Development of Molecular Markers for Detection of Acidovorax citrulli Strains Causing Bacterial Fruit Blotch Disease in Melon

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Pathogenicity of Acidovorax citrulli Strains NIHHS15-280 and KACC18782
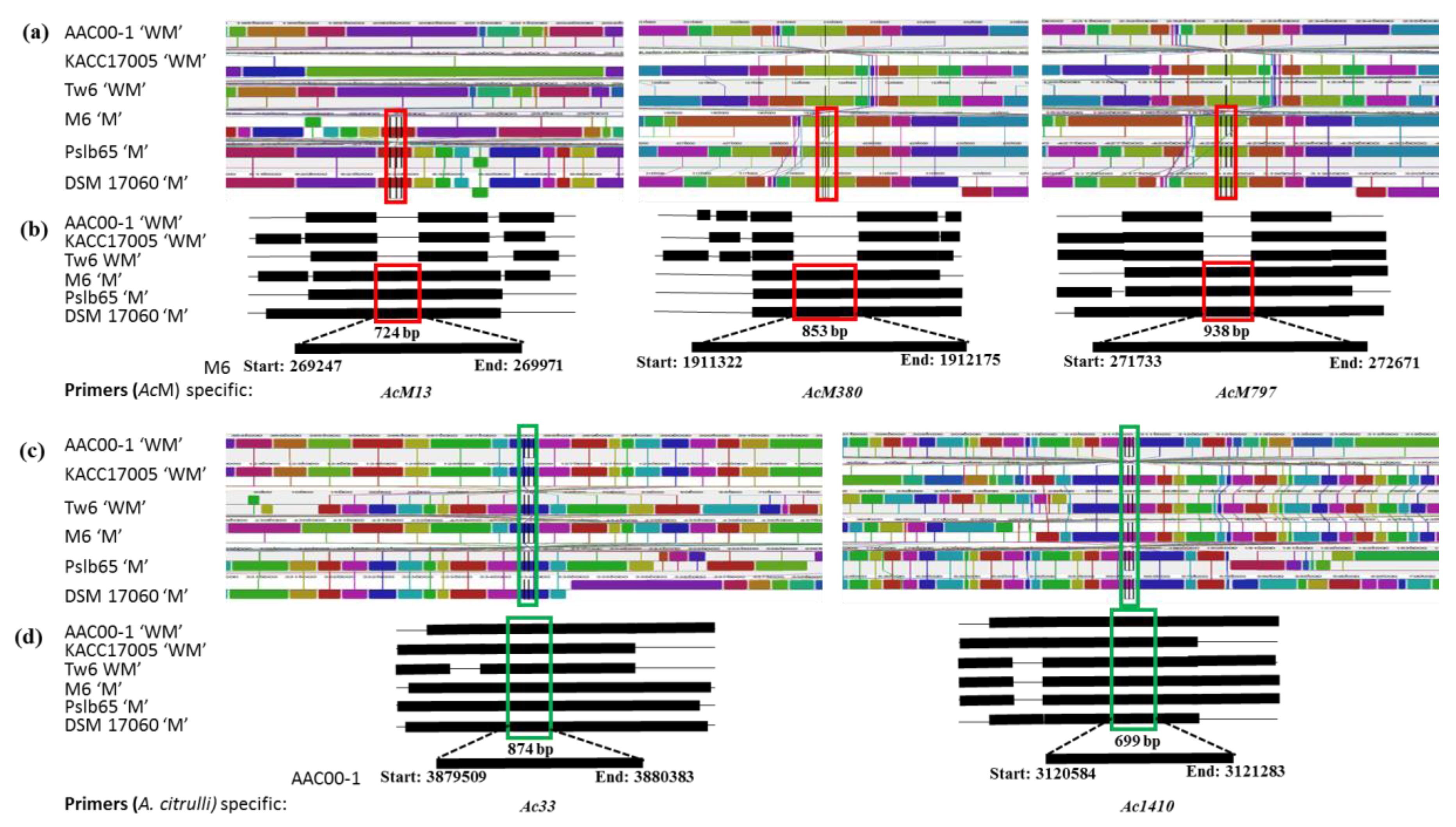
2.2. Whole-Genome Alignment Allows Design of Melon Host-Specific and A. citrulli Strain Specific Primers
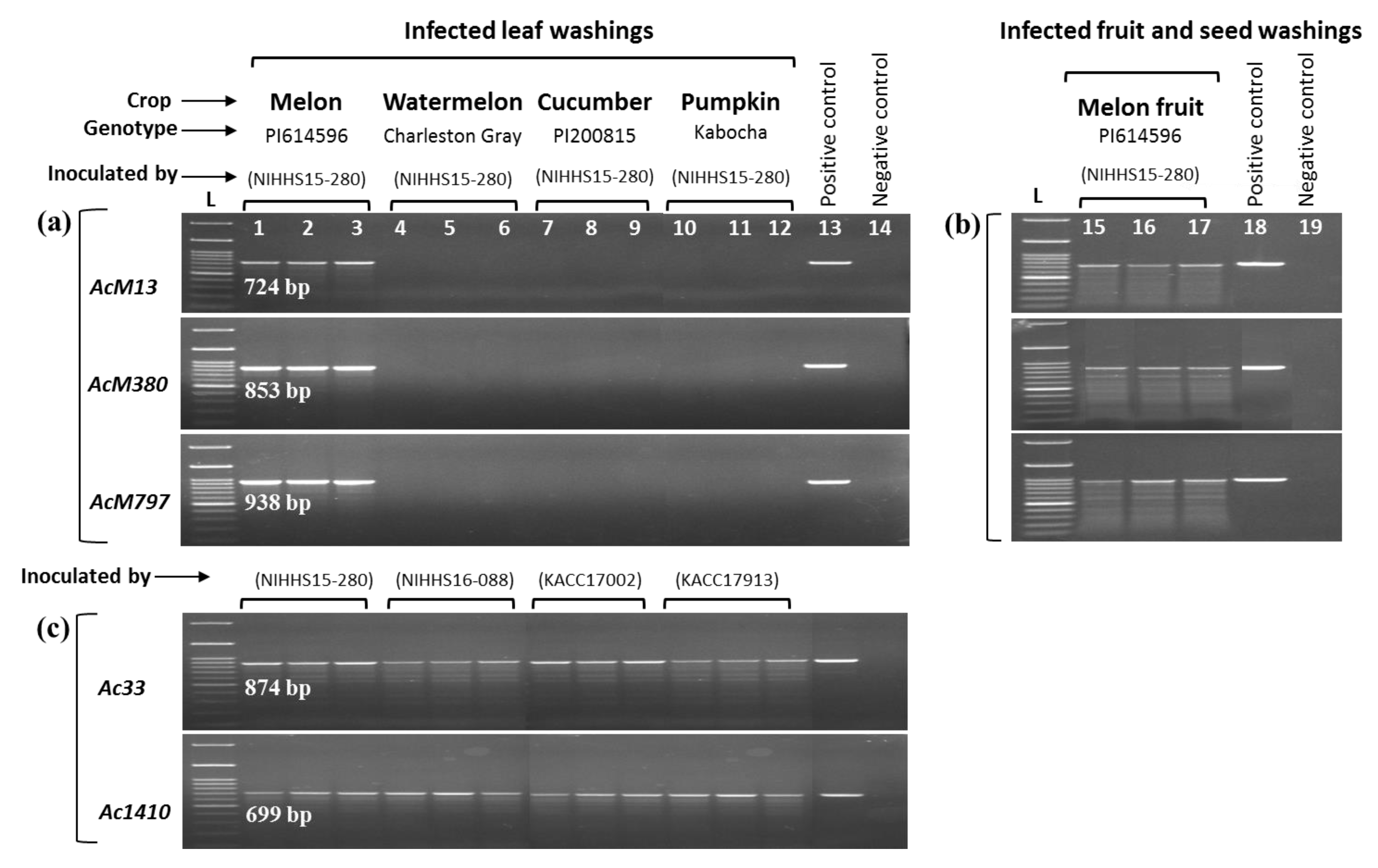
2.3. PCR-Based Assays Show Specificity of Primers
2.4. Bio-PCR Shows Sensitivity of the Primers
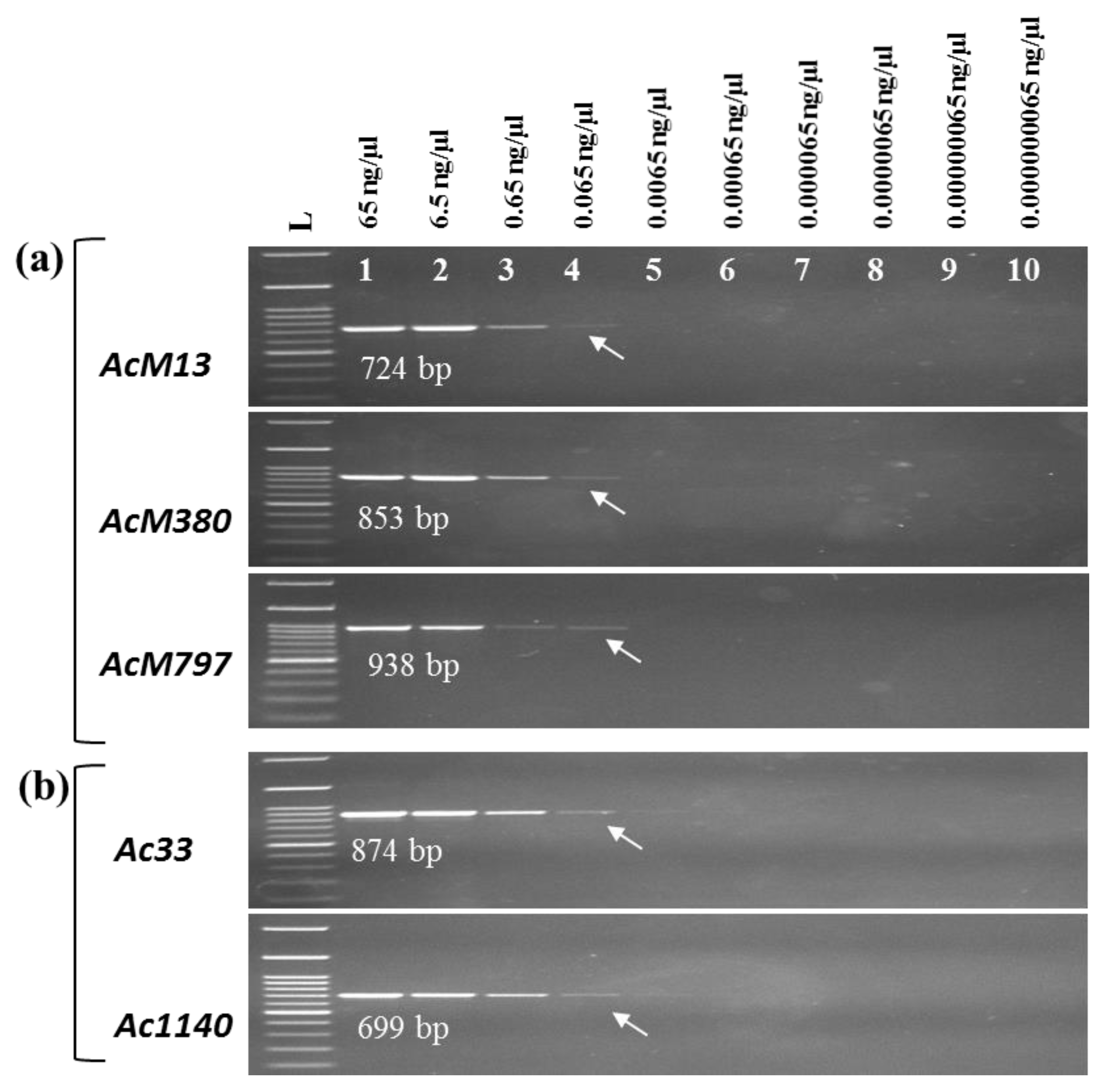
2.5. Detection Limits of the PCR Assays
3. Discussion
4. Materials and Methods
4.1. Retrieval and Alignment of Whole Genomes of A. citrulli Strains
4.2. Design of Primers for Detection of A. citrulli Strains
4.3. Bacterial Strains, Growing Conditions, and Inoculum Preparation
4.4. Extraction of Bacterial Genomic DNA
4.5. Specificity of Primers in PCR Assays
4.6. Pathogenicity Test of Bacterial Strains Using Cross Inoculation Assay
4.7. Bio-PCR Assay
4.8. Detection Limit of Bacterial DNA in the PCR Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schaad, N.W.; Postnikova, E.; Sechler, A.; Claflin, L.E.; Vidaver, A.K.; Jones, J.B.; Agarkova, I.; Ignatov, A.; Dickstein, E.; Ramundo, B.A. Reclassification of subspecies of Acidovorax avenae as A. Avenae (Manns 1905) emend., A. cattleyae (Pavarino, 1911) comb. nov., A. citrulli Schaad et al., 1978) comb. nov., and proposal of A. oryzae sp. nov. Syst. Appl. Microbiol. 2008, 31, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Webb, R.; Goth, R. A seedborne bacterium isolated from watermelon. Plant Dis. Report. 1965, 49, 818–821. [Google Scholar]
- Bahar, O.; Burdman, S. Bacterial fruit blotch: a threat to the cucurbit industry. Isr. J. Plant Sci. 2010, 58, 19–31. [Google Scholar] [CrossRef]
- Burdman, S.; Walcott, R. Acidovorax citrulli: Generating basic and applied knowledge to tackle a global threat to the cucurbit industry. Mol. Plant Pathol. 2012, 13, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.M.; Souza, R.M.; Yan, L.; Júnior, R.S.; Medeiros, F.H.; Walcott, R.R. Strains of the group I lineage of Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbitaceous crops, are predominant in Brazil. Phytopathology 2016, 106, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Santos, V. A new bacterial disease on watermelon in the Mariana Islands. Phytopathology 1988, 78, 1–9. [Google Scholar]
- Schaad, N.W.; Sowell, G.; Goth, R.W.; Colwell, R.R.; Webb, R.E. Pseudomonas pseudoalcaligenes subsp. citrulli subsp. nov. Int. J. Syst. Evol. Micr. 1978, 28, 117–125. [Google Scholar] [CrossRef]
