Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation



 and
and 
Abstract
:1. Introduction
2. Results
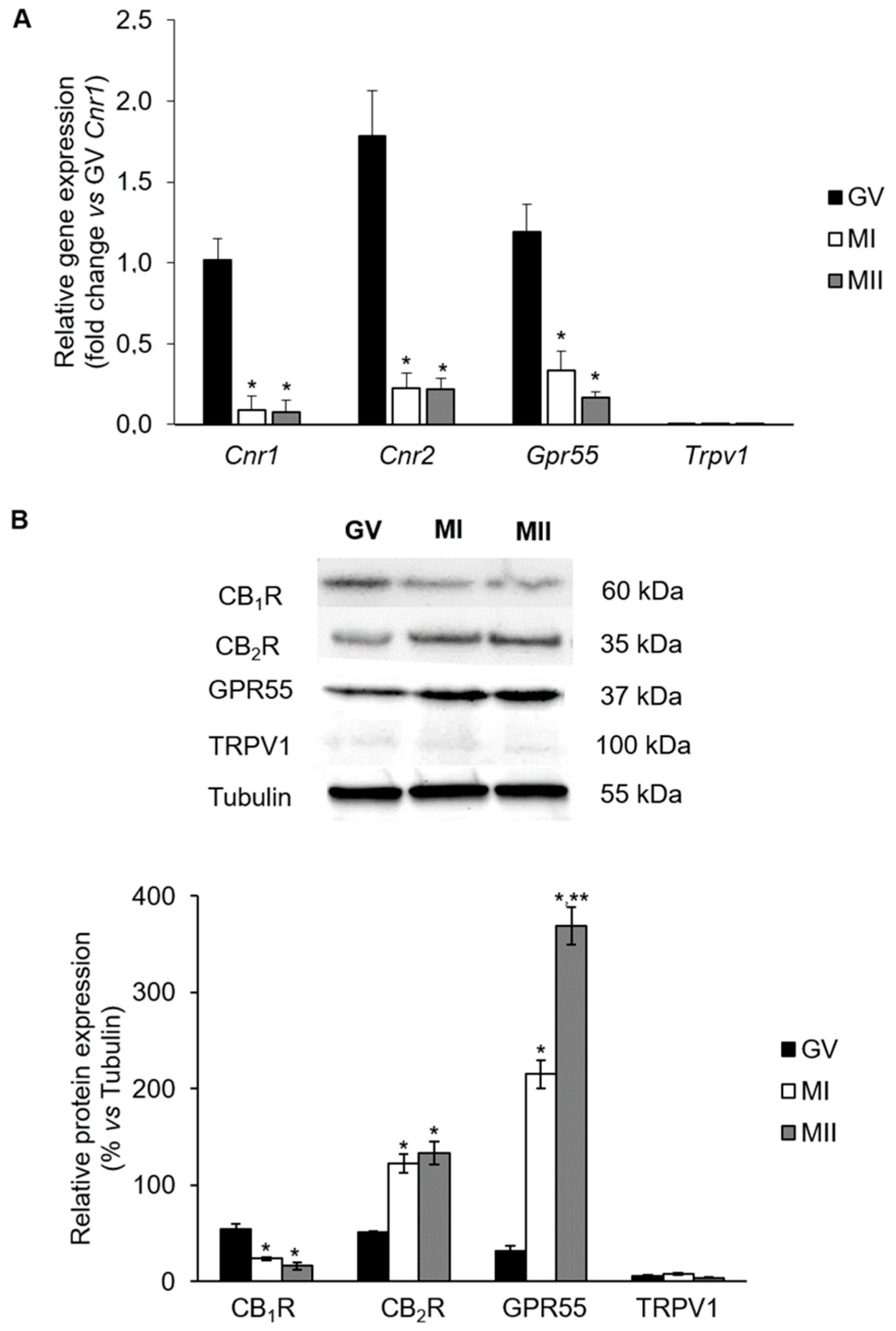
2.1. mRNA Levels of eCB-Binding Receptors in GV, MI, and MII Oocytes
2.2. Protein Levels of eCB-Binding Receptors in GV, MI, and MII Oocytes
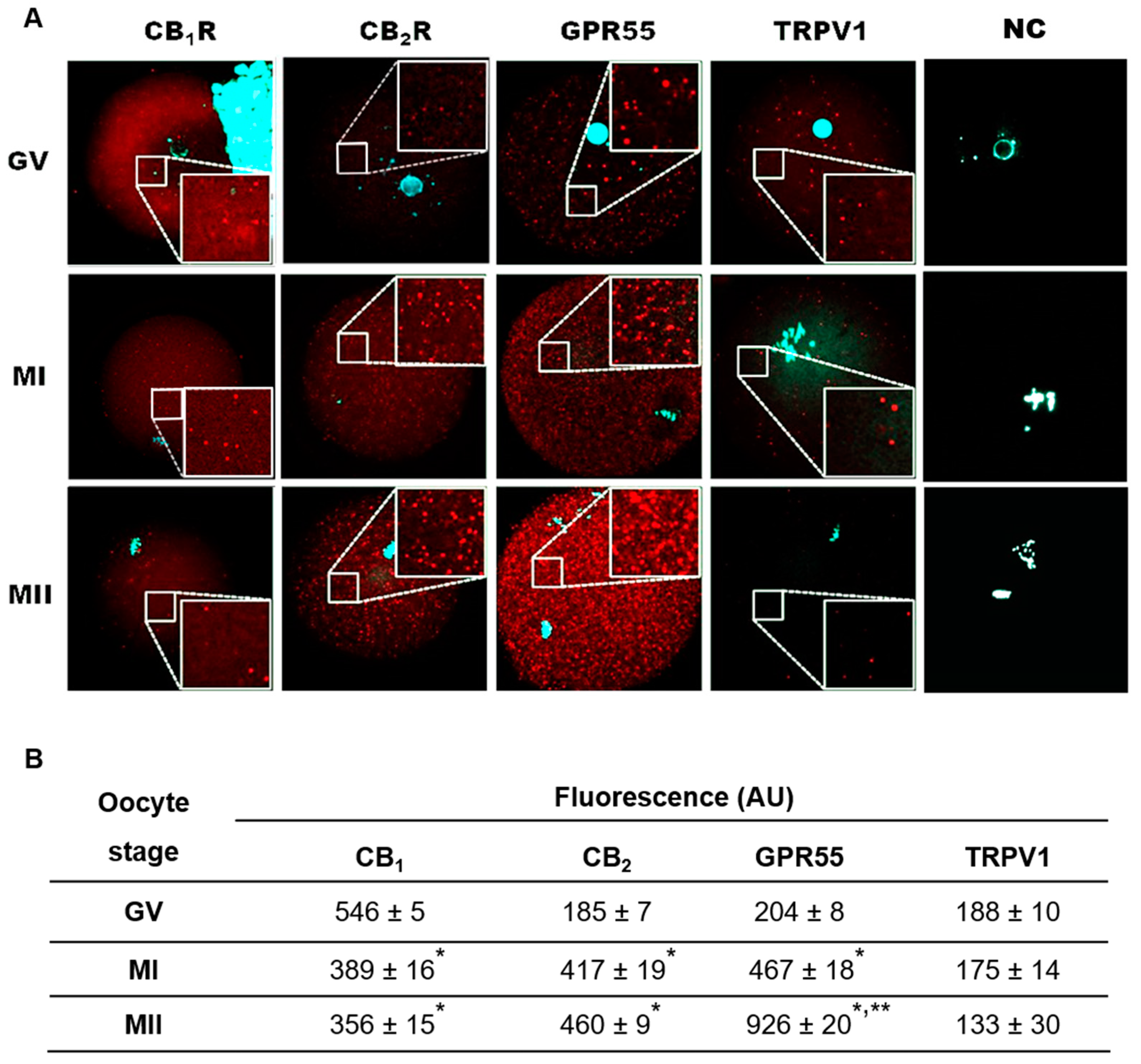
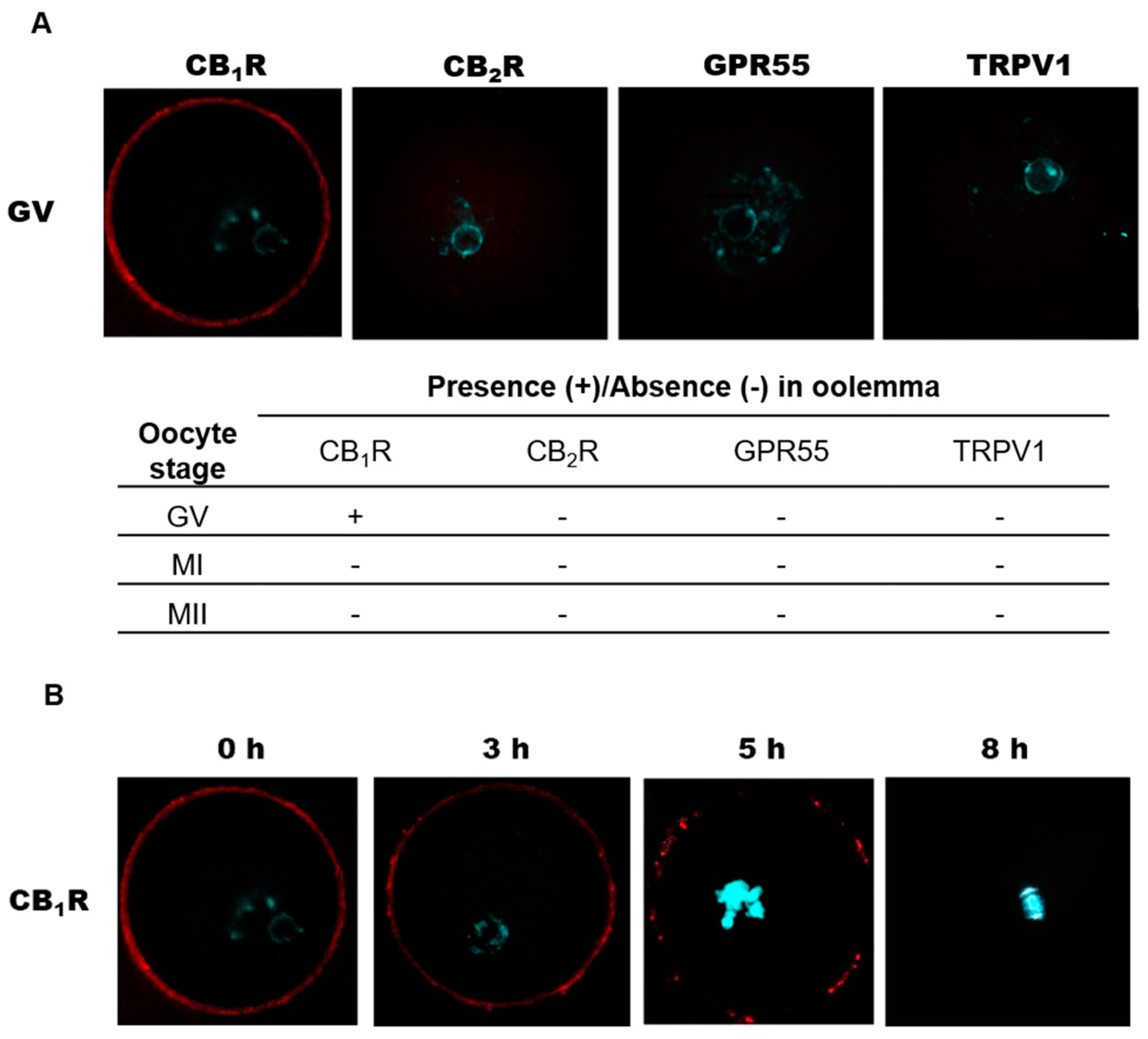
2.3. Localization of eCB-Binding Receptors in GV, MI, and MII Oocytes
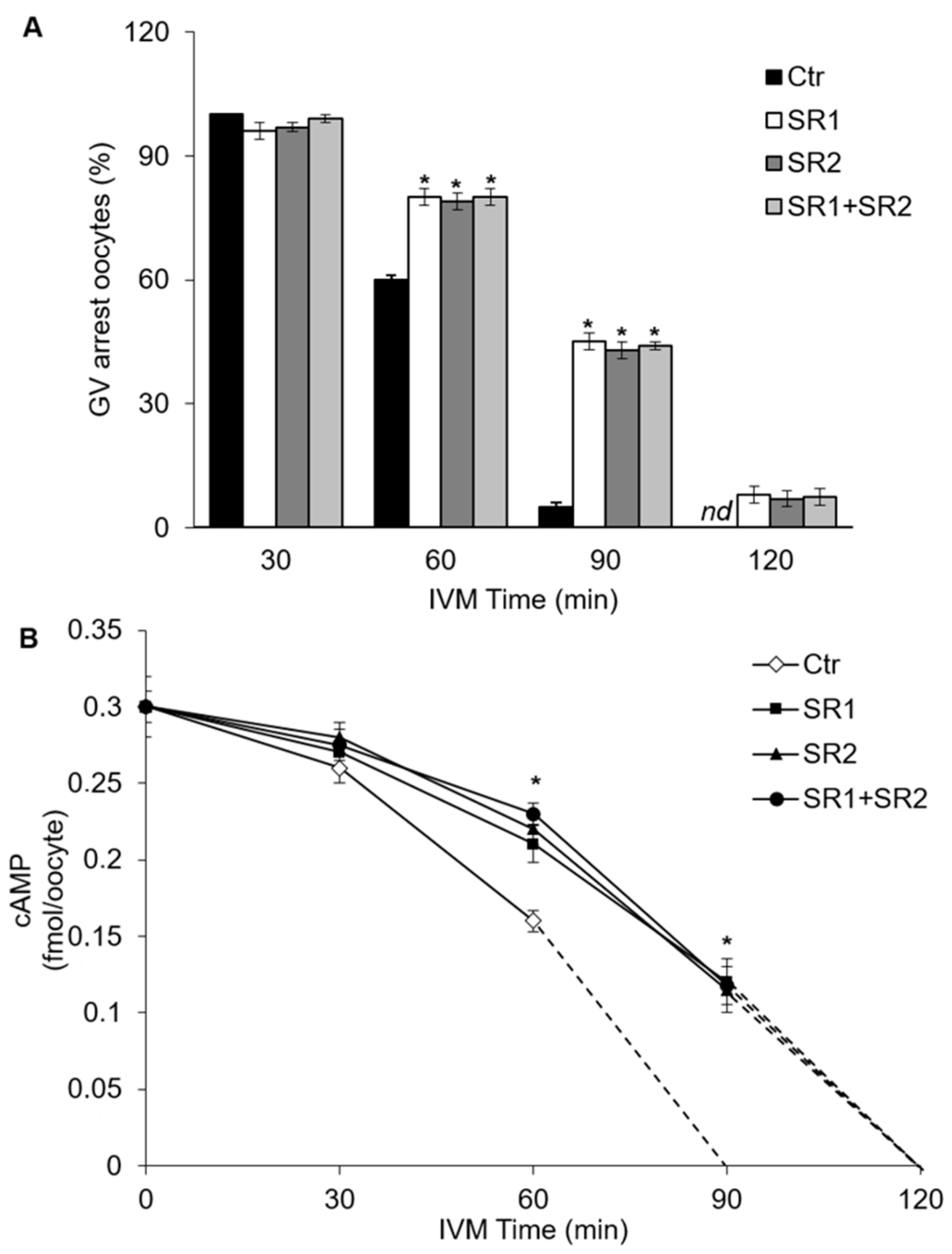
2.4. Effects of Receptor Antagonists on Intraoocyte cAMP Concentration
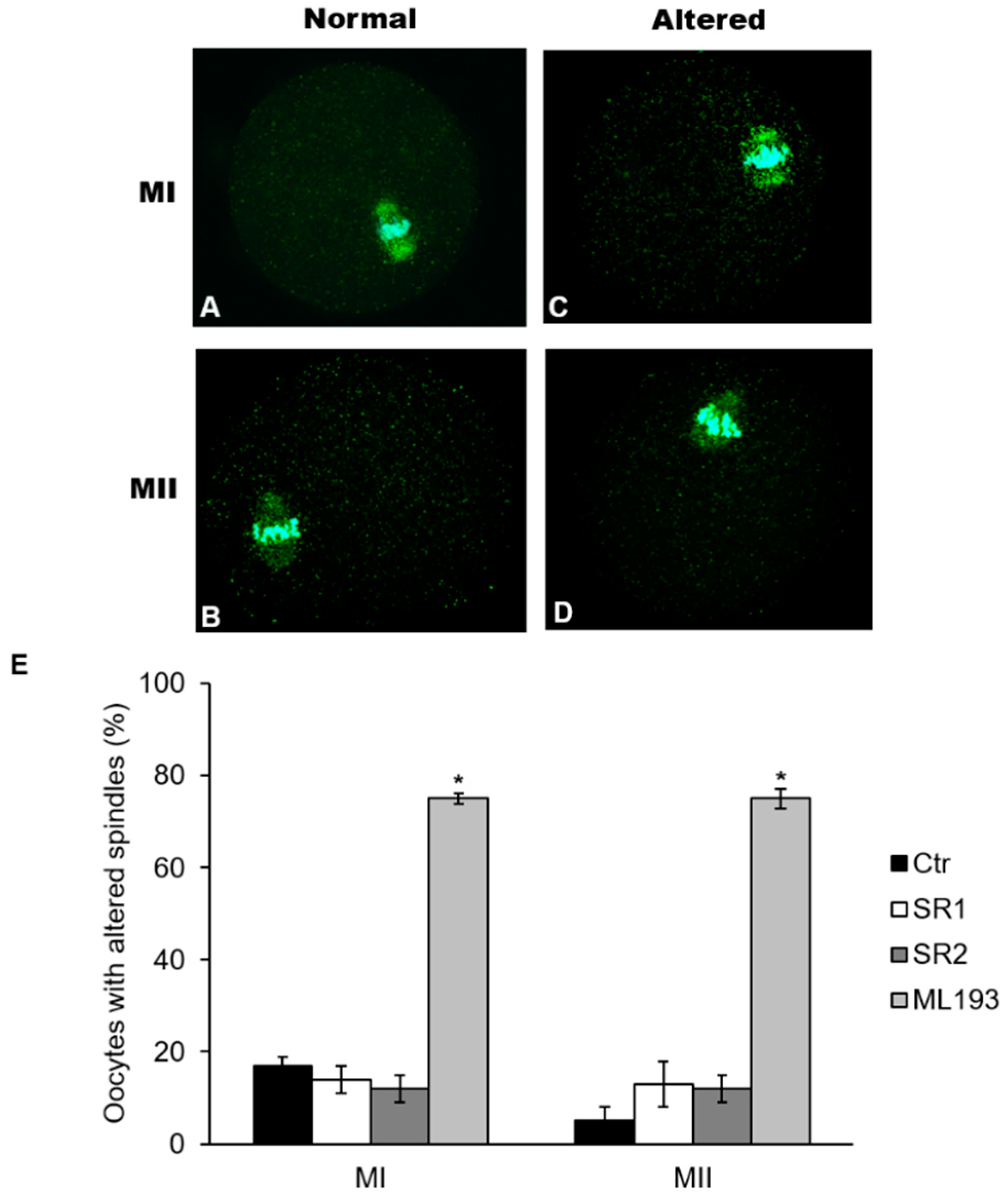
2.5. Effects of Receptor Antagonists on Polar Body I Emission and Spindle Morphology
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Ethical Approval
4.4. Collection of In Vivo Matured Oocytes
4.5. Quantitative Real-Time PCR Analysis
4.6. Western Blotting Analysis
4.7. Immunofluorescence
4.8. Effects of CB1R and CB2R Antagonists on Intraoocyte cAMP Content
4.9. Effects of Receptor Antagonists on Polar Body I Emission and Spindle Formation
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-AG | 2-arachidonoylglycerol |
| AEA | N-arachidonoylethanolamine |
| AU | Arbitrary units |
| BSA | Bovine Serum Albumin |
| cAMP | Cyclic adenosine monophosphate |
| CB1R | Cannabinoid receptor type-1 |
| CB2R | Cannabinoid receptor type-2 |
| CTR | Control |
| Cy-3 | Cyanine 3 bisacid |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMSO | Dimethyl sulfoxide |
| eCBs | Endocannabinoids |
| ECS | Endocannabinoid system |
| ERK 1/2 | Extracellular signal-regulated kinases 1/2 |
| FAAH | Fatty acid amide hydrolase |
| GCs | Granulosa cells |
| GPR3 | G protein-coupled receptor 3 |
| GPR55 | G-protein coupled receptor 55 |
| GRK | G protein-coupled receptor kinase |
| GV | Germinal vesicle |
| GVBD | Germinal vesicle breakdown |
| hCG | Human chorionic gonadotropin |
| HU-210 | 1,1-Dimethylheptyl-11-hydroxy-tetrahydrocannabinol |
| IP3 | Inositol trisphosphate |
| IVM | In vitro maturation |
| MI | Metaphase I |
| MII | Metaphase II |
| ML193 | N-[4-[[(3,4-dimethyl-5-isoxazolyl)amino]sulfonyl]phenyl]-6,8-dimethyl-2-(2-pyridinyl)-4-quinolinecarboxamide |
| NAPE-PLD | N-acyl phosphatidylethanolamine-specific phospholipase D |
| NC | Negative control |
| NuMA | Nuclear mitotic apparatus protein 1 |
| OCCs | Oocyte-cumulus complexes |
