Synaptic Vesicles Having Large Contact Areas with the Presynaptic Membrane are Preferentially Hemifused at Active Zones of Frog Neuromuscular Junctions Fixed during Synaptic Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
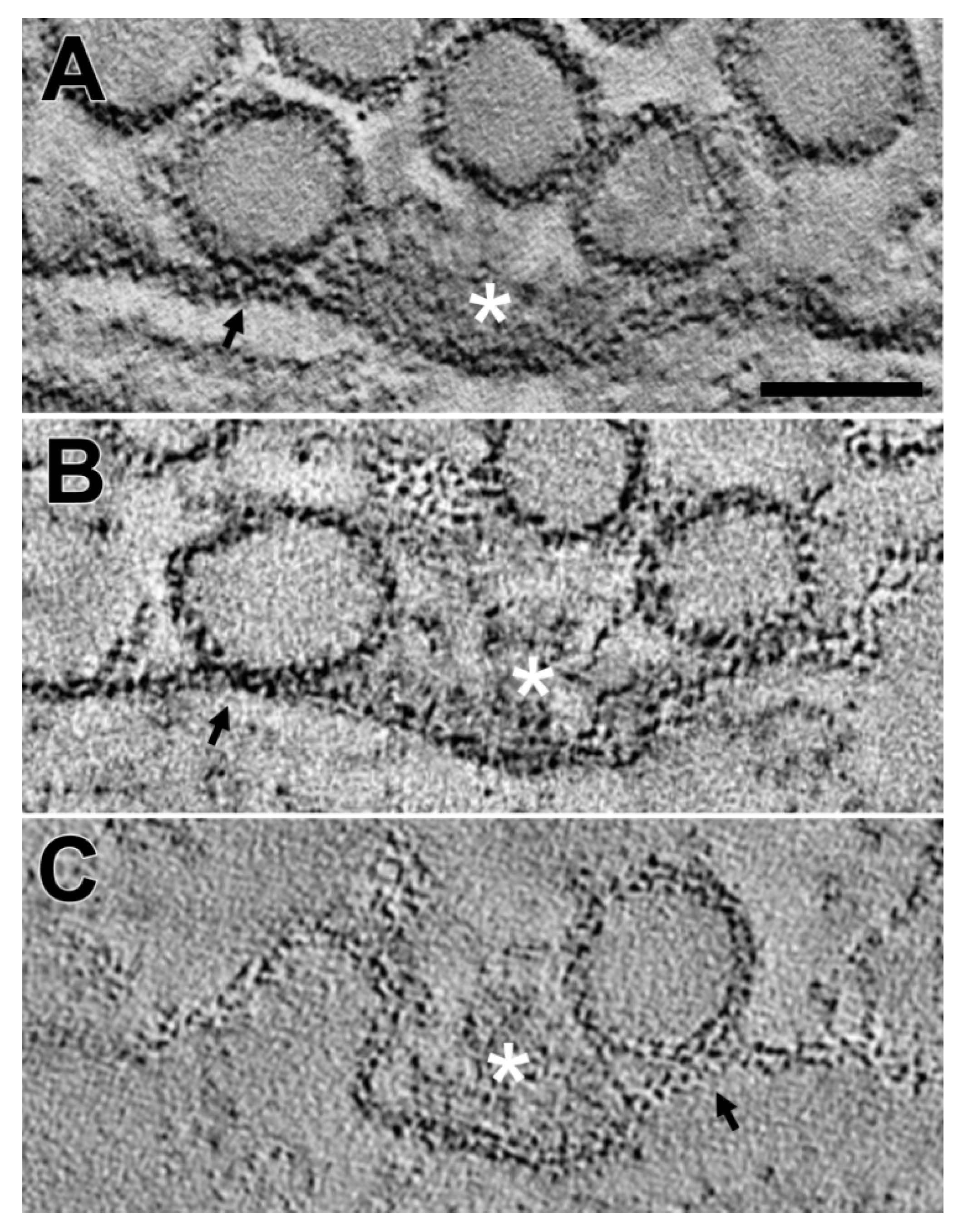
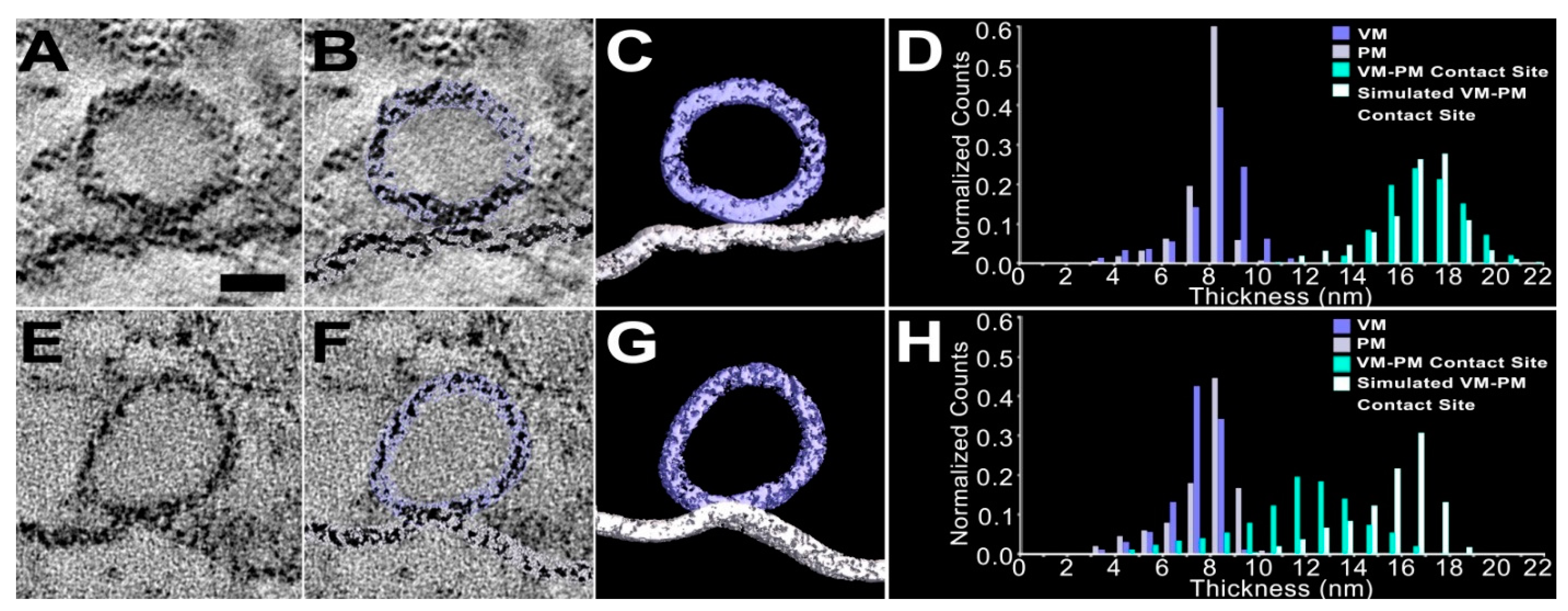
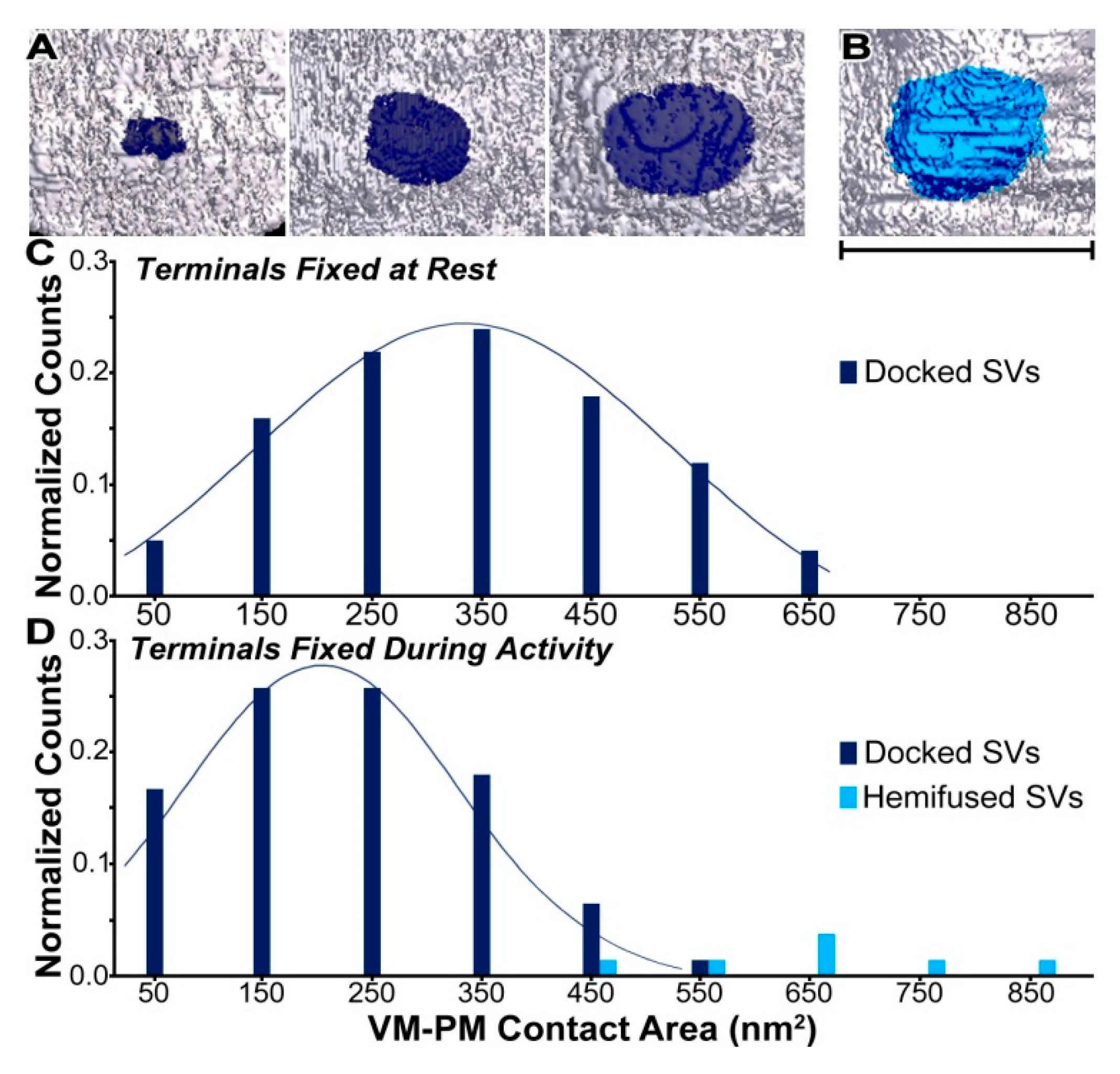
2. Results
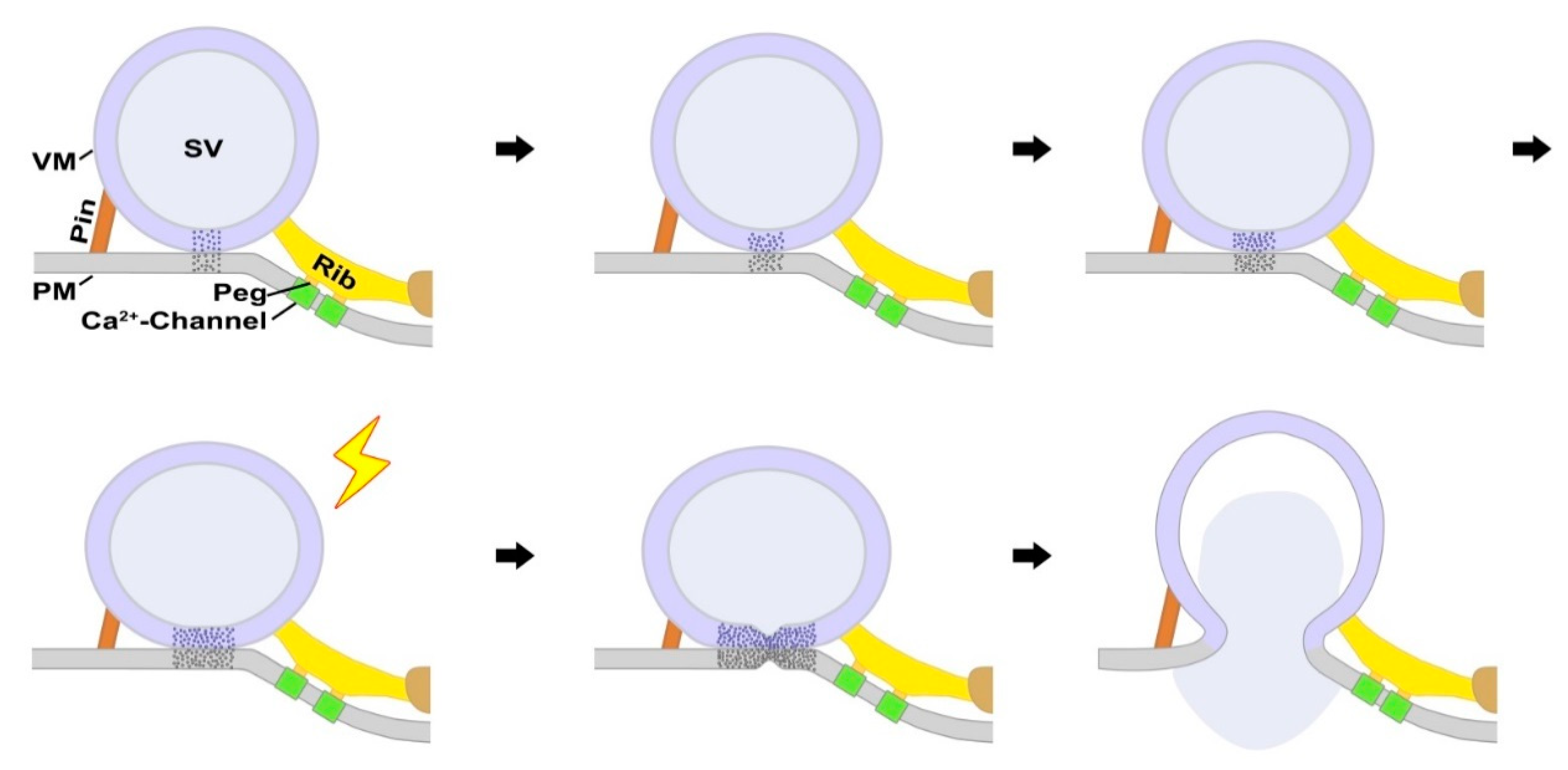
3. Discussion
4. Materials and Methods
4.1. Tissue Preparation
4.2. Sections
4.3. Data Collection
4.4. Reconstruction
4.5. Virtual Slices, Segmentations, and Surface Models
4.6. Measurements
4.6.1. Membrane Thickness
4.6.2. Extent of the Vesicle Membrane–Presynaptic Membrane Contact Area.
4.7. Statistical Analyses
Funding
Acknowledgments
Conflicts of Interest
References
- Couteaux, R.; Pecot-Dechavassine, M. synaptic vesicles and pouches at the level of "active zones" of the neuromuscular junction. C R Acad. Sci. Hebd Seances Acad. Sci. D 1970, 271, 2346–2349. [Google Scholar]
- Peters, A.; Palay, S.L.; Webster, H.D. The Fine Structure of the Nervous System: Neurons and Their Supporting Cells, 3rd ed.; Oxford University Press: New York, NY, USA, 1991; ISBN1 0195065719. ISBN2 978-0195065718. [Google Scholar]
- Heuser, J.E.; Reese, T.S.; Landis, D.M. Functional changes in frog neuromuscular junctions studied with freeze-fracture. J. Neurocytol 1974, 3, 109–131. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.W.; Jones, O.T.; Angelides, K.J. Distribution of Ca2+ channels on frog motor nerve terminals revealed by fluorescent omega-conotoxin. J. Neurosci. 1991, 11, 1032–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumplin, D.W.; Reese, T.S.; Llinas, R. Are the presynaptic membrane particles the calcium channels? Proc. Natl. Acad. Sci. USA 1981, 78, 7210–7213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robitaille, R.; Adler, E.M.; Charlton, M.P. Calcium channels and calcium-gated potassium channels at the frog neuromuscular junction. J. Physiol. Paris 1993, 87, 15–24. [Google Scholar] [CrossRef]