- Latin, R.; Rane, K. Bacterial fruit blotch of watermelon in Indiana. Plant Dis. 1990, 74. [Google Scholar] [CrossRef]
- Schaad, N.; Postnikova, E.; Randhawa, P. Emergence of Acidovorax avenae subsp. citrulli as a crop threatening disease of watermelon and melon. In Pseudomonas syringae and Related Pathogens; Iacobellis, N.S., Collmer, A., Hutcheson, S.W., Mansfield, J.W., Morris, C.E., Murillo, J., Schaad, N.W., Stead, D.E., Surico, G., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 573–581. [Google Scholar]
- Somodi, G.C.; Jones, J.; Hopkins, D.; Stall, R.; Kucharek, T.; Hodge, N.; Watterson, J. Occurrence of a bacterial watermelon fruit blotch in Florida. Plant Dis. 1991, 75, 1053–1056. [Google Scholar] [CrossRef]
- Hopkins, D. The Hypothetical Exam Question Becomes Reality. Plant Dis. 1995, 79, 761–765. [Google Scholar]
- Isakeit, T.; Black, M.; Barnes, L.; Jones, J. First report of infection of honeydew with Acidovorax avenae subsp. citrulli. Plant Dis. 1997, 81, 694. [Google Scholar] [CrossRef] [PubMed]
- Langston, D., Jr.; Walcott, R.; Gitaitis, R.; Sanders, F., Jr. First report of a fruit rot of pumpkin caused by Acidivorax avenae subsp. citrulli in Georgia. Plant Dis. 1999, 83, 199. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.; Horlock, C. First report of Acidovorax avenae subsp. citrulli as a pathogen of Gramma in Australia. Plant Dis. 2002, 86, 1406. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.; O’Brien, R.; Abbott, D. First report of Acidovorax avenae subsp. citrulli as a pathogen of cucumber. Plant Dis. 1999, 83, 965. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.-L.; Zhang, G.-Q.; Liu, H.; Lou, M.-M.; Tian, W.-X.; Li, B.; Zhou, X.-P.; Zhu, B.; Jin, G.-L. Genome sequence of the rice-pathogenic bacterium Acidovorax avenae subsp. avenae RS-1. J. Bacteriol. 2011, 193, 5013–5014. [Google Scholar] [CrossRef]
- Sowell, G., Jr. A bacterial disease causing severe damage to susceptible plant introductions of muskmelon [Cucumis melo]. Plant Dis. (USA) 1981. [Google Scholar] [CrossRef]
- Conceição, C.S.; Felix, K.C.S.; Mariano, R.L.; Medeiros, E.V.; Souza, E.B. Combined effect of yeast and silicon on the control of bacterial fruit blotch in melon. Sci. Hortic. 2014, 174, 164–170. [Google Scholar] [CrossRef]
- De Melo, E.A.; Rosa de Lima, R.M.; Laranjeira, D.; dos Santos, L.A.; de Omena Gusmão, L.; de Souza, E.B. Efficacy of yeast in the biocontrol of bacterial fruit blotch in melon plants. Trop. Plant Pathol. 2015, 40, 56–64. [Google Scholar] [CrossRef]
- Sales Júnior, R.; Menezes, J. Mapeamento das doenças fúngicas, bacterianas e viróticas do cultivo do melão no Estado do RN. Mossoró. Escola Superior de Agricultura de Mossoró 2001. [Google Scholar]