| OEA | N-oleoylethanolamine |
| OSE | Ovarian surface epithelium |
| PBI | Polar body I |
| PCOS | Polycystic ovary syndrome |
| PDE3A | Phosphodiesterase 3A |
| PEA | N-palmitoylethanolamine |
| PLC | Phospholipase C |
| PMSG | Pregnant mare serum gonadotropin |
| RhoA | Ras homolog gene family, member A |
| SR1 | 5-(4-Chloro-phenyl)-1-(2,4-dichloro-phenyl)-4-methyl-1H-pyrazole-3-carboxylic acid piperidin-1-ylamide hydrochloride |
| SR2 | 5-(4-Chloro-3-methylphenyl)-1-(4-methylbenzyl)-N-((1S,2S,4R)-1,3,3-trimethylbicyclo[2.2.1]heptan-2-yl)-1H-pyrazole-3-carboxamide |
| TRPV1 | Transient receptor potential vanilloid type 1 channel |
| ZF | Zona-free |
References
- Richards, J.S.; Russell, D.L.; Ochsner, S.; Hsieh, M.; Doyle, K.H.; Falender, A.E.; Sharma, S.C. Novel signaling pathways that control ovarian follicular development, ovulation, and luteinization. Recent Prog. Horm. Res. 2002, 57, 195–220. [Google Scholar] [CrossRef] [PubMed]
- Canipari, R.; Cellini, V.; Cecconi, S. The ovary feels fine when paracrine and autocrine networks cooperate with gonadotropins in the regulation of folliculogenesis. Curr. Pharm. Des. 2012, 18, 245–255. [Google Scholar] [PubMed]
- Wang, H.; Dey, S.K.; Maccarrone, M. Jekyll and hyde: Two faces of cannabinoid signaling in male and female fertility. Endocr. Rev. 2006, 27, 427–448. [Google Scholar] [CrossRef] [PubMed]
- Brents, L.K. Marijuana, the endocannabinoid system and the female reproductive system. Yale J. Biol. Med. 2016, 89, 175–191. [Google Scholar] [PubMed]
- Maccarrone, M.; Valensise, H.; Bari, M.; Lazzarin, N.; Romanini, C.; Finazzi-Agrò, A. Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet 2000, 355, 1326–1329. [Google Scholar] [CrossRef]
- Maccarrone, M.; Fride, E.; Bisogno, T.; Bari, M.; Cascio, M.G.; Battista, N.; Di Marzo, V. Up-regulation of the endocannabinoid system in the uterus of leptin knockout (ob/ob) mice and implications for fertility. Mol. Hum. Reprod. 2005, 11, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.; McPartland, J.M.; Glass, M. Cannabis, cannabinoids and reproduction. Prostaglandins Leukot. Essent. Fatty Acids 2004, 70, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Rossi, G.; Castellucci, A.; D’Andrea, G.; Maccarrone, M. Endocannabinoid signaling in mammalian ovary. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 178, 6–11. [Google Scholar] [CrossRef]
