Medical, H. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaetzel, C.S.; Frantz, A.L.; Stromberg, A.J.; Rogier, E.W.; Bruno, M.E.C.; Cohen, D.A.; Wedlund, L. Secretory antibodies in breast milk promote long-term intestinal homeostasis by regulating the gut microbiota and host gene expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3074–3079. [Google Scholar] [Green Version]
- Larsbrink, J.; Rogers, T.E.; Hemsworth, G.R.; Mckee, L.S.; Tauzin, A.S.; Spadiut, O.; Klinter, S.; Pudlo, N.A.; Urs, K.; Koropatkin, N.M.; et al. Inhibitors of Apoptosis Protein Antagonists (Smac Mimetic Compounds) Control Polarization of Macrophages during Microbial Challenge and Sterile Inflammatory Responses. Nature 2015, 506, 498–502. [Google Scholar] [CrossRef]
- Goh, Y.J.; Klaenhammer, T.R. Genetic Mechanisms of Prebiotic Oligosaccharide Metabolism in Probiotic Microbes. Annu. Rev. Food Sci. Technol. 2014, 6, 137–156. [Google Scholar] [CrossRef]
- Cani, P.D. Gutmicrobiota and obesity: Lessons from the microbiome. Brief. Funct. Genomics 2013, 12, 381–387. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, 36–44. [Google Scholar] [CrossRef]
- Bohez, L.; Boyen, F.; Timbermont, L.; Ducatelle, R.; Gantois, I.; Pasmans, F.; Haesebrouck, F.; van Immerseel, F. Oral immunisation of laying hens with the live vaccine strains of TAD Salmonella vac® E and TAD Salmonella vac® T reduces internal egg contamination with Salmonella Enteritidis. Vaccine 2006, 24, 6250–6255. [Google Scholar]
- Koutmos, M.; Kabil, O.; Smith, J.L.; Banerjee, R. Structural basis for substrate activation and regulation by cystathionine beta-synthase (CBS) domains in cystathionine –synthase. Proc. Natl. Acad. Sci. USA 2010, 107, 20958–20963. [Google Scholar] [CrossRef]
- Cebra, J.J. Influences of microbiota on intestinal immune system development. Am. J. Clin. Nutr. 1999, 69, 69–1046. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2014, 9, 25. [Google Scholar]
- Malitsky, S.; Rothschild, D.; Moresi, C.; Kuperman, Y.; Elinav, E.; Rozin, S.; Harmelin, A.; Thaiss, C.A.; Halpern, Z.; Levy, M.; et al. Persistent microbiome alterations modulate the rate of post-dieting weight regain. Nature 2016, 540, 544–551. [Google Scholar]
- Magrone, T.; Perez de Heredia, F.; Jirillo, E.; Morabito, G.; Marcos, A.; Serafini, M. Functional foods and nutraceuticals as therapeutic tools for the treatment of diet-related diseases. Can. J. Physiol. Pharmacol. 2013, 396, 387–396. [Google Scholar] [CrossRef]
- Lee, S.M.; Donaldson, G.P.; Mikulski, Z.; Boyajian, S.; Ley, K.; Mazmanian, S.K.; Engineering, B.; Jolla, L. Bacterial colonization factors control specificity and stability of the gut microbiota. Nature 2014, 501, 426–429. [Google Scholar] [CrossRef]
- Marrow, B.; Secreted, S.; Protect, C. Vitamin B12 as a modulator of gut microbial ecology. Cell Metab. 2014, 71, 3831–3840. [Google Scholar]
- Vederas, J.C.; Ross, R.P.; Rea, M.C.; Hill, C.; Sit, C.S.; Zheng, J.; Clayton, E.; O’Connor, P.M.; Whittal, R.M. Thuricin CD, a posttranslationally modified bacteriocin with a narrow spectrum of activity against Clostridium difficile. Proc. Natl. Acad. Sci. USA 2010, 107, 9352–9357. [Google Scholar] [Green Version]
