The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1


Abstract
:1. Introduction
2. Results
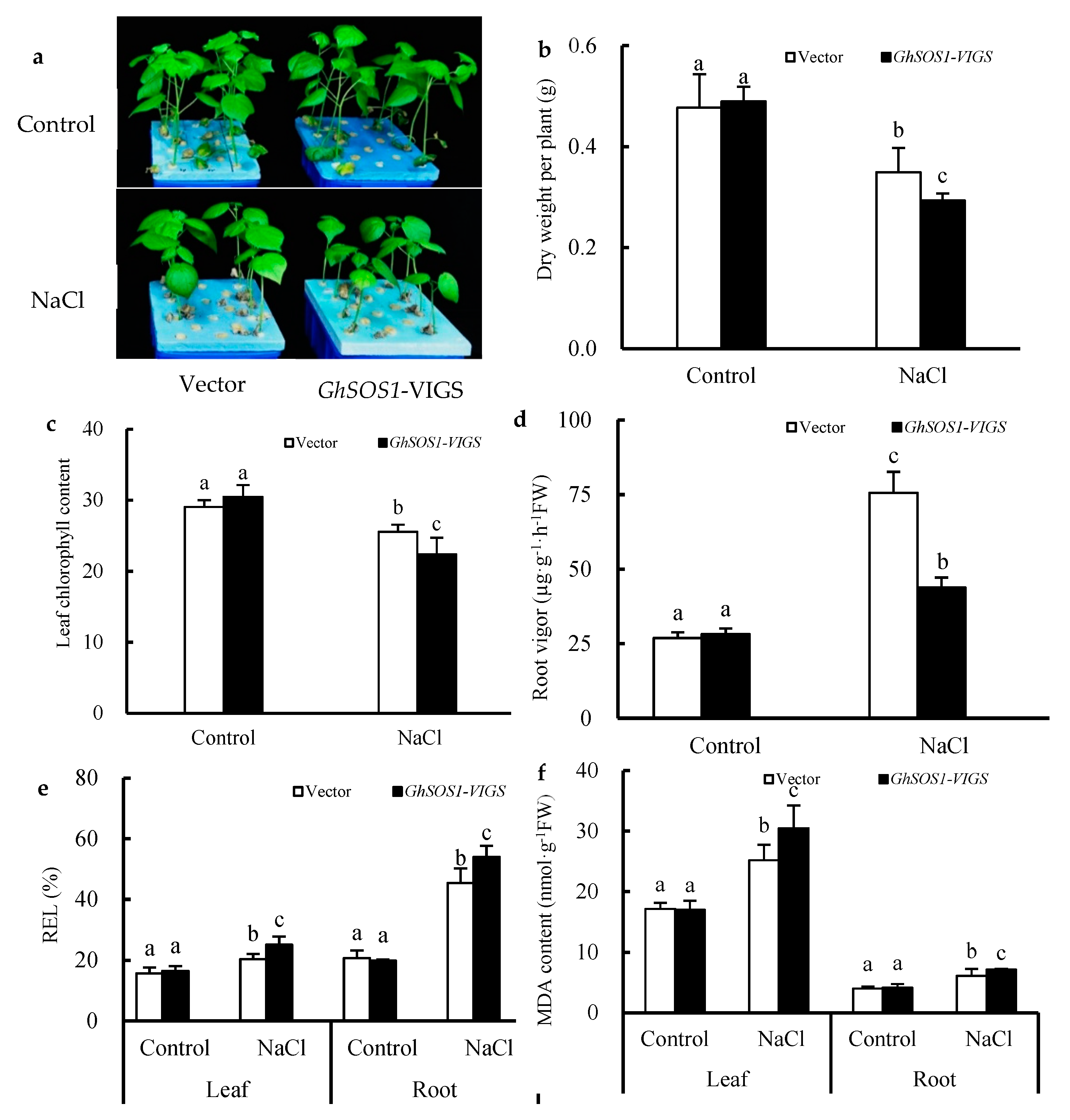
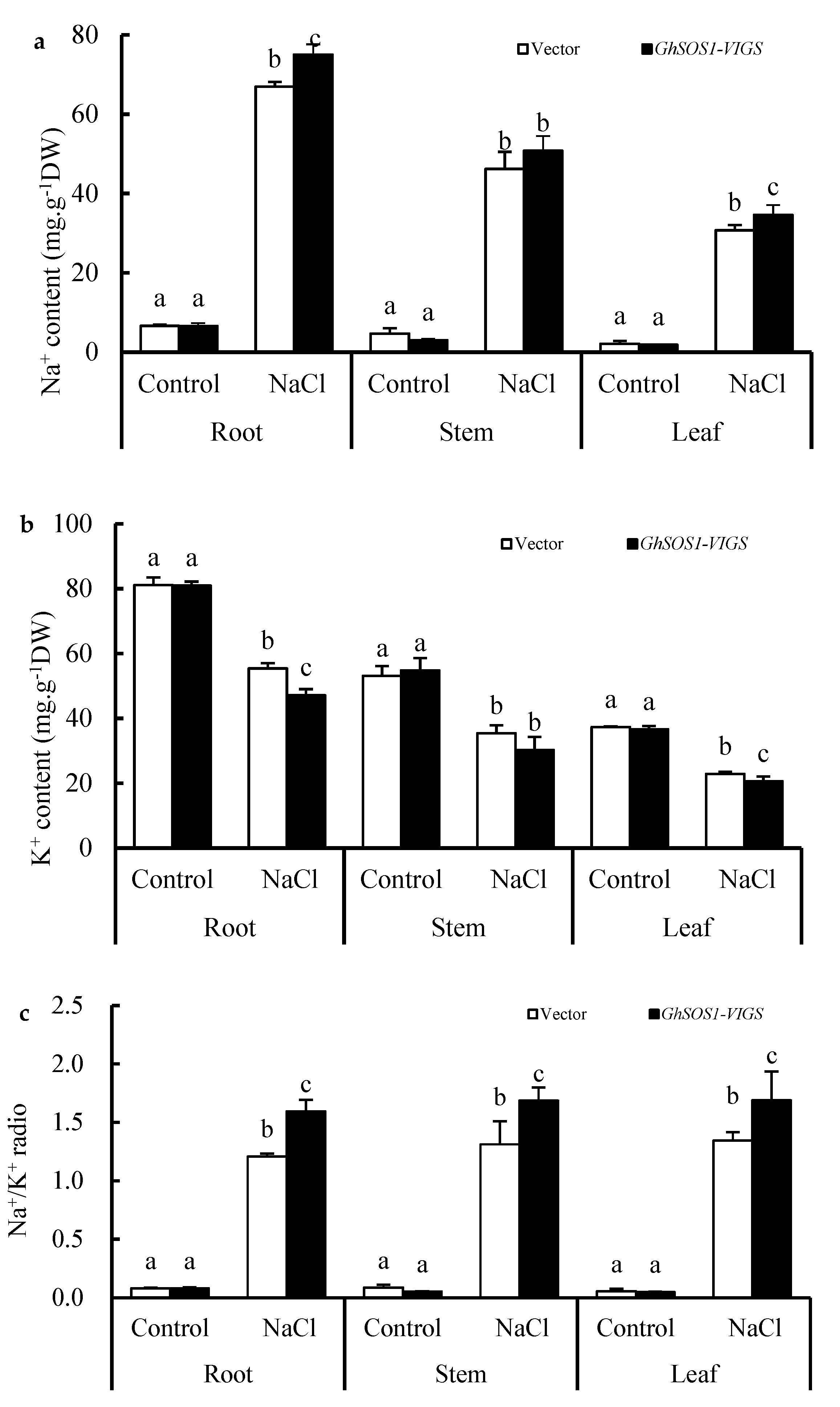
2.1. Effects of Salt Stress on the Growth and Related Physiological Parameters of GhSOS1-VIGS Cotton Seedlings
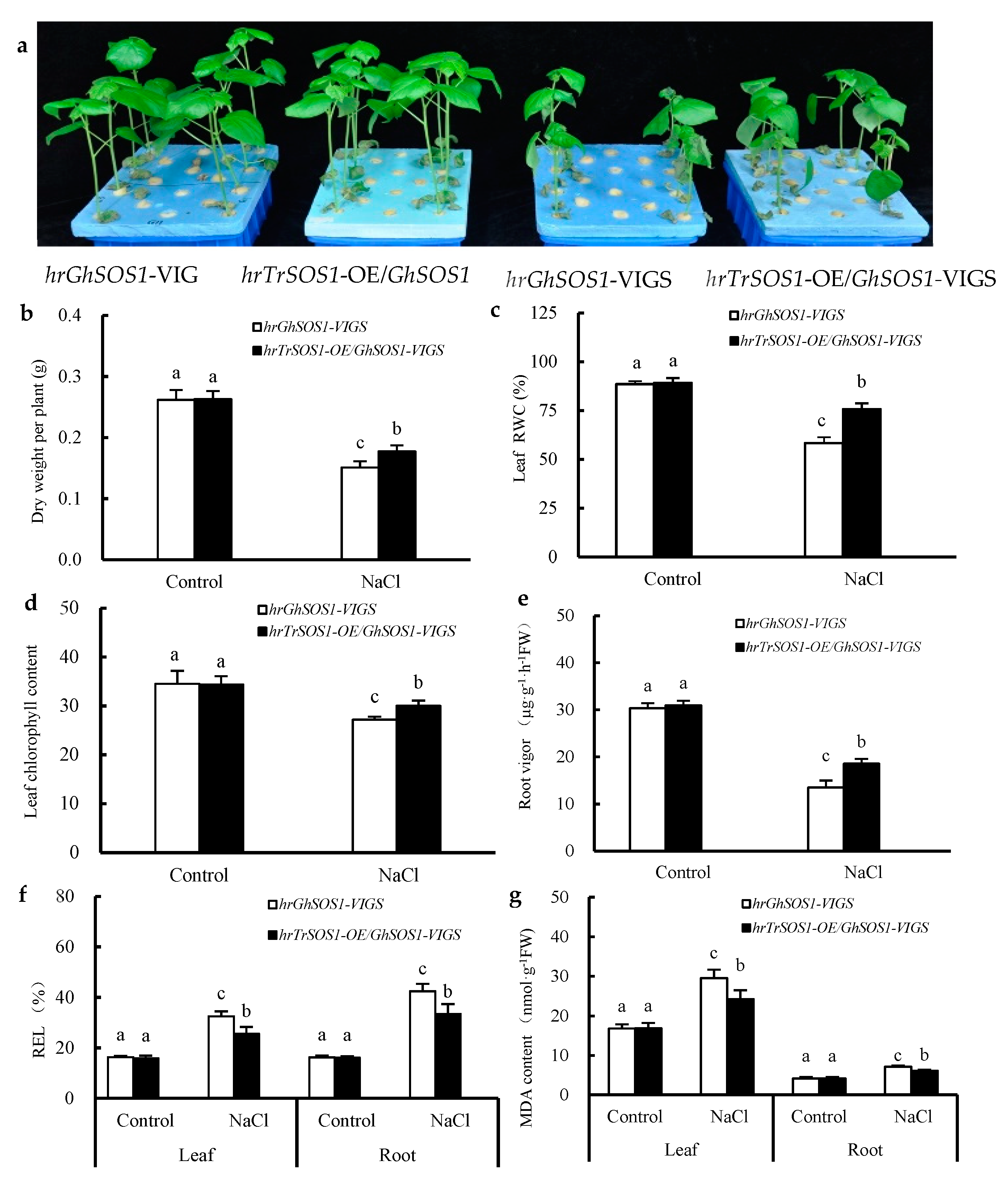
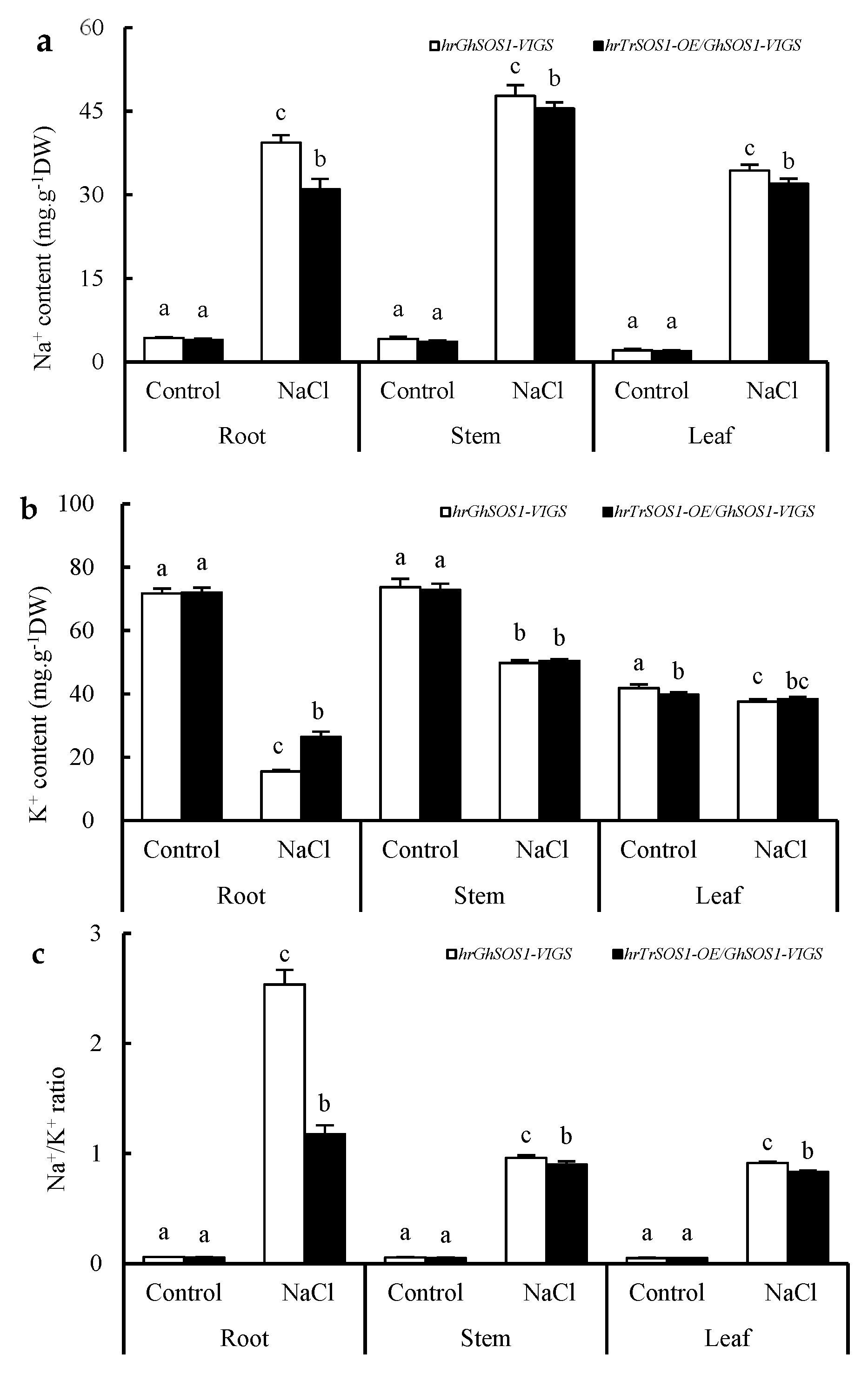
2.2. Effects of Salt Stress on the Growth and Related Physiological Parameters of TrSOS1-Transgenic Hairy Root Composite Cotton Plants Exhibiting VIGS of GhSOS1
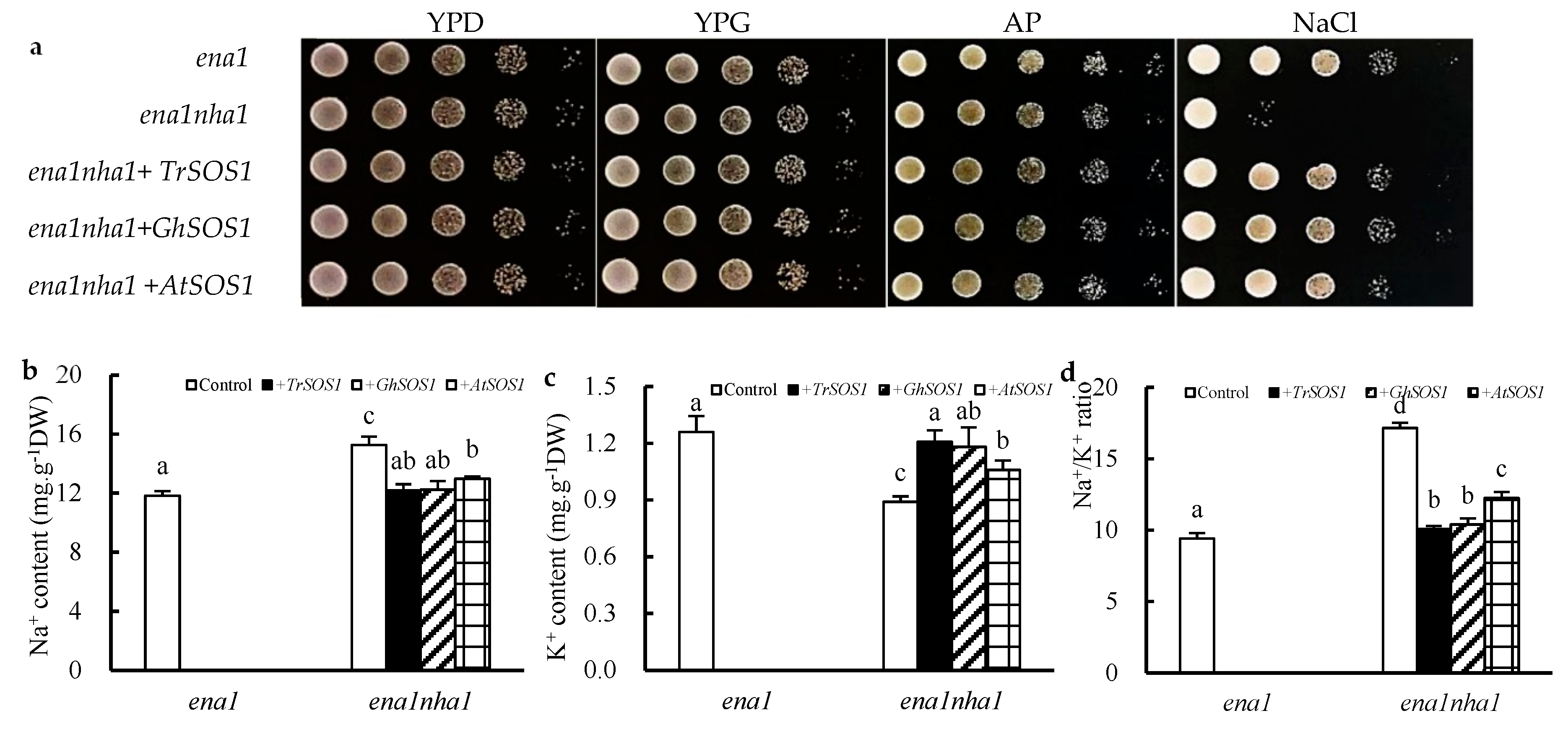
2.3. Comparison of TrSOS1, GhSOS1, and AtSOS1 complementation in the yeast NHA1 mutant
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Bacteria and Yeast Strains, and Plasmids
4.2. TrSOS1, GhSOS1, and AtSOS1 Gene Cloning and Vector Construction
4.3. Construction of GhSOS1-VIGS Cotton Plants
4.4. Construction of hrTrSOS1-OE/GhSOS1-VIGS Cotton Plants
4.5. Semi-Quantitative RT-PCR Analysis
4.6. Salt-Tolerance Tests of GhSOS1-VIGS or TrSOS1-OE/GhSOS1-VIGS Cotton Plants
4.7. Complementation Test of TrSOS1, GhSOS1, and AtSOS1 in Yeast Mutant