- Correa, F.; Wolfson, M.L.; Valchi, P.; Aisemberg, J.; Franchi, A.M. Endocannabinoid system and pregnancy. Reproduction 2016, 152, R191–R200. [Google Scholar] [CrossRef]
- Pertwee, R.G. Endocannabinoids and their pharmacological actions. In Endocannabinoids. Handbook of Experimental Pharmacology, 1st ed.; Pertwee, R., Ed.; Springer: Cham, Switzerland, 2015; Volume 231, pp. 1–37. [Google Scholar]
- Maccarrone, M. Endocannabinoids: Friends and foes of reproduction. Prog. Lipid Res. 2009, 48, 344–354. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; Di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapino, C.; Battista, N.; Bari, M.; Maccarrone, M. Endocannabinoids as biomarkers of human reproduction. Hum. Reprod. Update 2014, 20, 501–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Forester, E.; Piomelli, D.; Giuffrida, A. N-Acylethanolamines in human reproductive fluids. Chem. Phys. Lipids 2002, 121, 211–227. [Google Scholar] [CrossRef] [Green Version]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. Fluctuation in anandamide levels from ovulation to early pregnancy in in-vitro fertilization-embryo transfer women, and its hormonal regulation. Hum. Reprod. 2009, 24, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Luo, X.T.; Yao, Y.R.; Xiao, H.M.; Guo, M.Q. Investigation of changes in endocannabinoids and N-acylethanolamides in biofluids, and their correlations with female infertility. J. Chromatogr. A 2017, 1509, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Yang, Y.; Xu, Y.; Zhang, J.; Jiang, L.; Hao, G. Decreased expression of fatty acid amide hydrolase in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2017, 33, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Almada, M.; Amaral, C.; Diniz-da-Costa, M.; Correia-da-Silva, G.; Teixeira, N.A.; Fonseca, B.M. The endocannabinoid anandamide impairs in vitro decidualization of human cells. Reproduction 2016, 152, 351–361. [Google Scholar] [CrossRef]
- Bagavandoss, P.; Grimshaw, S. Temporal and spatial distribution of the cannabinoid receptors (CB1, CB2) and fatty acid amide hydroxylase in the rat ovary. Anat. Rec. (Hoboken) 2010, 293, 1425–1432. [Google Scholar] [CrossRef]
