Therapeutic Effects of rAAV-Mediated Concomittant Gene Transfer and Overexpression of TGF-β and IGF-I on the Chondrogenesis of Human Bone-Marrow-Derived Mesenchymal Stem Cells

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Successful rAAV-Mediated TGF-β and IGF-I Co-Overexpression in hMSC Aggregates
2.2. Biological and Chondrogenic Differentiation Activities of rAAV-Mediated TGF-β and IGF-I Co-Overexpression in hMSC Aggregates
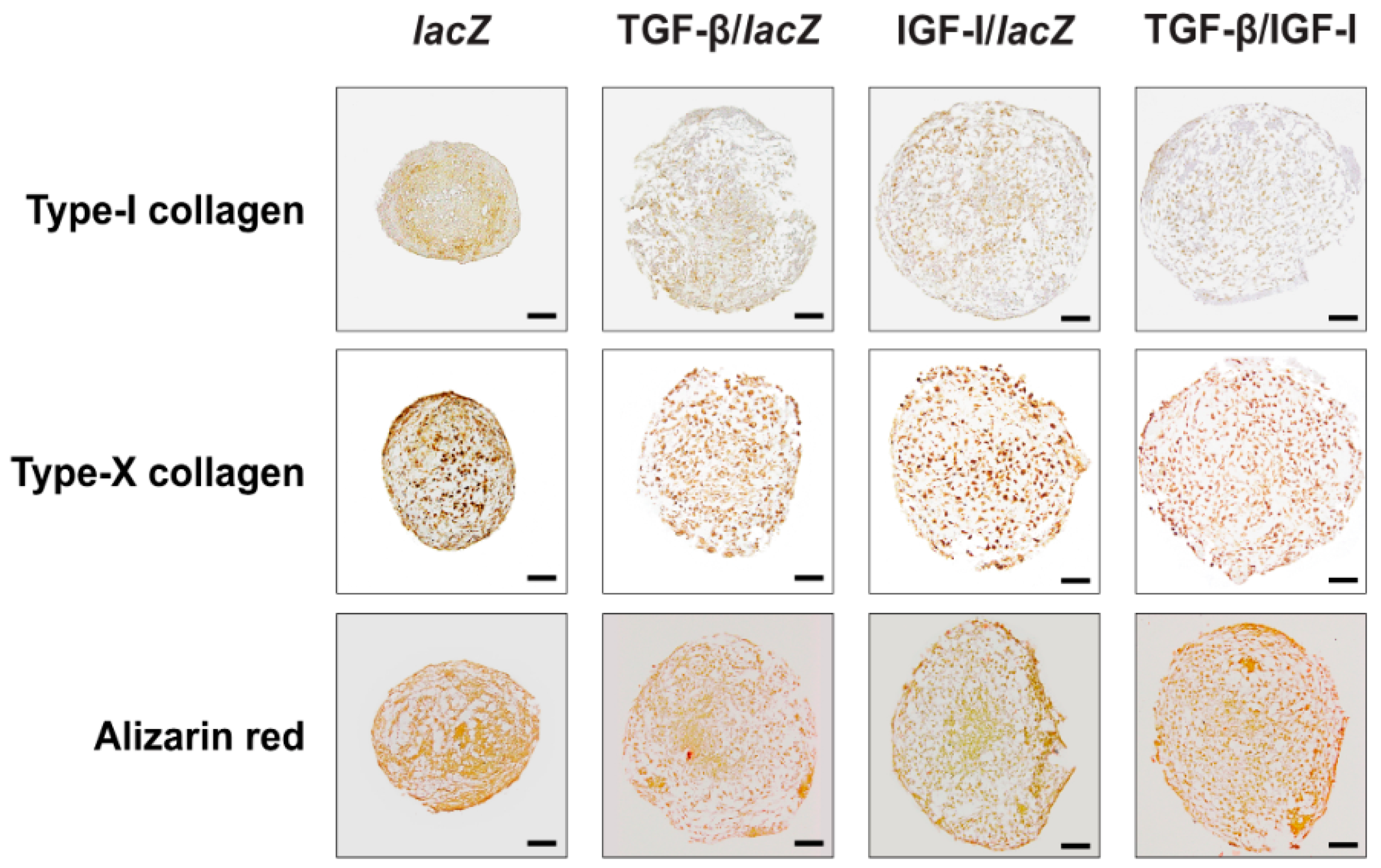
2.3. Effects of rAAV-Mediated TGF-β and IGF-I Co-Overexpression Upon the Osteogenic and Hypertrophic Differentiation Processes in hMSC Aggregates
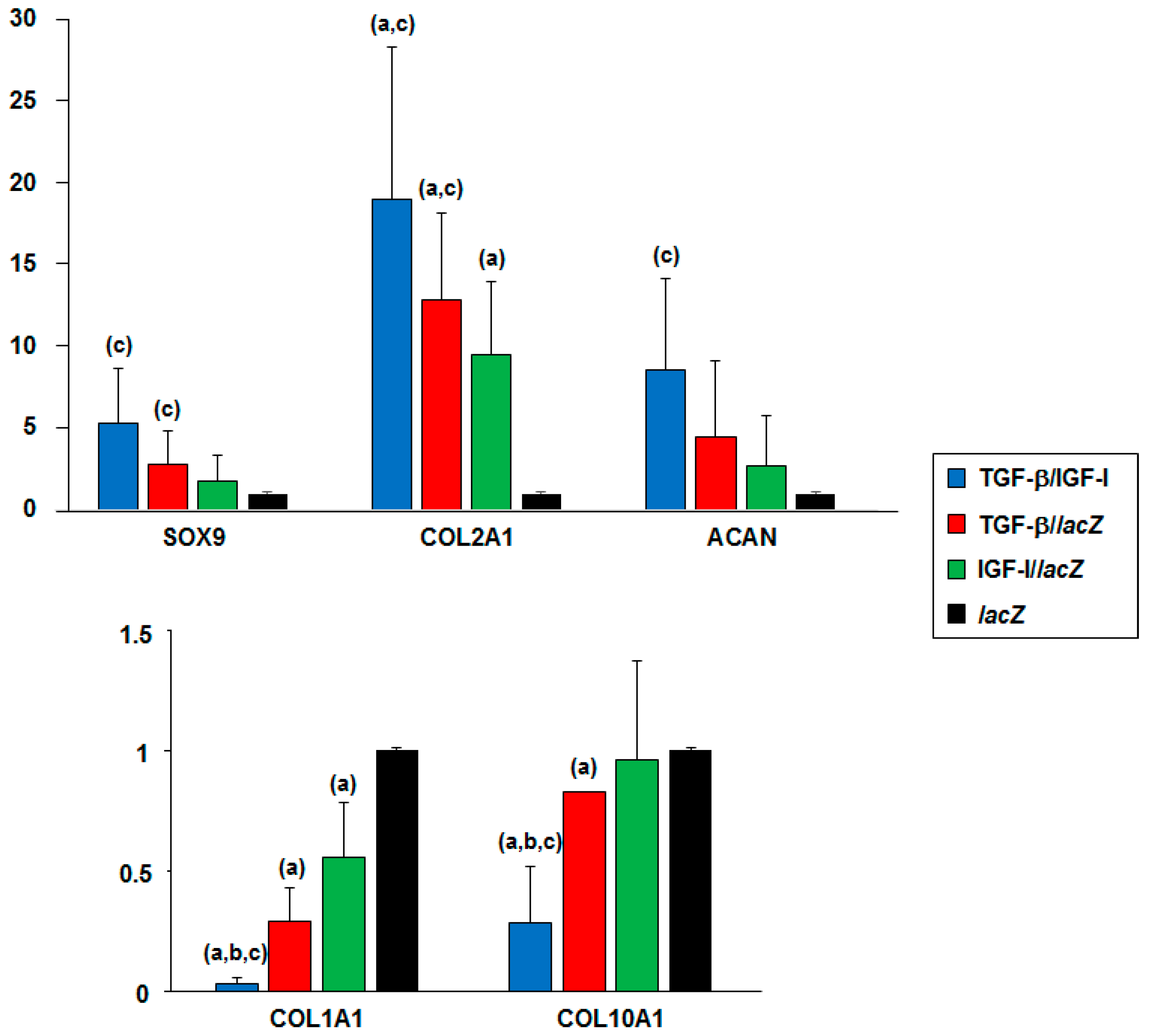
2.4. Real-Time RT-PCR Analyses in hMSC Aggregates Following rAAV-Mediated TGF-β and IGF-I Co-Overexpression
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Reagents
4.3. Cell Culture
4.4. Plasmids and rAAV Vectors
4.5. rAAV-Mediated Gene Transfer
4.6. Transgene Expression
4.7. Histological and Immunohistochemical Analyses
4.8. Histomorphometric Analyses
4.9. Biochemical Analyses
4.10. Real-time RT-PCR Analyses
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buckwalter, J.A. Articular cartilage: Injuries and potential for healing. J. Orthop. Sports Phys. Ther. 1998, 28, 192–202. [Google Scholar] [CrossRef]
- O’Driscoll, S.W. The healing and regeneration of articular cartilage. J. Bone Jt. Surg. Am. 1998, 80, 1795–1812. [Google Scholar] [CrossRef]
- Hunziker, E.B. Articular cartilage repair: Basic science and clinical progress. A review of the current status and prospects. Osteoarthr. Cartil. 2002, 10, 432–463. [Google Scholar] [CrossRef]
- Madry, H.; Grün, U.W.; Knutsen, G. Cartilage repair and joint preservation: Medical and surgical treatment options. Dtsch. Arztebl. Int. 2011, 108, 669–677. [Google Scholar] [PubMed]
- Wakitani, S.; Mitsuoka, T.; Nakamura, N.; Toritsuka, Y.; Nakamura, Y.; Horibe, S. Autologous bone marrow stromal cell transplantation for repair of fullthickness articular cartilage defects in human patellae: Two case reports. Cell Transplant. 2004, 13, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Skowroński, J.; Rutka, M. Osteochondral lesions of the knee reconstructed with mesenchymal stem cells—Results. Ortop. Traumatol. Rehabil. 2013, 15, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Orth, P.; Rey-Rico, A.; Venkatesan, J.K.; Madry, H.; Cucchiarini, M. Current perspectives in stem cell research for knee cartilage repair. Stem Cells Cloning. 2014, 7, 1–17. [Google Scholar] [Green Version]