- Noh, J.; Kim, J.-H.; Lim, J.H.; Kim, T.B.; Seong, M.H.; Jung, G.T.; Kim, J.M.; Cheong, S.-S.; Oh, N.K.; Lee, W.-H. Occurrence of diseases and case of clinical diagnosis on watermelon in South Korea, 2008–2012. Res. Plant Dis. 2014, 20, 8–14. [Google Scholar] [CrossRef]
- Bahar, O.; Kritzman, G.; Burdman, S. Bacterial fruit blotch of melon: screens for disease tolerance and role of seed transmission in pathogenicity. Eur. J. Plant Pathol. 2009, 123, 71–83. [Google Scholar] [CrossRef]
- Burdman, S.; Kots, N.; Kritzman, G.; Kopelowitz, J. Molecular, physiological, and host-range characterization of Acidovorax avenae subsp. citrulli isolates from watermelon and melon in Israel. Plant Dis. 2005, 89, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Feng, J.-J.; Hu, J.; Zhao, W.-J. Selective detection of viable seed-borne Acidovorax citrulli by real-time PCR with propidium monoazide. Sci. Rep. 2016, 6, 35457. [Google Scholar] [CrossRef] [PubMed]
- Schaad, N.W.; Sechler, A. An improved semiselective agar medium for Acidovorax avenae subsp. citrulli. Phytopathology 1999, 89 (Suppl. 6), 68–69. [Google Scholar]
- Zhao, T.; Feng, J.; Sechler, A.; Randhawa, P.; Li, J.; Schaad, N.W. An improved assay for detection of Acidovorax citrulli in watermelon and melon seed. Seed Sci. Technol. 2009, 37, 337–349. [Google Scholar] [CrossRef]
- Venette, J.; Lamppa, R.; Albaugh, D.; Nayes, J. Presumptive procedure (dome test) for detection of seedborne bacterial pathogens in dry beans. Plant Dis. 1987, 71, 984–990. [Google Scholar] [CrossRef]
- Feng, J.; Xu, Y.; Li, J.; Schaad, N. Comparison of immunostrip and real-time fluorescent PCR (TaqMan) for detection of Acidovorax avenae subsp. citrulli, the causal agent of bacterial fruit blotch of watermelon. Acta Phytopathol. Sin. 2006, 36, 102–108. [Google Scholar]
- Himananto, O.; Thummabenjapone, P.; Luxananil, P.; Kumpoosiri, M.; Hongprayoon, R.; Kositratana, W.; Gajanandana, O. Novel and Highly Specific Monoclonal Antibody to Acidovorax citrulli and Development of ELISA-Based Detection in Cucurbit Leaves and Seed. Plant Dis. 2011, 95, 1172–1178. [Google Scholar] [CrossRef]
- Horuz, S.; Cetinkaya-Yildiz, R.; Mirik, M.; Aysan, Y. Occurrence, isolation, and identification of Acidovorax citrulli from Melon in Turkey. Plant Protect. Sci. 2014, 50, 179–183. [Google Scholar] [CrossRef]
- Kuo, S.-Y.; Lin, Y.-C.; Lai, Y.-C.; Liao, J.-T.; Hsu, Y.-H.; Huang, H.-C.; Hu, C.-C. Production of fluorescent antibody-labeling proteins in plants using a viral vector and the application in the detection of Acidovorax citrulli and Bamboo mosaic virus. PLoS ONE 2018, 13, e0192455. [Google Scholar] [CrossRef]