- El-Talatini, M.R.; Taylor, A.H.; Elson, J.C.; Brown, L.; Davidson, A.C.; Konje, J.C. Localisation and function of the endocannabinoid system in the human ovary. PLoS ONE 2009, 4, e4579. [Google Scholar] [CrossRef]
- Peralta, L.; Agirregoitia, E.; Mendoza, R.; Expósito, A.; Casis, L.; Matorras, R.; Agirregoitia, N. Expression and localization of cannabinoid receptors in human immature oocytes and unfertilized metaphase-II oocytes. Reprod. Biomed. Online 2011, 23, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Agirregoitia, E.; Ibarra-Lecue, I.; Totorikaguena, L.; Mendoza, R.; Expósito, A.; Matorras, R.; Agirregoitia, N. Dynamics of expression and localization of the cannabinoid system in granulosa cells during oocyte nuclear maturation. Fertil. Steril. 2015, 104, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Agirregoitia, E.; Totorikaguena, L.; Expósito, A.; Mendoza, R.; Matorras, R.; Agirregoitia, N. Dynamic of expression and localization of cannabinoid-degrading enzymes FAAH and MGLL in relation to CB1 during meiotic maturation of human oocytes. Cell Tissue Res. 2016, 365, 393–401. [Google Scholar] [CrossRef] [PubMed]
- López-Cardona, A.P.; Sánchez-Calabuig, M.J.; Beltran-Breña, P.; Agirregoitia, N.; Rizos, D.; Agirregoitia, E.; Gutierrez-Adán, A. Exocannabinoids effect on in vitro bovine oocyte maturation via activation of AKT and ERK1/2. Reproduction 2016, 152, 603–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Cardona, A.P.; Pérez-Cerezales, S.; Fernández-González, R.; Laguna-Barraza, R.; Pericuesta, E.; Agirregoitia, N.; Agirregoitia, E. CB1 cannabinoid receptor drives oocyte maturation and embryo development via PI3K/Akt and MAPK pathways. FASEB J. 2017, 31, 3372–3382. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Totaro, A.; Scipioni, L.; Dufrusine, B.; Stepniewski, T.M.; Selent, J.; Dainese, E. Role of palmitoylation of cysteine 415 in functional coupling CB1 receptor to Gαi2 protein. Biotechnol. Appl. Biochem. 2018, 65, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Horner, K.; Livera, G.; Hinckley, M.; Trinh, K.; Storm, D.; Conti, M. Rodent oocytes express an active adenylyl cyclase required for meiotic arrest. Dev. Biol. 2003, 258, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Vaccari, S.; Horner, K.; Mehlmann, L.M.; Conti, M. Generation of mouse oocytes defective in cAMP synthesis and degradation: Endogenous cyclic AMP is essential for meiotic