- Heuser, J.E.; Reese, T.S.; Dennis, M.J.; Jan, Y.; Jan, L.; Evans, L. Synaptic vesicle exocytosis captured by quick freezing and correlated with quantal transmitter release. J. Cell Biol. 1979, 81, 275–300. [Google Scholar] [CrossRef]
- Katz, B. The Release of Neural Transmitter Substances; Liverpool University Press: Liverpool, UK, 1969; ISBN1 0853230609. ISBN2 9780853230601. [Google Scholar]
- Brose, N.; Petrenko, A.G.; Sudhof, T.C.; Jahn, R. Synaptotagmin: A calcium sensor on the synaptic vesicle surface. Science 1992, 256, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Geppert, M.; Goda, Y.; Hammer, R.E.; Li, C.; Rosahl, T.W.; Stevens, C.F.; Sudhof, T.C. Synaptotagmin i: A major Ca2+ sensor for transmitter release at a central synapse. Cell 1994, 79, 717–727. [Google Scholar] [CrossRef]
- Hanson, P.I.; Roth, R.; Morisaki, H.; Jahn, R.; Heuser, J.E. Structure and conformational changes in nsf and its membrane receptor complexes visualized by quick-freeze/deep-etch electron microscopy. Cell 1997, 90, 523–535. [Google Scholar] [CrossRef]
- Hata, Y.; Slaughter, C.A.; Sudhof, T.C. Synaptic vesicle fusion complex contains unc-18 homologue bound to syntaxin. Nature 1993, 366, 347–351. [Google Scholar] [CrossRef]
- Schiavo, G.; Benfenati, F.; Poulain, B.; Rossetto, O.; Polverino de Laureto, P.; DasGupta, B.R.; Montecucco, C. Tetanus and botulinum-b neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 1992, 359, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Schoch, S.; Castillo, P.E.; Jo, T.; Mukherjee, K.; Geppert, M.; Wang, Y.; Schmitz, F.; Malenka, R.C.; Sudhof, T.C. Rim1alpha forms a protein scaffold for regulating neurotransmitter release at the active zone. Nature 2002, 415, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Sollner, T.; Whiteheart, S.W.; Brunner, M.; Erdjument-Bromage, H.; Geromanos, S.; Tempst, P.; Rothman, J.E. Snap receptors implicated in vesicle targeting and fusion. Nature 1993, 362, 318–324. [Google Scholar] [CrossRef]