- Matsuura, T.; Shirakawa, T.; Sato, M.; Inoue, Y.; Azegami, K. Detection and isolation of Acidovorax avenae subsp. citrulli from watermelon [Citrullus lanatus] seeds using membrane filtration immunostaining. Jpn. J. Phytopathol. (Japan) 2008. [Google Scholar] [CrossRef]
- Melo, L.A.; Tebaldi, N.D.; Mehta, A.; Marques, A.S.A. Comparing Acidovorax citrulli strains from melon and watermelon: Phenotypic characteristics, pathogenicity and genetic diversity. Trop. Plant Pathol. 2014, 39, 154–162. [Google Scholar] [CrossRef]
- Puttharugsa, C.; Wangkam, T.; Huangkamhang, N.; Gajanandana, O.; Himananto, O.; Sutapun, B.; Amarit, R.; Somboonkaew, A.; Srikhirin, T. Development of surface plasmon resonance imaging for detection of Acidovorax avenae subsp. citrulli (Aac) using specific monoclonal antibody. Biosens. Bioelectron. 2011, 26, 2341–2346. [Google Scholar] [CrossRef]
- Walcott, R.; Fessehaie, A.; Castro, A. Differences in pathogenicity between two genetically distinct groups of Acidovorax avenae subsp. citrulli on cucurbit hosts. J. Phytopathol. 2004, 152, 277–285. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Q.; Li, B.; Liu, B.; Wu, G.; Ibrahim, M.; Xie, G.; Li, H.; Sun, G. Differentiation in MALDI-TOF MS and FTIR spectra between two closely related species Acidovorax oryzae and Acidovorax citrulli. BMC Microbiol. 2012, 12, 182. [Google Scholar] [CrossRef]
- Zeng, H.; Guo, W.; Liang, B.; Li, J.; Zhai, X.; Song, C.; Zhao, W.; Fan, E.; Liu, Q. Self-paired monoclonal antibody lateral flow immunoassay strip for rapid detection of Acidovorax avenae subsp. citrulli. Anal. Bioanal. Chem. 2016, 408, 6071–6078. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kim, M.S.; Qin, J.; Park, E.; Song, Y.-R.; Oh, C.-S.; Cho, B.-K. Raman Hyperspectral Imaging for Detection of Watermelon Seeds Infected with Acidovorax citrulli. Sensors (Basel, Switzerland) 2017, 17, 2188. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Zhang, D.; Zhai, X.; Wang, S.; Liu, Q. Enhancing the immunofluorescent sensitivity for detection of Acidovorax citrulli using fluorescein isothiocyanate labeled antigen and antibody. Anal. Bioanal. Chem. 2018, 410, 71–77. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, Q.; Zhu, S.-F.; Zhao, W.-J.; Liu, F.-Q. Rapid on-site detection of Acidovorax citrulli by cross-priming amplification. Mol. Cell. Probes 2012, 26, 175–176. [Google Scholar] [CrossRef]
- Yan, L.; Zhao, Y.; Zhou, J.; Chen, S.; Bai, S.; Tian, Y.; Gong, W.; Hu, B. Rapid and sensitive detection of Acidovorax citrulli in cucurbit seeds by visual loop-mediated isothermal amplification assay. J. Phytopathol. 2019, 167, 10–18. [Google Scholar] [CrossRef]