arrest. Dev. Biol. 2008, 316, 124–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susor, A.; Jansova, D.; Anger, M.; Kubelka, M. Translation in the mammalian oocyte in space and time. Cell Tissue Res. 2016, 363, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Oddi, S.; Dainese, E.; Sandiford, S.; Fezza, F.; Lanuti, M.; Chiurchiù, V.; Maccarrone, M. Effects of palmitoylation of Cys(415) in helix 8 of the CB(1) cannabinoid receptor on membrane localization and signalling. Br. J. Pharmacol. 2012, 165, 2635–2651. [Google Scholar] [CrossRef]
- Hirohashi, N.; Yanagimachi, R. Sperm acrosome reaction: Its site and role in fertilization. Biol. Reprod. 2018, 99, 127–133. [Google Scholar] [CrossRef]
- Yang, Z.M.; Paria, B.C.; Dey, S.K. Activation of brain-type cannabinoid receptors interferes with preimplantation mouse embryo development. Biol. Reprod. 1996, 55, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Paria, B.C.; Dey, S.K. Ligand-receptor signaling with endocannabinoids in preimplantation embryo development and implantation. Chem. Phys. Lipids 2000, 108, 211–220. [Google Scholar] [CrossRef]
- Buckley, N.E.; Hansson, S.; Harta, G.; Mezey, E. Expression of the CB1 and CB2 receptor messenger RNAs during embryonic development in the rat. Neuroscience 1998, 82, 1131–1149. [Google Scholar] [CrossRef]
- Conti, M. Phosphodiesterases and regulation of female reproductive function. Curr. Opin. Pharmacol. 2011, 11, 665–669. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Luciano, A.M.; Richani, D.; Zeng, H.T.; Wang, X.; Vos, M.D.; Thompson, J.G. Oocyte maturation and quality: Role of cyclic nucleotides. Reproduction 2016, 152, R143–R157. [Google Scholar] [CrossRef] [PubMed]
- Richani, D.; Gilchrist, R.B. The epidermal growth factor network: Role in oocyte growth, maturation and developmental competence. Hum. Reprod. Update 2018, 24, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lowther, K.M.; Nikolaev, V.O.; Mehlmann, L.M. Endocytosis in the mouse oocyte and its contribution to cAMP signaling during meiotic arrest. Reproduction 2011, 141, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nader, N.; Dib, M.; Daalis, A.; Kulkarni, R.P.; Machaca, K. Role for endocytosis of a constitutively active GPCR (GPR185) in releasing vertebrate oocyte meiotic arrest. Dev. Biol. 2014, 395, 355–366. [Google Scholar] [CrossRef] [PubMed]