- Sutton, R.B.; Fasshauer, D.; Jahn, R.; Brunger, A.T. Crystal structure of a snare complex involved in synaptic exocytosis at 2.4 a resolution. Nature 1998, 395, 347–353. [Google Scholar] [CrossRef]
- Poirier, M.A.; Xiao, W.; Macosko, J.C.; Chan, C.; Shin, Y.K.; Bennett, M.K. The synaptic snare complex is a parallel four-stranded helical bundle. Nat. Struct. Biol. 1998, 5, 765–769. [Google Scholar] [CrossRef]
- Trimble, W.S.; Cowan, D.M.; Scheller, R.H. Vamp-1: A synaptic vesicle-associated integral membrane protein. Proc. Natl. Acad. Sci. USA 1988, 85, 4538–4542. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Okamoto, M.; Schmitz, F.; Hofmann, K.; Sudhof, T.C. Rim is a putative rab3 effector in regulating synaptic-vesicle fusion. Nature 1997, 388, 593–598. [Google Scholar] [CrossRef]
- Reim, K.; Mansour, M.; Varoqueaux, F.; McMahon, H.T.; Sudhof, T.C.; Brose, N.; Rosenmund, C. Complexins regulate a late step in Ca2+-dependent neurotransmitter release. Cell 2001, 104, 71–81. [Google Scholar] [CrossRef]
- Bennett, M.K.; Calakos, N.; Scheller, R.H. Syntaxin: A synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science 1992, 257, 255–259. [Google Scholar] [CrossRef]
- Maximov, A.; Tang, J.; Yang, X.; Pang, Z.P.; Sudhof, T.C. Complexin controls the force transfer from snare complexes to membranes in fusion. Science 2009, 323, 516–521. [Google Scholar] [CrossRef]
- Südhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with snare and sm proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Abdulreda, M.H.; Bhalla, A.; Chapman, E.R.; Moy, V.T. Atomic force microscope spectroscopy reveals a hemifusion intermediate during soluble n-ethylmaleimide-sensitive factor-attachment protein receptors-mediated membrane fusion. Biophys. J. 2008, 94, 648–655. [Google Scholar] [CrossRef]
- Deak, F.; Xu, Y.; Chang, W.P.; Dulubova, I.; Khvotchev, M.; Liu, X.; Sudhof, T.C.; Rizo, J. Munc18-1 binding to the neuronal snare complex controls synaptic vesicle priming. J. Cell Biol. 2009, 184, 751–764. [Google Scholar] [CrossRef]