- Rollins, D.; Clements, C.V.; Rodrigues, J.L.M.; Duerkop, B.A.; Hooper, L.V. A composite bacteriophage alters colonization by an intestinal commensal bacterium. Proc. Natl. Acad. Sci. USA 2012, 109, 17621–17626. [Google Scholar] [Green Version]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; Rudensky, A.Y.; Donald, B.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2014, 504, 451–455. [Google Scholar] [CrossRef]
- Da Silva, C.; Wagner, C.; Bonnardel, J.; Gorvel, J.; Lelouard, H. The Peyer’ s Patch Mononuclear Phagocyte System at Steady State and during Infection. Front. Immunol. 2017, 8, 1254. [Google Scholar] [CrossRef]
- Chemouny, J.M.; Gleeson, P.J.; Abbad, L.; Lauriero, G.; Bredel, M.; Bex-coudrat, J.; Boedec, E.; Le Roux, K.; Daugas, E.; Vrtovsnik, F.; et al. Modulation of the microbiota by oral antibiotics treats immunoglobulin A nephropathy in humanized mice. Nephrol. Dial. Transplant. 2018. [Google Scholar] [CrossRef]
- Telesford, K.M.; Yan, W.; Ochoa-reparaz, J.; Pant, A.; Kircher, C.; Christy, M.A.; Begum-haque, S.; Kasper, D.L.; Kasper, L.H.; Telesford, K.M.; et al. A commensal symbiotic factor derived from Bacteroidesfragilis promotes human CD39 C Foxp3 C T cells and T reg function. Gut Microbes 2015, 6, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Kozakova, H.; Schwarzer, M.; Tuckova, L.; Srutkova, D.; Czarnowska, E.; Rosiak, I.; Hudcovic, T.; Schabussova, I.; Hermanova, P.; Zakostelska, Z.; et al. Colonization of germ-free mice with a mixture of three lactobacillus strains enhances the integrity of gut mucosa and ameliorates allergic sensitization. Cell. Mol. Immunol. 2015, 13, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Conlon, M.A.; Bird, A.R. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Arrazuria, R.; Pérez, V.; Molina, E.; Juste, R.A.; Khafipour, E. Diet induced changes in the microbiota and cell composition of rabbit gut associated lymphoid tissue (GALT). Sci. Rep. 2018, 8, 14103. [Google Scholar] [CrossRef] [PubMed]
- Thanabalasuriar, A.; Koutsouris, A.; Hecht, G.; Gruenheid, S. The bacterial virulence factor NleA’s involvement in intestinal tight junction disruption during enteropathogenic E. coli infection is independent of its putative PDZ binding domain. Gut Microbes 2010, 1, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Powell, D.N.; Kalman, D. Layered defense: How mucus and tight junctions seal the intestinal barrier. J. Mol. Med. 2017, 95, 927–934. [Google Scholar] [CrossRef]
- Mohan, M.; Chow, C.T.; Ryan, C.N.; Chan, L.S.; Dufour, J.; Aye, P.P.; Blanchard, J.; Moehs, C.P.; Sestak, K. Dietary Gluten-Induced Gut Dysbiosis Is Accompanied by Selective Upregulation of microRNAs with Intestinal Tight Junction and Bacteria-Binding Motifs in Rhesus Macaque Model of Celiac Disease. Nutrients 2016, 11, 684. [Google Scholar] [CrossRef]
- Hamilton, X.M.K.; Boudry, G.; Lemay, D.G.; Raybould, H.E. Changes in intestinal barrier function and gut microbiota in high-fat diet-fed rats are dynamic and region dependent. Am. J. Physiol. Gastrointest. Liver. Physiol. 2018, 308, G840–G851. [Google Scholar] [CrossRef]
- Inohara, N.; Nun, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal. Immunol. 2017, 10, 18–26. [Google Scholar]