- Johnstone, B.; Hering, T.M.; Caplan, A.I.; Goldberg, V.M.; Yoo, J.U. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef]
- Mackay, A.M.; Beck, S.C.; Murphy, J.M.; Barry, F.P.; Chichester, C.O.; Pittenger, M.F. Chondrogenic differentiation of cultured human mesenchymal stem cells from marrow. Tissue Eng. 1998, 4, 415–428. [Google Scholar] [CrossRef]
- Barry, F.; Boynton, R.E.; Liu, B.; Murphy, J.M. Chondrogenic differentiation of mesenchymal stem cells from bone marrow: Differentiation-dependent gene expression of matrix components. Exp. Cell. Res. 2001, 268, 189–200. [Google Scholar] [CrossRef]
- Johnstone, B.; Alini, M.; Cucchiarini, M.; Dodge, G.R.; Eglin, D.; Guilak, F.; Madry, H.; Mata, A.; Mauck, R.L.; Semino, C.E.; et al. Tissue engineering for articular cartilage repair—The state of the art. Eur. Cell. Mater. 2013, 25, 247–267. [Google Scholar] [CrossRef]
- Frisch, J.; Venkatesan, J.K.; Rey-Rico, A.; Madry, H.; Cucchiarini, M. Current progress in stem cell-based gene therapy for articular cartilage repair. Curr. Stem Cell Res. Ther. 2015, 10, 121–131. [Google Scholar] [CrossRef]
- Cucchiarini, M. Human gene therapy: Novel approaches to improve the current gene delivery systems. Discov. Med. 2016, 21, 495–506. [Google Scholar]
- Pagnotto, M.R.; Wang, Z.; Karpie, J.C.; Ferretti, M.; Xiao, X.; Chu, C.R. Adeno-associated viral gene transfer of transforming growth factor-beta1 to human mesenchymal stem cells improves cartilage repair. Gene Ther. 2007, 14, 804–813. [Google Scholar] [CrossRef]
- Stender, S.; Murphy, M.; O’Brien, T.; Stengaard, C.; Ulrich-Vinther, M.; Søballe, K.; Barry, F. Adeno-associated viral vector transduction of human mesenchymal stem cells. Eur. Cell Mater. 2007, 13, 93–99. [Google Scholar] [CrossRef]
- Cucchiarini, M.; Ekici, M.; Schetting, S.; Kohn, D.; Madry, H. Metabolic activities and chondrogenic differentiation of human mesenchymal stem cells following recombinant adeno-associated virus-mediated gene transfer and overexpression of fibroblast growth factor 2. Tissue Eng. Part A 2011, 17, 1921–1933. [Google Scholar] [CrossRef]