- Liu, T.; Wang, T.; Chapman, E.R.; Weisshaar, J.C. Productive hemifusion intermediates in fast vesicle fusion driven by neuronal snares. Biophys. J. 2008, 94, 1303–1314. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Su, L.; Seven, A.B.; Xu, Y.; Rizo, J. Reconstitution of the vital functions of munc18 and munc13 in neurotransmitter release. Science 2013, 339, 421–425. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, F.; Su, Z.; McNew, J.A.; Shin, Y.K. Hemifusion in snare-mediated membrane fusion. Nat. Struct. Mol. Biol. 2005, 12, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Yoon, T.Y.; Okumus, B.; Zhang, F.; Shin, Y.K.; Ha, T. Multiple intermediates in snare-induced membrane fusion. Proc. Natl. Acad. Sci. USA 2006, 103, 19731–19736. [Google Scholar] [CrossRef]
- Hernandez, J.M.; Stein, A.; Behrmann, E.; Riedel, D.; Cypionka, A.; Farsi, Z.; Walla, P.J.; Raunser, S.; Jahn, R. Membrane fusion intermediates via directional and full assembly of the snare complex. Science 2012, 336, 1581–1584. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, F.; McNew, J.A.; Shin, Y.K. Membrane fusion induced by neuronal snares transits through hemifusion. J. Biol. Chem. 2005, 280, 30538–30541. [Google Scholar] [CrossRef]
- Shi, L.; Shen, Q.T.; Kiel, A.; Wang, J.; Wang, H.W.; Melia, T.J.; Rothman, J.E.; Pincet, F. Snare proteins: One to fuse and three to keep the nascent fusion pore open. Science 2012, 335, 1355–1359. [Google Scholar] [CrossRef]
- Chernomordik, L.V.; Kozlov, M.M. Mechanics of membrane fusion. Nat. Struct. Mol. Biol. 2008, 15, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zampighi, G.A.; Fain, N.; Zampighi, L.M.; Cantele, F.; Lanzavecchia, S.; Wright, E.M. Conical electron tomography of a chemical synapse: Polyhedral cages dock vesicles to the active zone. J. Neurosci. 2008, 28, 4151–4160. [Google Scholar] [CrossRef]