- Salzer, E.; Kansu, A.; Sic, H.; Dogu, F.E.; Prengemann, N.K.; Santos-valente, E.; Pickl, W.F.; Demir, A.M.; Ensari, A.; Colinge, J.; et al. Early-onset inflammatory bowel disease and common variable immunodeficiency—like disease caused by IL-21 deficiency. J. Allergy Clin. Immunol. 2014, 133, 1651–1659. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Stappenbeck, T.S.; Martens, E.C. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Article A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Vatanen, T.; Kostic, A.D.; Hennezel, E.; Cullen, T.W.; Knip, M.; Xavier, R.J.; Huttenhower, C.; Gevers, D.; Cullen, T.W.; Szabo, S.J.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans Article Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Wexler, A.G.; Goodman, A.L. An insider’s perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2017, 2, 1–11. [Google Scholar] [CrossRef]
- Barreau, F.; Meinzer, U.; Chareyre, F.; Berrebi, D.; Niwa-Kawakita, M.; Dussaillant, M.; Foligne, B.; Ollendorff, V.; Heyman, M.; Bonacorsi, S.; et al. CARD15/NOD2 Is Required for Peyer’s Patches Homeostasis in Mice. PLoS ONE 2007, 13, e523. [Google Scholar] [CrossRef] [PubMed]
- Sheflin, A.M.; Whitney, A.K.; Weir, T.L. Cancer-Promoting Effects of Microbial Dysbiosis. Curr. Oncol. Rep. 2014, 16, 406. [Google Scholar] [CrossRef] [PubMed]
- Spurgeon, M.E.; Den Boon, J.A.; Horswill, M.; Barthakur, S.; Forouzan, O.; Rader, J.S. Human papillomavirus oncogenes reprogram the cervical cancer microenvironment independently of and synergistically with estrogen. Proc. Natl. Acad. Sci. USA 2017, 114, E9076–E9085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.L.; Madak-erdogan, Z. Estrogen and Microbiota Crosstalk: Should We Pay Attention? Trends Endocrinol. Metab. 2016, 27, 752–755. [Google Scholar] [CrossRef]
- Bhat, M.I.; Kapila, R. Dietary metabolites derived from gut microbiota: Critical modulators of epigenetic changes in mammals. Nutr. Rev. 2017, 75, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Nadella, V.; Mohanty, A.; Sharma, L.; Yellaboina, S.; Mollenkopf, H.-J.; Mazumdar, V.B.; Palaparthi, R.; Mylavarapu, M.B.; Maurya, R.; Kurukuti, S.; et al. Inhibitors of Apoptosis Protein Antagonists (Smac Mimetic Compounds) Control Polarization of Macrophages during Microbial Challenge and Sterile Inflammatory Responses. Front. Immunol. 2018, 8, 1–21. [Google Scholar]
- Karpiński, T. Role of Oral Microbiota in Cancer Development. Microorganisms 2019, 7, 20. [Google Scholar] [CrossRef]
- Xiong, Y.B.; Zhu, H.R.; Cheng, Y.L.; Yu, Z.H.; Chai, N. Gut microbiome and risk for colorectal cancer. World Chin. J. Dig. 2014, 22, 5653–5658. [Google Scholar] [CrossRef]
- Nagy, K.N.; Sonkodi, I.; Szöke, I.; Nagy, E.; Newman, H.N. The microflora associated with human oral carcinomas. Oral Oncol. 1998, 34, 304–308. [Google Scholar] [CrossRef]
- Gagnaire, A.; Nadel, B.; Raoult, D.; Neefjes, J.; Gorvel, J. Collateral damage: insights into bacterial mechanisms that predispose host cells to cancer. Nat. Rev. Microbiol. 2017, 15, 109. [Google Scholar] [CrossRef] [PubMed]
- Frosali, S.; Pagliari, D.; Gambassi, G.; Landolfi, R.; Pandolfi, F.; Cianci, R. How the Intricate Interaction among Toll-Like Receptors, Microbiota, and Intestinal Immunity Can Influence Gastrointestinal Pathology. J. Immunol. Res. 2015, 2015, 1–12. [Google Scholar] [CrossRef]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut 2018, 67, 1024–1032. [Google Scholar] [CrossRef]