- Giovanardi, D.; Sutton, S.A.; Stefani, E.; Walcott, R.R. Factors influencing the detection of Acidovorax citrulli in naturally contaminated cucurbitaceous seeds by PCR-based assays. Seed Sci. Technol. 2018, 46, 93–106. [Google Scholar] [CrossRef]
- Feng, J.-J.; Kim, J.-Y.; Liu, X.-L.; Schaad, N.W.; Li, J. Differentiation of live and dead cell of bacterial plant pathogen in polymerase chain reaction assays using a DNA binding dye. Chem. J. Chin. U 2008, 29, 944–948. [Google Scholar]
- Cho, M.S.; Park, D.H.; Ahn, T.-Y.; Park, D.S. Rapid and specific detection of Acidovorax avenae subsp. citrulli using SYBR green-based real-time PCR amplification of the YD-repeat protein gene. J. Microbiol. Biotechnol. 2015, 25, 1401–1409. [Google Scholar] [CrossRef]
- Choi, O.; Cho, S.K.; Kang, B.; Cho, J.; Park, J.; Lee, Y.; Kim, J. Two genetically distinct groups of Acidovorax citrulli are present in watermelon-growing fields in Korea. J. Agric. Life Sci. 2016, 50, 53–59. [Google Scholar] [CrossRef]
- Rui, S.; Qing, L.; Yali, L.; Zhaohui, W.; Junping, W.; Liping, Z.; Jianxiong, H.; Zhijie, L. Rapid Immuno-PCR Detection of Acidovorax avenae subsp. citrulli. Plant Quar. 2009, 2. [Google Scholar]
- Xu, F.; Wang, X.; Xie, G.; Su, T.; Yu, S. Detection of Acidovorax avenae subsp. citrulli from seeds of watermelon by immuno-capture PCR. J. Fruit Sci. 2008, 2. [Google Scholar]
- Walcott, R.; Gitaitis, R. Detection of Acidovorax avenae subsp. citrulli in watermelon seed using immunomagnetic separation and the polymerase chain reaction. Plant Dis. 2000, 84, 470–474. [Google Scholar] [CrossRef]
- Amagliani, G.; Omiccioli, E.; Campo, A.; Bruce, I.J.; Brandi, G.; Magnani, M. Development of a magnetic capture hybridization-PCR assay for Listeria monocytogenes direct detection in milk samples. J. Appl. Microbiol. 2006, 100, 375–383. [Google Scholar] [CrossRef]
- Eckshtain-Levi, N.; Shkedy, D.; Gershovits, M.; Da Silva, G.M.; Tamir-Ariel, D.; Walcott, R.; Pupko, T.; Burdman, S. Insights from the genome sequence of Acidovorax citrulli M6, a group I strain of the causal agent of bacterial fruit blotch of cucurbits. Front. Microbiol. 2016, 7, 430. [Google Scholar] [CrossRef] [PubMed]
- Slovareva, O.Y.; Kornev, K.P.; Matyashova, G.N.; Stakheev, A.A.; Prikhodko, S.I. Recommended Procedure for Detection and Identification Acidovorax citrulli in Seeds; AIP Publishing: Melville, NY, USA, 2019; Volume 2063, p. 030020. [Google Scholar]
- Song, J.Y.; Oo, M.M.; Park, S.Y.; Seo, M.W.; Lee, S.-C.; Jeon, N.B.; Nam, M.H.; Lee, Y.S.; Kim, H.G.; Oh, S.-K. Analysis of intraspecific genetic diversity in Acidovorax citrulli causing bacterial fruit blotch on cucurbits in Korea. Korean J. Agric. Sci. 2018, 45, 575–582. [Google Scholar]