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Wan, Q.; He, X.L.; Ning, L.H.; Huang, Y.H.; Xu, Z.L.; Liu, J.; Shao, H.B. Genome-wide characterization of the ankyrin repeats gene family under salt stress in soybean. Sci. Total Environ. 2016, 568, 899–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Choi, J.; An, G.; Kim, S.R. Ectopic expression of OsSta2 enhances salt stress tolerance in rice. Front. Plant Sci. 2017, 8, 316. [Google Scholar] [CrossRef] [PubMed]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef]
- Adem, G.D.; Roy, S.J.; Zhou, M.X.; Bowman, J.P.; Shabala, S. Evaluating contribution of ionic, osmotic and oxidative stress components towards salinity tolerance in barley. BMC Plant Biol. 2014, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.N.; Zhou, Q.; Yu, B.J. Effects of Zn2+ and niflumic acid on photosynthesis in Glycine soja and Glycine max seedlings under NaCl stress. Environ. Exp. Bot. 2009, 65, 304–309. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.Z.; Ishitani, M.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef]
- Yu, B.J.; Liu, Y.L. SOS genes family and plant salt tolerance. Plant Physiol. Commun. 2004, 4, 409–413. [Google Scholar]
- Bartels, D.; Dinakar, C. Balancing salinity stress responses in halophytes and non-halophytes: A comparison between Thellungiella and Arabidopsis thaliana. Funct. Plant Biol. 2013, 40, 819–831. [Google Scholar] [CrossRef]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, I.; Iqbal, M.; Hakvoort, H.W.J.; Bliek, M.; de Boer, B.; Schat, H. Expression levels and promoter activities of candidate salt tolerance genes in halophyte and glycophytic Brassicaceae. Environ. Exp. Bot. 2014, 99, 59–66. [Google Scholar] [CrossRef]
- Razzaque, S.; Elias, S.M.; Biswas, S.; Haque, T.; Seraj, Z.I. Cloning of the plasma membrane sodium/hydrogen antiporter SOS1 for its over expression in rice. Plant Tissue Cult. Biotechnol. 2013, 23, 263–273. [Google Scholar] [CrossRef]
- Olías, R.; Eljakaoui, Z.; Li, J.; De Morales, P.A.; Marı’n-Manzano, M.C.; Pardo, J.M.; Belver, A. The plasma membrane Na+/H+ antiporter SOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+ between plant organs. Plant Cell Environ. 2009, 32, 904–916. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.J.; Sun, J.; Cao, P.P.; Ren, L.P.; Liu, C.; Chen, S.M.; Chen, F.D.; Jiang, J.F. Variation in tissue Na+ content and the activity of SOS1 genes among two species and two related genera of Chrysanthemum. BMC Plant Biol. 2016, 16, 98. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.G.; Lu, X.K.; Shu, N.; Wang, D.L.; Wang, S.; Wang, J.J.; Guo, L.X.; Guo, X.N.; Fan, W.L.; Lin, Z.X.; et al. GhSOS1, a plasma membrane Na+/H+ antiporter gene from upland cotton, enhances salt tolerance in transgenic Arabidopsis thaliana. PLoS ONE 2017, 12, e0181450. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.X.; Xu, L.; Yu, B.J. A putative soybean GmsSOS1 confers enhanced salt tolerance to transgenic Arabidopsis sos1-1 mutant. Protoplasma 2015, 252, 127–134. [Google Scholar] [CrossRef]
- Zhao, X.F.; Wei, P.P.; Liu, Z.; Yu, B.J.; Shi, H.Z. Soybean Na+/H+ antiporter GmsSOS1 enhances antioxidant enzyme activity and reduces Na+ accumulation in Arabidopsis and yeast cells under salt stress. Acta Physiol. Plant. 2017, 39, 19. [Google Scholar] [CrossRef]
- Oh, D.H.; Leidi, E.; Zhang, Q.; Hwang, S.M.; Li, Y.; Quintero, F.J.; Jiang, X.Y.; D’Urzo, M.P.; Lee, S.Y.; Zhao, Y.X.; et al. Loss of halophytism by interference with SOS1 expression. Plant Physiol. 2009, 151, 210–222. [Google Scholar] [CrossRef]