- Venkatesan, J.K.; Ekici, M.; Madry, H.; Schmitt, G.; Kohn, D.; Cucchiarini, M. SOX9 gene transfer via safe, stable, replication-defective recombinant adeno-associated virus vectors as a novel, powerful tool to enhance the chondrogenic potential of human mesenchymal stem cells. Stem Cell Res. Ther. 2012, 3, 22–36. [Google Scholar] [CrossRef]
- Frisch, J.; Venkatesan, J.K.; Rey-Rico, A.; Schmitt, G.; Madry, H.; Cucchiarini, M. Determination of the chondrogenic differentiation processes in human bone marrow-derived mesenchymal stem cells genetically modified to overexpress transforming growth factor-β via recombinant adeno-associated viral vectors. Hum. Gene Ther. 2014, 25, 1050–1060. [Google Scholar] [CrossRef]
- Frisch, J.; Venkatesan, J.K.; Rey-Rico, A.; Schmitt, G.; Madry, H.; Cucchiarini, M. Influence of insulin-like growth factor I overexpression via recombinant adeno-associated vector gene transfer upon the biological activities and differentiation potential of human bone marrow-derived mesenchymal stem cells. Stem Cell Res. Ther. 2014, 5, 103–114. [Google Scholar] [CrossRef]
- Tao, K.; Frisch, J.; Rey-Rico, A.; Venkatesan, J.K.; Schmitt, G.; Madry, H.; Lin, J.; Cucchiarini, M. Co-overexpression of TGF-β and SOX9 via rAAV gene transfer modulates the metabolic and chondrogenic ativities of human bone marrow-derived mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 20–31. [Google Scholar] [CrossRef]
- Trippel, S.B.; Ghivizzani, S.C.; Nixon, A.J. Gene-based approaches for the repair of articular cartilage. Gene Ther. 2004, 11, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Rey-Rico, A.; Venkatesan, J.K.; Sohier, J.; Moroni, L.; Cucchiarini, M.; Madry, H. Adapted chondrogenic differentiation of human mesenchymal stem cells via controlled release of TGF-β from poly(ethylene oxide)-terephtalate/poly(butylene terephtalate) multiblock scaffolds. J. Biomed. Mater. Res. A 2015, 103, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, F.; Koide, S.; Glimcher, M.J. Cell origin and differentiation in the repair of full-thickness defects of articular cartilage. J. Bone Jt. Surg. Am. 1993, 75, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Mercer, S.; Eckert, G.J.; Trippel, S.B. Growth factor stimulation of growth factors in articular chondrocytes. J. Biol. Chem. 2009, 284, 6697–6704. [Google Scholar] [CrossRef] [PubMed]