- Xu, T.; Fu, D.; Ren, Y.; Dai, Y.; Lin, J.; Tang, L.; Ji, J. Genetic variations of TLR5 gene interacted with Helicobacter pylori infection among carcinogenesis of gastric cancer. Oncotarget 2017, 8, 31016–31022. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Fang, L.; Lee, M. Dysbiosis of gut microbiota in promoting the development of colorectal cancer. Gastroenterol. Rep. 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, J.; Schlitzer, A.; et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonanantanasarn, K.; Gill, A.L.; Yap, Y.; Jayaprakash, V.; Sullivan, M.A.; Gill, S.R. Enterococcus faecalis Enhances Cell Proliferation through Hydrogen Peroxide-Mediated Epidermal Growth Factor Receptor Activation. Infect. Immun. 2012, 80, 3545–3558. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Qin, H. Microbiota disbiosis is associated with colorectal cancer. Front. Microbiol. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Nougayrède, J.P.; Homburg, S.; Taieb, F.; Boury, M.; Brzuszkiewicz, E.; Gottschalk, G.; Buchrieser, C.; Hacker, J.; Dobrindt, U.; Oswald, E. Escherichia coli induces DNA double-strand breaks in eukaryotic cells. Science 2006, 313, 848–851. [Google Scholar] [CrossRef]
- Boleij, A.; Hechenbleikner, E.M.; Goodwin, A.C.; Badani, R.; Stein, E.M.; Lazarev, M.G.; Ellis, B.; Carroll, K.C.; Albesiano, E.; Wick, E.C.; et al. The bacteroidesfragilis toxin gene is prevalent in the colon mucosa of colorectal cancer patients. Clin. Infect. Dis. 2015, 60, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Biarc, J.; Nguyen, I.S.; Pini, A.; Gossé, F.; Richert, S.; Thiersé, D.; Van Dorsselaer, A.; Leize-Wagner, E.; Raul, F.; Klein, J.P.; et al. Carcinogenic properties of proteins with pro-inflammatory activity from Streptococcusinfantarius (formerly S. bovis). Carcinogenesis 2004, 25, 1477–1484. [Google Scholar] [CrossRef]
- Blot, W.J. Helicobacter pylori protein-specific antibodies and risk of colorectal cancer. Ann. Intern. Med. 2013, 86, 517. [Google Scholar]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum Potentiates Intestinal Tumorigenesis and Modulates the Tumor-Immune Microenvironment. Cell. Host. Microbe. 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, D.; Kim, D.; Cho, J.; Yang, J.; Chung, M.; Kim, K.; Ha, N. Inhibition of proliferation in colon cancer cell lines and harmful enzyme activity of colon bacteria by Bifidobacterium adolescentis SPM0212. Arch. Pharm. Res. 2008, 31, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-time polymerase chain reaction quantification of specific butyrate-producing bacteria, Desulfovibrio and Enterococcus faecalis in the feces of patients with colorectal cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Khan, T.M.; Duncan, S.H.; Harmsen, H.J.M.; Garcia-Gil, L.J.; Flint, H.J. Cultured representatives of two major phylogroups of human colonic Faecalibacteriumprausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth. Appl. Environ. Microbiol. 2012, 78, 420–428. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Wang, L.; Zhou, Y.; Hassan, J.S.; Li, M. Distribution and gene mutation of enteric flora carrying β-glucuronidase among patients with colorectal cancer. Int. J. Clin. Exp. Med. 2015, 8, 5310–5316. [Google Scholar]