- Maughan, P.J.; Turner, T.B.; Coleman, C.E.; Elzinga, D.B.; Jellen, E.N.; Morales, J.A.; Udall, J.A.; Fairbanks, D.J.; Bonifacio, A. Characterization of Salt Overly Sensitive 1 (SOS1) gene homologous in quinoa (Chenopodium quinoa Willd.). Genome 2009, 52, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.S.; Shukla, P.S.; Jha, A.; Agarwal, P.K.; Jha, B. The SbSOS1 gene from the extreme halophyte Salicornia brachiata enhances Na+ loading in xylem and confers salt tolerance in transgenic tobacco. BMC Plant Biol. 2012, 12, 188. [Google Scholar] [CrossRef]
- Zhou, Y.; Yin, X.C.; Duan, R.J.; Hu, Y.P.; Xie, Q.; Li, R.M.; Zhu, B.B.; Fu, S.P.; Guo, J.C.; Jiang, X.Y. The Sesuvium portulacastrum Plasma Membrane Na+/H+ Antiporter SpSOS1 Complemented the Salt Sensitivity of Transgenic Arabidopsis sos1 Mutant Plants. Plant Mol. Biol. Rep. 2018, 36, 553–563. [Google Scholar] [CrossRef]
- Abid, M.A.; Liang, C.; Malik, W.; Meng, Z.G.; Tao, Z.; Meng, Z.G.; Ashraf, J.; Guo, S.D.; Zhang, R. Cascades of ionic and molecular networks involved in expression of genes underpin salinity tolerance in cotton. J. Plant Growth Regul. 2018, 37, 668–679. [Google Scholar] [CrossRef]
- Peng, Z.; He, S.P.; Sun, J.L.; Pan, Z.E.; Gong, W.F.; Lu, Y.L.; Du, X.M. Na+ compartmentalization related to salinity stress tolerance in upland cotton (Gossypium hirsutum) seedlings. Sci. Rep. 2016, 6, 34548. [Google Scholar] [CrossRef] [PubMed]