- Wang, T.; Sun, B.; Yang, Y.; Zhao, T. Genome sequence of Acidovorax citrulli group 1 strain pslb65 causing bacterial fruit blotch of melons. Genome Announc. 2015, 3, e00315–e00327. [Google Scholar] [CrossRef] [PubMed]
- Copeland, A.; Lucas, S.; Lapidus, A.; Barry, K.; Detter, J.; Glavina del Rio, T.; Dalin, E.; Tice, H.; Pitluck, S.; Kiss, H. Complete Sequence of Acidovorax avenae subsp. citrulli AAC00-1; US DOE Joint Genome Institute: Walnut Creek, CA, USA, 2006. [Google Scholar]
- Park, H.-J.; Seong, H.J.; Sul, W.J.; Oh, C.-S.; Han, S.-W. Complete genome sequence of Acidovorax citrulli strain KACC17005, a causal agent for bacterial fruit blotch on watermelon. J. Microbiol. 2017, 53, 340–341. [Google Scholar]
- Wang, T.; Yang, Y.; Zhao, T. Genome sequence of a copper-resistant strain of Acidovorax citrulli causing bacterial fruit blotch of melons. Genome Announc. 2015, 3, e00310–e00315. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.J.; Li, J.Q.; Walcott, R.R.; Zhang, G.M.; Luo, L.X.; Kang, L.; Zheng, Y.; Schaad, N.W. Advances in detection of Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbits. Seed Sci. Technol. 2013, 41, 1–15. [Google Scholar] [CrossRef]
- Ha, Y.; Fessehaie, A.; Ling, K.; Wechter, W.; Keinath, A.; Walcott, R. Simultaneous detection of Acidovorax avenae subsp. citrulli and Didymella bryoniae in cucurbit seedlots using magnetic capture hybridization and real-time polymerase chain reaction. Phytopathology 2009, 99, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, H.; Sato, M.; Suzuki, A. Detection of Acidovorax avenae subsp. citrulli using PCR and MALDI-TOF MS. J. Electrophor. 2012, 56, 13–17. [Google Scholar] [CrossRef]
- Song, W.Y.; Kim, H.M.; Hwang, C.Y.; Schaad, N.W. Detection of Acidovorax avenae ssp. avenae in Rice Seeds Using BIO-PCR. J. Phytopathol. 2004, 152, 667–676. [Google Scholar] [CrossRef]
- Zivanovic, M.; Walcott, R. Further characterization of genetically distinct groups of Acidovorax citrulli strains. Phytopathology 2016, 107, 29–35. [Google Scholar] [CrossRef]
- Wang, T.; Yang, Y.; Zhao, T. Genome Sequence of a Copper-Resistant Strain of span class “named-content genus-species” “named-content-1” Acidovorax citrulli span Causing Bacterial Fruit Blotch of Melons. Genome Announc. 2015, 3, e00310–e00315. [Google Scholar] [CrossRef]
- Varghese, N.A.S. Acidovorax citrulli Strain DSM 17060, Whole Genome Shotgun Sequencing Project. Available online: https://www.ncbi.nlm.nih.gov/nuccore/FNEY00000000 (accessed on 15 February 2019).