- den Boon, F.S.; Chameau, P.; Schaafsma-Zhao, Q.; van Aken, W.; Bari, M.; Oddi, S.; Werkman, T.R. Excitability of prefrontal cortical pyramidal neurons is modulated by activation of intracellular type-2 cannabinoid receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 3534–3539. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.; Prasad, S.; Shrivastav, T.G.; Chaube, S.K. Calcium signaling during meiotic cell cycle regulation and apoptosis in mammalian oocytes. J. Cell Physiol. 2017, 232, 976–981. [Google Scholar] [CrossRef]
- Willoughby, D.; Cooper, D.M.F. Organization and Ca2+ regulation of adenylyl cyclases in cAMP microdomains. Physiol. Rev. 2007, 87, 965–1010. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P. Role of G-protein-coupled estrogen receptor (GPER/GPR30) in maintenance of meiotic arrest in fish oocytes. J. Steroid Biochem. Mol. Biol. 2017, 167, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Bagavandoss, P.; Grimshaw, S. Distribution of adenylyl cyclases in the rat ovary by immunofluorescence microscopy. Anat. Rec. (Hoboken) 2012, 295, 1717–1726. [Google Scholar] [CrossRef] [PubMed]
- Gaetz, J.; Kapoor, T.M. Dynein/dynactin regulate metaphase spindle length by targeting depolymerizing activities to spindle poles. J. Cell Biol. 2004, 166, 465–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennabi, I.; Terret, M.E.; Verlhac, M.H. Meiotic spindle assembly and chromosome segregation in oocytes. J. Cell Biol. 2016, 215, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Jiao, X.F.; Zhang, J.Y.; Wu, D.; Ding, Z.M.; Wang, Y.S.; Huo, L.J. Nucleoporin35 is a novel microtubule associated protein functioning in oocyte meiotic spindle architecture. Exp. Cell Res. 2018, 371, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Tomari, H.; Honjo, K.; Kunitake, K.; Aramaki, N.; Kuhara, S.; Hidaka, N.; Horiuchi, T. Meiotic spindle size is a strong indicator of human oocyte quality. Reprod. Med. Biol. 2018, 17, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.W.; Zhang, G.L.; Schatten, H.; Carroll, J.; Sun, Q.Y. Cytoplasmic determination of meiotic spindle size revealed by a unique inter-species germinal vesicle transfer model. Sci. Rep. 2016, 6, 19827. [Google Scholar] [CrossRef]