- Zampighi, G.A.; Zampighi, L.M.; Fain, N.; Lanzavecchia, S.; Simon, S.A.; Wright, E.M. Conical electron tomography of a chemical synapse: Vesicles docked to the active zone are hemi-fused. Biophys. J. 2006, 91, 2910–2918. [Google Scholar] [CrossRef]
- Almers, W.; Tse, F.W. Transmitter release from synapses: Does a preassembled fusion pore initiate exocytosis? Neuron 1990, 4, 813–818. [Google Scholar] [CrossRef]
- Jackson, M.B. Snare complex zipping as a driving force in the dilation of proteinaceous fusion pores. J. Membr. Biol. 2010, 235, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.B.; Chapman, E.R. The fusion pores of Ca2+ -triggered exocytosis. Nat. Struct. Mol. Biol. 2008, 15, 684–689. [Google Scholar] [CrossRef]
- Gundersen, C.B. The structure of the synaptic vesicle-plasma membrane interface constrains snare models of rapid, synchronous exocytosis at nerve terminals. Front. Mol. Neurosci. 2017, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Wang, C.T.; Bai, J.; Chapman, E.R.; Jackson, M.B. Transmembrane segments of syntaxin line the fusion pore of Ca2+-triggered exocytosis. Science 2004, 304, 289–292. [Google Scholar] [CrossRef]
- Fernandez-Busnadiego, R.; Asano, S.; Oprisoreanu, A.M.; Sakata, E.; Doengi, M.; Kochovski, Z.; Zurner, M.; Stein, V.; Schoch, S.; Baumeister, W.; et al. Cryo-electron tomography reveals a critical role of rim1alpha in synaptic vesicle tethering. J. Cell Biol. 2013, 201, 725–740. [Google Scholar] [CrossRef]
- Fernandez-Busnadiego, R.; Zuber, B.; Maurer, U.E.; Cyrklaff, M.; Baumeister, W.; Lucic, V. Quantitative analysis of the native presynaptic cytomatrix by cryoelectron tomography. J. Cell Biol. 2010, 188, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Imig, C.; Min, S.W.; Krinner, S.; Arancillo, M.; Rosenmund, C.; Sudhof, T.C.; Rhee, J.; Brose, N.; Cooper, B.H. The morphological and molecular nature of synaptic vesicle priming at presynaptic active zones. Neuron 2014, 84, 416–431. [Google Scholar] [CrossRef]