- Yan, S.; Yang, Y.; Wang, T.; Zhao, T.; Schaad, N.W. Genetic diversity analysis of Acidovorax citrulli in China. Eur. J. Plant Pathol. 2013, 136, 171–181. [Google Scholar] [CrossRef]
- Adhikari, M.; Yadav, D.R.; Kim, S.W.; Um, Y.H.; Kim, H.S.; Lee, S.C.; Song, J.Y.; Kim, H.G.; Lee, Y.S. Biological control of bacterial fruit blotch of watermelon pathogen (Acidovorax citrulli) with rhizosphere associated bacteria. Plant Pathol. J. 2017, 33, 170. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Midani, A.; Lee, Y.S.; Kang, S.-W.; Kim, M.-K.; Choi, T.-J. First Isolation and Molecular Characterization of Bacteriophages Infecting Acidovorax citrulli, the Causal Agent of Bacterial Fruit Blotch. Plant Pathol. J. 2018, 34, 59. [Google Scholar] [PubMed]
- Bahar, O.; Efrat, M.; Hadar, E.; Dutta, B.; Walcott, R.; Burdman, S. New subspecies-specific polymerase chain reaction-based assay for the detection of Acidovorax avenae subsp. citrulli. Plant Pathol. 2008, 57, 754–763. [Google Scholar] [CrossRef]
- Zhong, J.; Lin, Z.-Y.; Ma, Y.-M.; Gao, B.-D.; Liu, H.-Q.; Zhao, T.-C.; Schaad, N.W. Rapid Discrimination between Groups I and II of Acidovorax citrulli Using a Primer Pair Specific to a pil L Gene. J. Phytopathol. 2016, 164, 558–562. [Google Scholar] [CrossRef]
- Song, J.Y.; Park, S.Y.; Seo, M.W.; Nam, M.H.; Lim, H.S.; Lee, S.-C.; Lee, Y.S.; Kim, H.G. Genetic characteristics of Acidovorax citrulli population causing bacterial fruit blotch against cucurbits in Korea. Res. Plant Dis. 2015, 21, 82–88. [Google Scholar] [CrossRef]
- Walcott, R.R.; Gitaitis, R.D.; Castro, A.C. Role of Blossoms in Watermelon Seed Infestation by Acidovorax avenae subsp. citrulli. Phytopathology 2003, 93, 528–534. [Google Scholar] [CrossRef]
- Oya, H.; Nakagawa, H.; Saito, N.; Uematsu, H.; Ohara, T. Detection of Acidovorax avenae subsp. citrulli from seed using LAMP method. Jpn. J. Phytopathol. (Japan) 2008. [Google Scholar] [CrossRef]
- Tian, Y.; Xu, J.; Zhao, Y.; Li, X.; Hu, B.; Liu, F. Specific detection of Acidovorax avenae subsp. citrulli by PCR. Jiangsu J. Agric. Sci. 2010, 26, 512–516. [Google Scholar]
- Rubel, M.H.; Robin, A.H.K.; Natarajan, S.; Vicente, J.G.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Whole-Genome re-alignment facilitates development of specific molecular markers for races 1 and 4 of Xanthomonas campestris pv. campestris, the cause of black rot disease in Brassica oleracea. Int. J. Mol. Sci. 2017, 18, 2523. [Google Scholar] [CrossRef]
- Rubel, M.H.; Hossain, M.R.; Nath, U.K.; Natarajan, S.; Lee, J.-H.; Jung, H.-J.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Development of a PCR test for detection of Xanthomonas campestris pv. raphani. Australas. Plant Pathol. 2019, 48, 179–182. [Google Scholar] [CrossRef]
- Rubel, M.H.; Natarajan, S.; Hossain, M.R.; Nath, U.K.; Afrin, K.S.; Lee, J.-H.; Jung, H.-J.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Pathovar specific molecular detection of Xanthomonas campestris pv. campestris, the causal agent of black rot disease in cabbage. Can. J. Plant Pathol. 2019. [Google Scholar] [CrossRef]
- Larrea-Sarmiento, A.; Dhakal, U.; Boluk, G.; Fatdal, L.; Alvarez, A.; Strayer-Scherer, A.; Paret, M.; Jones, J.; Jenkins, D.; Arif, M. Development of a genome-informed loop-mediated isothermal amplification assay for rapid and specific detection of Xanthomonas euvesicatoria. Sci. Rep. 2018, 8, 14298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Zhang, G.; Liu, X.; Wang, C.; Xu, J. Comparison of mitochondrial genomes provides insights into intron dynamics and evolution in the caterpillar fungus Cordyceps militaris. Fungal Genet. Biol. 2015, 77, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-Y.; Robin, A.H.K.; Natarajan, S.; Laila, R.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Race- and Isolate-specific Molecular Marker Development through Genome-Realignment Enables Detection of Korean Plasmodiophora brassicae Isolates, Causal agents of Clubroot Disease. Plant Pathol. J. 2018, 34, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.L.; Dassa, E.; Orelle, C.; Chen, J. Structure, function, and evolution of bacterial ATP-binding cassette systems. Microbiol. Mol. Biol. Rev. MMBR 2008, 72, 317–364. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.J.; Song, S.; Mason, K.; Pinkett, H.W. Selective substrate uptake: The role of ATP-binding cassette (ABC) importers in pathogenesis. Biochim. Biophys. Acta (BBA) Biomembr. 2018, 1860, 868–877. [Google Scholar] [CrossRef] [PubMed]