- Radford, S.J.; Nguyen, A.L.; Schindler, K.; McKim, K.S. The chromosomal basis of meiotic acentrosomal spindle assembly and function in oocytes. Chromosoma 2017, 126, 351–364. [Google Scholar] [CrossRef]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.Y.; Lu, H.C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [Green Version]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Mahony, M.C.; Giuffrida, A.; Makriyannis, A. Evidence that anandamide-signaling regulates human sperm functions required for fertilization. Mol. Reprod. Dev. 2002, 63, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Amoako, A.A.; Marczylo, T.H.; Elson, J.; Taylor, A.H.; Willets, J.M.; Konje, J.C. Relationship between seminal plasma levels of anandamide congeners palmitoylethanolamide and oleoylethanolamide and semen quality. Fertil. Steril. 2014, 102, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Rossi, G.; Santilli, A.; Stefano, L.D.; Hoshino, Y.; Sato, E.; Macchiarelli, G. Akt expression in mouse oocytes matured in vivo and in vitro. Reprod. Biomed. Online 2010, 20, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compagnucci, C.; Di Siena, S.; Bustamante, M.B.; Di Giacomo, D.; Di Tommaso, M.; Maccarrone, M.; Sette, C. Type-1 (CB1) cannabinoid receptor promotes neuronal differentiation and maturation of neural stem cells. PLoS ONE 2013, 8, e54271. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; D’Addario, C. Assessing Gene Expression of the Endocannabinoid System. Methods Mol. Biol. 2016, 1412, 237–246. [Google Scholar] [PubMed]
- Pucci, M.; Pasquariello, N.; Battista, N.; Di Tommaso, M.; Rapino, C.; Fezza, F.; Maccarrone, M. Endocannabinoids stimulate human melanogenesis via type-1 cannabinoid receptor. J. Biol. Chem. 2012, 287, 15466–15478. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Palmerini, M.G.; Macchiarelli, G.; Buccione, R.; Cecconi, S. Mancozeb adversely affects meiotic spindle organization and fertilization in mouse oocytes. Reprod. Toxicol. 2006, 22, 51–55. [Google Scholar] [CrossRef]
- Di Nisio, V.; Rossi, G.; Palmerini, M.G.; Macchiarelli, G.; Tiboni, G.M.; Cecconi, S. Increased rounds of gonadotropin stimulation have side effects on mouse fallopian tubes and oocytes. Reproduction 2018, 155, 245–250. [Google Scholar] [CrossRef]