- Siksou, L.; Varoqueaux, F.; Pascual, O.; Triller, A.; Brose, N.; Marty, S. A common molecular basis for membrane docking and functional priming of synaptic vesicles. Eur. J. Neurosci. 2009, 30, 49–56. [Google Scholar] [CrossRef]
- Jung, J.H.; Szule, J.A.; Marshall, R.M.; McMahan, U.J. Variable priming of a docked synaptic vesicle. Proc. Natl. Acad. Sci. USA 2016, 113, E1098–E1107. [Google Scholar] [CrossRef] [PubMed]
- Siksou, L.; Triller, A.; Marty, S. Ultrastructural organization of presynaptic terminals. Curr. Opin. Neurobiol. 2011, 21, 261–268. [Google Scholar] [CrossRef]
- Szule, J.A.; Harlow, M.L.; Jung, J.H.; De-Miguel, F.F.; Marshall, R.M.; McMahan, U.J. Regulation of synaptic vesicle docking by different classes of macromolecules in active zone material. PLoS One 2012, 7, e33333. [Google Scholar] [CrossRef] [PubMed]
- Ress, D.B.; Harlow, M.L.; Marshall, R.M.; McMahan, U.J. Methods for generating high-resolution structural models from electron microscope tomography data. Structure 2004, 12, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Harlow, M.L.; Ress, D.; Stoschek, A.; Marshall, R.M.; McMahan, U.J. The architecture of active zone material at the frog’s neuromuscular junction. Nature 2001, 409, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Harlow, M.L.; Szule, J.A.; Xu, J.; Jung, J.H.; Marshall, R.M.; McMahan, U.J. Alignment of synaptic vesicle macromolecules with the macromolecules in active zone material that direct vesicle docking. PLoS ONE 2013, 8, e69410. [Google Scholar] [CrossRef]
- Nagwaney, S.; Harlow, M.L.; Jung, J.H.; Szule, J.A.; Ress, D.; Xu, J.; Marshall, R.M.; McMahan, U.J. Macromolecular connections of active zone material to docked synaptic vesicles and presynaptic membrane at neuromuscular junctions of mouse. J. Comp. Neurol. 2009, 513, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.H.; Szule, J. Automatic optimization method for segmentation and surface model generation in electron tomography. IEEE Life Sci. Lett. 2017, 3, 5–8. [Google Scholar] [CrossRef]