- Balzola, F.; Bernstein, C.; Ho, G.T.; Lees, C. Inducible Foxp3+regulatory T-cell development by a commensal bacterium of the intestinal microbiota: Commentary. Inflamm. Bowel Dis. Monit. 2010, 11, 79–80. [Google Scholar]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G.; et al. Faecalibacteriumprausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Delday, M.; Mulder, I.; Logan, E.T.; Grant, G. Bacteroides thetaiotaomicron ameliorates colon inflammation in preclinical models of Crohn’s disease. Inflamm. Bowel Dis. 2019, 25, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Schultz, B.M.; Paduro, C.A.; Salazar, G.A.; Salazar-Echegarai, F.J.; Sebastián, V.P.; Riedel, C.A.; Kalergis, A.M.; Alvarez-Lobos, M.; Bueno, S.M. A potential role of Salmonella infection in the onset of inflammatory bowel diseases. Front. Immunol. 2017, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Boshoff, A.C.; Comprehensive, H.; Boshoff, C. Adherent-invasive E coli in Crohn disease: Bacterial “agent provocateur”. J. Clin. Invest. 2011, 121, 841–844. [Google Scholar]
- Steinert, A.; Linas, I.; Kaya, B.; Ibrahim, M.; Schlitzer, A.; Hruz, P.; Radulovic, K.; Terracciano, L.; Macpherson, A.J.; Niess, J.H. The Stimulation of Macrophages with TLR Ligands Supports Increased IL-19 Expression in Inflammatory Bowel Disease Patients and in Colitis Models. J. Immunol. 2017, 199, 2570–2584. [Google Scholar] [CrossRef]
- Islander, U.; Andersson, A.; Lindberg, E.; Adlerberth, I.; Wold, A.E.; Rudin, A. Superantigenic Staphylococcus aureus stimulates production of interleukin-17 from memory but not naive T cells. Infect. Immun. 2010, 78, 381–386. [Google Scholar] [CrossRef]
- Sook Lee, E.; Ji Song, E.; Do Nam, Y. Dysbiosis of Gut Microbiome and Its Impact on Epigenetic Regulation. J. Clin. Epigenetics 2017, 3, 1–7. [Google Scholar] [CrossRef]
- Neuman, H.; Forsythe, P.; Uzan, A.; Avni, O.; Koren, O.; Medicine, F.; Szold, H.; St, H.; Israel, S.; College, Z.A.; et al. Antibiotics in early life: Dysbiosis and the damage done. FEMS Microbiol. Rev. 2018, 42, 489–499. [Google Scholar] [CrossRef]
- Kristensen, N.B.; Bryrup, T.; Allin, K.H.; Nielsen, T.; Hansen, T.H.; Pedersen, O. Alterations in fecal microbiota composition by probiotic supplementation in healthy adults: A systematic review of randomized controlled trials. Genome Med. 2016, 8, 52. [Google Scholar] [CrossRef]
- Sáez-lara, M.J.; Robles-sanchez, C.; Ruiz-ojeda, F.J.; Plaza-diaz, J.; Gil, A. Effects of Probiotics and Synbiotics on Obesity, Insulin Resistance Syndrome, Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease: A Review of Human Clinical Trials. Int. J. Mol. Sci. 2016, 17, 928. [Google Scholar] [CrossRef]
- Villena, J.; Kitazawa, H. Modulation of intestinal TLR4-inflammatory signaling pathways by probiotic microorganisms: Lessons learned from Lactobacillus jensenii TL2937. Front Immunol. 2014, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Betsi, G.I.; Tokas, T.; Athanasiou, S. Probiotics for Prevention of Recurrent Urinary Tract Infections in Women A Review of the Evidence from Microbiological and Clinical Studies. Drugs 2006, 66, 1253–1261. [Google Scholar] [CrossRef]
- Ooi, L.; Ahmad, R.; Yuen, K.; Liong, M. Lactobacillus acidophilus CHO-220 and inulin reduced plasma total cholesterol and low-density lipoprotein cholesterol via alteration of lipid transporters. J. Dairy Sci. 2010, 93, 5048–5058. [Google Scholar] [CrossRef] [PubMed]