- Coticchio, G.; Rossi, G.; Borini, A.; Grøndahl, C.; Macchiarelli, G.; Flamigni, C.; Cecconi, S. Mouse oocyte meiotic resumption and polar body extrusion in vitro are differentially influenced by FSH, epidermal growth factor and meiosis-activating sterol. Hum. Reprod. 2004, 19, 2913–2918. [Google Scholar] [CrossRef] [Green Version]
- Sanfins, A.; Lee, G.Y.; Plancha, C.E.; Overstrom, E.W.; Albertini, D.F. Distinctions in meiotic spindle structure [1] and assembly during in vitro and in vivo maturation of mouse oocytes. Biol. Reprod. 2003, 69, 2059–2067. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Num. of Oocytes | MI (%) | MII (%) |
|---|---|---|---|
| Ctr | 30 | 96.02 ± 3.98 | 81.25 ± 2.13 |
| SR1 | 45 | 95.80 ± 3.15 | 82.93 ± 1.42 |
| SR2 | 45 | 94.56 ± 5.03 | 80.95 ± 3.26 |
| ML193 | 80 | 95.51 ± 4.36 | 81.16 ± 2.70 |
| SR1+SR2+ML193 | 50 | 94.83 ± 2.91 | 81.60 ± 2.24 |
| Gene | Corresponding Protein | PCR Primers | Annealing T (°C) | Reference |
|---|---|---|---|---|
| Cnr1 | CB1R | Fw: 5′-CCAAGAAAAGATGACGGCAG-3′ Rev: 5′-AGGATGACACATAGCACCAG-3′ | 57 | [55] |
| Cnr2 | CB2R | Fw: 5′-TCGCTTACATCCTTCAGACAG-3′ Rev: 5′-TCTTCCCTCCCAACTCCTTC-3′ | 57 | [55] |
| Gpr55 | GPR55 | Fw: 5′-ATTCGATTCCGTGGATAAGC-3′ Rev: 5′-ATGCTGATGAAGTAGAGGC-3′ | 57 | [56] |
| Trpv1 | TRPV1 | Fw: 5′-TGAACTGGACTACCTGGAAC-3′ Rev: 5′-TCCTTGAAGACCTCAGCATC-3′ | 57 | [56] |
| Actb | Actin | Fw: 5′-CTGTCGAGTCGCGTCCACCC-3′ Rev: 5′-GCTTTGCACATGCCGGAGCC-3′ | 57 | [55] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cecconi, S.; Rossi, G.; Oddi, S.; Di Nisio, V.; Maccarrone, M. Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation. Int. J. Mol. Sci. 2019, 20, 2866. https://doi.org/10.3390/ijms20122866
Cecconi S, Rossi G, Oddi S, Di Nisio V, Maccarrone M. Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation. International Journal of Molecular Sciences. 2019; 20(12):2866. https://doi.org/10.3390/ijms20122866
Chicago/Turabian StyleCecconi, Sandra, Gianna Rossi, Sergio Oddi, Valentina Di Nisio, and Mauro Maccarrone. 2019. "Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation" International Journal of Molecular Sciences 20, no. 12: 2866. https://doi.org/10.3390/ijms20122866
APA StyleCecconi, S., Rossi, G., Oddi, S., Di Nisio, V., & Maccarrone, M. (2019). Role of Major Endocannabinoid-Binding Receptors during Mouse Oocyte Maturation. International Journal of Molecular Sciences, 20(12), 2866. https://doi.org/10.3390/ijms20122866