- Robertson, J.D. The ultrastructure of cell membranes and their derivatives. Biochem Soc. Symp. 1959, 16, 3–43. [Google Scholar]
- Yamamoto, T. On the thickness of the unit membrane. J. Cell Biol. 1963, 17, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Chernomordik, L.; Chanturiya, A.; Green, J.; Zimmerberg, J. The hemifusion intermediate and its conversion to complete fusion: Regulation by membrane composition. Biophys. J. 1995, 69, 922–929. [Google Scholar] [CrossRef]
- Cohen, F.S.; Zimmerberg, J.; Finkelstein, A. Fusion of phospholipid vesicles with planar phospholipid bilayer membranes. Ii. Incorporation of a vesicular membrane marker into the planar membrane. J. Gen. Physiol. 1980, 75, 251–270. [Google Scholar] [CrossRef]
- Lee, J.; Lentz, B.R. Evolution of lipidic structures during model membrane fusion and the relation of this process to cell membrane fusion. Biochemistry 1997, 36, 6251–6259. [Google Scholar] [CrossRef]
- Schaub, J.R.; Lu, X.; Doneske, B.; Shin, Y.K.; McNew, J.A. Hemifusion arrest by complexin is relieved by Ca2+-synaptotagmin i. Nat. Struct. Mol. Biol. 2006, 13, 748–750. [Google Scholar] [CrossRef]
- Wong, J.L.; Koppel, D.E.; Cowan, A.E.; Wessel, G.M. Membrane hemifusion is a stable intermediate of exocytosis. Dev. Cell 2007, 12, 653–659. [Google Scholar] [CrossRef]
- Rosenmund, C.; Stevens, C.F. Definition of the readily releasable pool of vesicles at hippocampal synapses. Neuron 1996, 16, 1197–1207. [Google Scholar] [CrossRef]
- Cole, A.A.; Chen, X.; Reese, T.S. A network of three types of filaments organizes synaptic vesicles for storage, mobilization, and docking. J. Neurosci. 2016, 36, 3222–3230. [Google Scholar] [CrossRef]
- Zampighi, G.A.; Serrano, R.; Vergara, J.L. A novel synaptic vesicle fusion path in the rat cerebral cortex: The "saddle" point hypothesis. PLoS ONE 2014, 9, e100710. [Google Scholar] [CrossRef]