- Brunkwall, L.; Orho-melander, M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: From current human evidence to future possibilities. Diabetologia 2017, 60, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Invest. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.Y.L. Bile Acid Metabolism and Signaling. In Comprehensive Physiology; John Wiley & Sons: Hoboken, NJ, USA, July 2013; pp. 1191–1212. [Google Scholar]
- Wahlström, A.; Sayin, S.I.; Marschall, H.U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota cross-talk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Microorganism | Epigenetic Modifications | Disease |
|---|---|---|
| Enterococcus faecalis | Extracellular superoxide causing DNA breaks | CRC [62] |
| Shigella | Inflammation | CRC [63] |
| Escherichia coli | Syntheses of toxins | CRC [64] |
| Bacteroidesfragilis | Toxin production Inflammatory response by Th17/IL-17 | CRC [65] |
| Streptococcus bovis | Inflammation | CRC [66] |
| Helicobacter pylori | Syntheses of toxins, DNA damage, p53 degradation | CRC [67] |
| Fusobacterium nucleatum | Modulates the tumor immune microenvironment | CRC [68] |
| Bifidobacterium | Decreases b-glucuronidase activity | CRC [69] |
| Eubacterium rectale | Butyrate inducer | CRC [70] |
| Clostridium septicum | Secondary bile acids synthesis | CRC [71] |
| Faecalibacterium prausnitzii | Induces butyrate | CRC [72] |
| Lactobacillus | Decreases lactic acid; activation of Toll-like receptors | CRC [73] |
| Bacteroides fragilis | TLR2 ligand, orchestrates anti-inflammatory immune responses, stimulatesFoxp3C Treg cells | Colitis [74] |
| Faecalibacterium prausnitzii | Inhibits NF-kB activation | Crohn’s disease [75] |
| B. thetaiotaomicron | Attenuates proinflammatory cytokine expression | Colitis [76] |
| Salmonella enteric | Flagellin is recognized by TLR5 which activates proinflammatory pathways in response to infections | Decreased susceptibility to IBD [77] |
| Escherichia coli | NOD2 mutation | Crohn’s disease [78] |
| Staphylococcus aureus | Binds TLR2, inhibits proinflammatory cytokines TNF, IL-12, and IL-6 | IBD [79,80] |
| Eubacterium rectale, Eubacterium hallii, and Roseburia | Natural HDAC inhibitors epigenetically activate p21, bax or suppress Cox-2 | Cancer [81] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toor, D.; Wsson, M.K.; Kumar, P.; Karthikeyan, G.; Kaushik, N.K.; Goel, C.; Singh, S.; Kumar, A.; Prakash, H. Dysbiosis Disrupts Gut Immune Homeostasis and Promotes Gastric Diseases. Int. J. Mol. Sci. 2019, 20, 2432. https://doi.org/10.3390/ijms20102432
Toor D, Wsson MK, Kumar P, Karthikeyan G, Kaushik NK, Goel C, Singh S, Kumar A, Prakash H. Dysbiosis Disrupts Gut Immune Homeostasis and Promotes Gastric Diseases. International Journal of Molecular Sciences. 2019; 20(10):2432. https://doi.org/10.3390/ijms20102432
Chicago/Turabian StyleToor, Devinder, Mishi Kaushal Wsson, Prashant Kumar, G. Karthikeyan, Naveen Kumar Kaushik, Chhavi Goel, Sandhya Singh, Anil Kumar, and Hridayesh Prakash. 2019. "Dysbiosis Disrupts Gut Immune Homeostasis and Promotes Gastric Diseases" International Journal of Molecular Sciences 20, no. 10: 2432. https://doi.org/10.3390/ijms20102432
APA StyleToor, D., Wsson, M. K., Kumar, P., Karthikeyan, G., Kaushik, N. K., Goel, C., Singh, S., Kumar, A., & Prakash, H. (2019). Dysbiosis Disrupts Gut Immune Homeostasis and Promotes Gastric Diseases. International Journal of Molecular Sciences, 20(10), 2432. https://doi.org/10.3390/ijms20102432