- De Buck, E.; Lammertyn, E.; Anné, J. The importance of the twin-arginine translocation pathway for bacterial virulence. Trends Microbiol. 2008, 16, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Lavander, M.; Ericsson, S.K.; Bröms, J.E.; Forsberg, A. The twin arginine translocation system is essential for virulence of Yersinia pseudotuberculosis. Infect. Immun. 2006, 74, 1768–1776. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.A.; Tullman-Ercek, D.; Georgiou, G. The Bacterial Twin-Arginine Translocation Pathway. Annu. Rev. Microbiol. 2006, 60, 373–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, W.; Itoh, T.; Maruyama, Y.; Mikami, B.; Murata, K. Hydration of vinyl ether groups by unsaturated glycoside hydrolases and their role in bacterial pathogenesis. Int. Microbiol. 2007, 10, 233–243. [Google Scholar] [PubMed]
- Olivera, I.E.; Fins, K.C.; Rodriguez, S.A.; Abiff, S.K.; Tartar, J.L.; Tartar, A. Glycoside hydrolases family 20 (GH20) represent putative virulence factors that are shared by animal pathogenic oomycetes, but are absent in phytopathogens. BMC Microbiol. 2016, 16, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjögren, J.; Collin, M. Bacterial glycosidases in pathogenesis and glycoengineering. Future Microbiol. 2014, 9, 1039–1051. [Google Scholar] [CrossRef]
- Wechter, W.P.; Levi, A.; Ling, K.-S.; Kousik, C.; Block, C.C. Identification of resistance to Acidovorax avenae subsp. citrulli among melon (Cucumis spp.) plant introductions. HortScience 2011, 46, 207–212. [Google Scholar] [CrossRef]
- Carvalho, F.C.; Santos, L.A.; Dias, R.C.; Mariano, R.L.; Souza, E.B. Selection of watermelon genotypes for resistance to bacterial fruit blotch. Euphytica 2013, 190, 169–180. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Scientific Name | Host Plant | SC |
|---|---|---|---|
| NIHHS15-280, KACC18782 | Acidovorax citrulli | Melon | South Korea |
| NIHHS16-088, KACC17000, KACC17001, KACC18785, KACC17005, KACC17909, KACC17910, KACC17911, KACC17912, KACC18649, KACC18783 | Acidovorax citrulli | Watermelon | |
| KACC17913 | Acidovorax citrulli | Pumpkin seed | |
| KACC17002, KACC18784 | Acidovorax citrulli | Cucumber | |
| KACC16207 | Acidovorax avenae subsp. avenae | Proso- millet | |
| ICMP13051 | Pseudomonas syringae pv. maculicola | Cabbage | New Zealand |
| NIHHS1326 | Didymella bryoniae | Cucumis melo | South Korea |
| SI | Strain | HP | GS (Mb) | GC% | Contigs | 16S rRNA | BioSample | SI (%) | GI (%) | References |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | AAC00-1 | Watermelon | 5.35277 | 68.53 | 1 | 4 | SAMN02598334 | 99.80 | 99.98 | [54] |
| 2 | KACC17005 | Watermelon | 5.34992 | 68.54 | 1 | 4 | SAMN07718226 | [55] | ||
| 3 | Tw6 | Watermelon | 5.07516 | 68.70 | 24 | 5 | SAMN03268415 | 90.48 | 99.52 | [62] |
| 4 | M6 | Melon | 4.82187 | 68.87 | 139 | 5 | SAMN04157986 | 91.36 | 99.63 | [50] |
| 5 | pslb65 | Melon | 4.90344 | 68.80 | 24 | 5 | SAMN03333326 | 92.04 | 99.54 | [53] |
| 6 | DSM 17060 | Melon | 4.84827 | 71 | 5 | SAMN04489709 | 92.04 | 99.51 | [63] |
| Primer Name | Primer Sequence | LCB | Genomic Location | Gene Description | Amplicon Size (bp) | Annealing Condition |
|---|---|---|---|---|---|---|
| AcM13 | F: TCGCGGGCCGTGATGTTCCG R: TGGACTTCGGGTGGGCCTTCA | 13 | (+) 269247 to 269971 | Glycosyl hydrolase | 724 | 65 °C for 45 s |
| AcM380 | F: GCATCCGGTGTGCTGCTGGA R: GAGATGTCAGAGTCGCACGGT | 380 | (+) 1911322 to 1912175 | ATP-binding protein | 853 | |
| AcM797 | F: AAGGCGGACATGGGTTGGCT R: CTGCGCCTGCGCCCACACCA | 797 | (+) 271733 to 272671 | Twin-arginine translocation pathway signal | 938 | |
| Ac33 | F: TCGATAAGGCCACCAAGTTC R: GACTGGGGTAACGTGGGGCT | 33 | (−) 3879509 to 3880383 | Hypothetical protein phosphatase | 874 | 63 °C for 30 s |
| Ac1410 | F: TAGCGCAGCCCCACCCAGTG R: CAAGGGCGACAAGATGATGT | 1410 | (+) 3120584 to 3121283 | Sugar-binding protein | 699 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.R.; Hossain, M.R.; Kim, H.-T.; Jesse, D.M.I.; Abuyusuf, M.; Jung, H.-J.; Park, J.-I.; Nou, I.-S. Development of Molecular Markers for Detection of Acidovorax citrulli Strains Causing Bacterial Fruit Blotch Disease in Melon. Int. J. Mol. Sci. 2019, 20, 2715. https://doi.org/10.3390/ijms20112715
Islam MR, Hossain MR, Kim H-T, Jesse DMI, Abuyusuf M, Jung H-J, Park J-I, Nou I-S. Development of Molecular Markers for Detection of Acidovorax citrulli Strains Causing Bacterial Fruit Blotch Disease in Melon. International Journal of Molecular Sciences. 2019; 20(11):2715. https://doi.org/10.3390/ijms20112715
Chicago/Turabian StyleIslam, Md. Rafiqul, Mohammad Rashed Hossain, Hoy-Taek Kim, Denison Michael Immanuel Jesse, Md. Abuyusuf, Hee-Jeong Jung, Jong-In Park, and Ill-Sup Nou. 2019. "Development of Molecular Markers for Detection of Acidovorax citrulli Strains Causing Bacterial Fruit Blotch Disease in Melon" International Journal of Molecular Sciences 20, no. 11: 2715. https://doi.org/10.3390/ijms20112715
APA StyleIslam, M. R., Hossain, M. R., Kim, H. -T., Jesse, D. M. I., Abuyusuf, M., Jung, H. -J., Park, J. -I., & Nou, I. -S. (2019). Development of Molecular Markers for Detection of Acidovorax citrulli Strains Causing Bacterial Fruit Blotch Disease in Melon. International Journal of Molecular Sciences, 20(11), 2715. https://doi.org/10.3390/ijms20112715