- Jung, J.H.; Doniach, S. A stochastic model of active zone material mediated synaptic vesicle docking and priming at resting active zones. Sci. Rep. 2017, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Szule, J.A.; Stouder, K.; Marshall, R.M.; McMahan, U.J. Active zone material-directed orientation, docking, and fusion of dense core vesicles alongside synaptic vesicles at neuromuscular junctions. Front. Neuroanat. 2018, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Katz, B.; Miledi, R. Estimates of quantal content during ’chemical potentiation’ of transmitter release. Proc. R. Soc. Lond. B Biol. Sci. 1979, 205, 369–378. [Google Scholar] [PubMed]
- Efron, B.; Tibshirani, R. An Introduction to the Bootstrap; Chapman & Hall: New York, NY, USA, 1993; ISBN1 0412042312. ISBN2 978-0412042317. [Google Scholar]




© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, J.H. Synaptic Vesicles Having Large Contact Areas with the Presynaptic Membrane are Preferentially Hemifused at Active Zones of Frog Neuromuscular Junctions Fixed during Synaptic Activity. Int. J. Mol. Sci. 2019, 20, 2692. https://doi.org/10.3390/ijms20112692
Jung JH. Synaptic Vesicles Having Large Contact Areas with the Presynaptic Membrane are Preferentially Hemifused at Active Zones of Frog Neuromuscular Junctions Fixed during Synaptic Activity. International Journal of Molecular Sciences. 2019; 20(11):2692. https://doi.org/10.3390/ijms20112692
Chicago/Turabian StyleJung, Jae Hoon. 2019. "Synaptic Vesicles Having Large Contact Areas with the Presynaptic Membrane are Preferentially Hemifused at Active Zones of Frog Neuromuscular Junctions Fixed during Synaptic Activity" International Journal of Molecular Sciences 20, no. 11: 2692. https://doi.org/10.3390/ijms20112692
APA StyleJung, J. H. (2019). Synaptic Vesicles Having Large Contact Areas with the Presynaptic Membrane are Preferentially Hemifused at Active Zones of Frog Neuromuscular Junctions Fixed during Synaptic Activity. International Journal of Molecular Sciences, 20(11), 2692. https://doi.org/10.3390/ijms20112692