- Leidi, E.O.; Saiz, J. Is salinity tolerance related to Na accumulation in upland cotton (Gossypium hirsutum) seedlings? Plant and Soil 1997, 190, 67–75. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, G.Z.; Du, L.; Shang, X.G.; Cheng, C.Z.; Yang, B.; Hu, Y.; Cai, C.P.; Guo, W.Z. Genetic regulation of salt stress tolerance revealed by RNA-Seq in cotton diploid wild species, Gossypium davidsonii. Sci. Rep. 2016, 6, 20682. [Google Scholar] [CrossRef]
- Wang, N.; Qiao, W.Q.; Liu, X.H.; Shi, J.B.; Xu, Q.H.; Zhou, H.; Yan, G.T.; Huang, Q. Relative contribution of Na+/H+, homeostasis, photochemical efficiency and antioxidant defense system to differential salt tolerance in cotton (Gossypium hirsutum L.) cultivars. Plant Physiol. Biochem. 2017, 119, 121–131. [Google Scholar] [CrossRef]
- Liu, Y.M.; Cheng, C.; Jiang, L.; Yu, B.J. Comparisons on growth, salt ion distribution, and relative expression of SOS1 gene in three species of Tamarix Linn. under NaCl stress. J. Plant Resour. Environ. 2019, 28, 1–9. [Google Scholar]