- Osborn, K.D.; Trippel, S.B.; Mankin, H.J. Growth factor stimulation of adult articular cartilage. J. Orthop. Res. 1989, 7, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H.; Kitoh, H.; Sugiura, F.; Ishiguro, N. Chondrogenesis enhanced by overexpression of sox9 gene in mouse bone marrow-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2003, 301, 338–343. [Google Scholar] [CrossRef]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2006, 97, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Rey-Rico, A.; Frisch, J.; Venkatesan, J.K.; Schmitt, G.; Rial-Hermida, I.; Taboada, P.; Concheiro, A.; Madry, H.; Alvarez-Lorenzo, C.; Cucchiarini, M. PEO-PPO-PEO carriers for rAAV-mediated transduction of human articular chondrocytes in vitro and in a human osteochondral defect model. ACS Appl. Mater. Interfaces 2016, 8, 20600–20613. [Google Scholar] [CrossRef] [PubMed]
- Cucchiarini, M.; Madry, H. Overexpression of human IGF-I via direct rAAV-mediated gene transfer improves the early repair of articular cartilage defects in vivo. Gene Ther. 2014, 21, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Cucchiarini, M.; Asen, A.K.; Goebel, L.; Venkatesan, J.K.; Schmitt, G.; Zurakowski, D.; Menger, M.D.; Laschke, M.W.; Madry, H. Effects of TGF-β overexpression via rAAV gene transfer on the early repair processes in an osteochondral defect model in minipigs. Am. J. Sports Med. 2018, 46, 1987–1996. [Google Scholar] [CrossRef] [PubMed]
- Samulski, R.J.; Chang, L.S.; Shenk, T. A recombinant plasmid from which an infectious adeno-associated virus genome can be excised in vitro and its use to study viral replication. J. Virol. 1987, 61, 3096–3101. [Google Scholar] [PubMed]
- Samulski, R.J.; Chang, L.S.; Shenk, T. Helper-free stocks of recombinant adeno-associated viruses: Normal integration does not require viral gene expression. J. Virol. 1989, 63, 3822–3828. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | lacZ | TGF-β/lacZ | IGF-I/lacZ | TGF-β/IGF-I | |
|---|---|---|---|---|---|
| TGF-β | day 2 | 162 ± 158 | 196 ± 121 c | 166 ± 127 | 286 ± 169 a,b,c |
| day 7 | 89 ± 93 | 169 ± 119 a,c | 105 ± 93 | 241 ± 183 a,c | |
| day 14 | 19 ± 8 | 80 ± 73 | 22 ± 6 | 121 ± 97 a,c | |
| day 21 | 11 ± 3 | 55 ± 51 a | 15 ± 4 a | 96 ± 80 a,c | |
| IGF-I | day 2 | 52 ± 16 | 59 ± 22 | 76 ± 27 a,b | 93 ± 53 a |
| day 7 | 52 ± 11 | 53 ± 14 | 65 ± 17 a,b | 76 ± 29 a,b | |
| day 14 | 54 ± 13 | 52 ± 15 | 69 ± 15 a,b | 73 ± 17 a,b | |
| day 21 | 53 ± 16 | 49 ± 16 | 67 ± 18 a,b | 71 ± 16 a,b | |
| Assay | lacZ | TGF-β/lacZ | IGF-I/lacZ | TGF-β/IGF-I |
|---|---|---|---|---|
| TGF-β+ cells | 26 ± 6 | 84 ± 6 a,c | 29 ± 12 | 89 ± 4 a,b,c |
| IGF-I+ cells | 27 ± 14 | 33 ± 8 | 82 ± 4 a,b | 90 ± 5 a,b,c |
| Cell densities | 4817 ± 1348 | 9061 ± 840 a,c | 5628 ± 1116 | 14,267 ± 1224 a,b,c |
| Toluidine blue | 2.50 ± 0.58 | 3.75 ± 0.50 a | 3.25 ± 0.50 | 4.00 ± 0.01 a,c |
| Type-II collagen | 1.10 ± 0.32 | 2.50 ± 0.67 a,c | 1.92 ± 0.67 a | 3.20 ± 0.42 a,b,c |
| Type-I collagen | 2.60 ± 0.51 | 1.50 ± 0.52 a | 2.00 ± 0.67 a | 1.30 ± 0.89 a,c |
| Type-X collagen | 3.40 ± 0.67 | 2.20 ± 0.83 a,c | 2.90 ± 0.67 a | 1.30 ± 0.65 a,b,c |
| Alizarin red | 2.90 ± 0.64 | 1.60 ± 0.74 a,c | 2.40 ± 0.52 | 1.10 ± 0.64 a,b,c |
| Assay | lacZ | TGF-β/lacZ | IGF-I/lacZ | TGF-β/IGF-I |
|---|---|---|---|---|
| DNA | 0.029 ± 0.006 | 0.042 ± 0.018 a | 0.034 ± 0.009 a | 0.046 ± 0.010 a,c |
| Proteoglycans | 0.3 ± 0.1 | 0.8 ± 0.7 | 0.6 ± 0.5 | 1.0 ± 0.8 a,b,c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morscheid, S.; Rey-Rico, A.; Schmitt, G.; Madry, H.; Cucchiarini, M.; Venkatesan, J.K. Therapeutic Effects of rAAV-Mediated Concomittant Gene Transfer and Overexpression of TGF-β and IGF-I on the Chondrogenesis of Human Bone-Marrow-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 2591. https://doi.org/10.3390/ijms20102591
Morscheid S, Rey-Rico A, Schmitt G, Madry H, Cucchiarini M, Venkatesan JK. Therapeutic Effects of rAAV-Mediated Concomittant Gene Transfer and Overexpression of TGF-β and IGF-I on the Chondrogenesis of Human Bone-Marrow-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2019; 20(10):2591. https://doi.org/10.3390/ijms20102591
Chicago/Turabian StyleMorscheid, Stephanie, Ana Rey-Rico, Gertrud Schmitt, Henning Madry, Magali Cucchiarini, and Jagadeesh Kumar Venkatesan. 2019. "Therapeutic Effects of rAAV-Mediated Concomittant Gene Transfer and Overexpression of TGF-β and IGF-I on the Chondrogenesis of Human Bone-Marrow-Derived Mesenchymal Stem Cells" International Journal of Molecular Sciences 20, no. 10: 2591. https://doi.org/10.3390/ijms20102591
APA StyleMorscheid, S., Rey-Rico, A., Schmitt, G., Madry, H., Cucchiarini, M., & Venkatesan, J. K. (2019). Therapeutic Effects of rAAV-Mediated Concomittant Gene Transfer and Overexpression of TGF-β and IGF-I on the Chondrogenesis of Human Bone-Marrow-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences, 20(10), 2591. https://doi.org/10.3390/ijms20102591