- Cao, D.; Hou, W.S.; Song, S.K.; Sun, H.B.; Wu, C.X.; Gao, Y.S.; Han, T.F. Assessment of conditions affecting Agrobacterium rhizogenes-mediated transformation of soybean. Plant Cell Tissue Organ Cult. 2009, 96, 45–52. [Google Scholar] [CrossRef]
- Wei, P.P.; Wang, L.C.; Liu, A.L.; Yu, B.J.; Lam, H.M. GmCLC1 confers enhanced salt tolerance through regulating chloride accumulation in soybean. Front. Plant Sci. 2016, 7, 1082. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Schiff, M.; Liu, Y.L.; Dinesh-Kumar, S.P. Efficient virus-induced gene silencing in Arabidopsis. Plant Physiol. 2006, 142, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J.K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Feki, K.; Quintero, F.J.; Khoudi, H.; Leidi, E.O.; Masmoudi, K.; Pardo, J.M.; Brini, F. A constitutively active form of a durum wheat Na+/H+ antiporter SOS1 confers high salt tolerance to transgenic Arabidopsis. Plant Cell Rep. 2014, 33, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.S.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J.K. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Glazebrook, J. Transformation of Agrobacterium Using the Freeze-Thaw Method. Cold Spring Harb Protoc. 2006, 7, 1031–1036. [Google Scholar] [CrossRef]
- Gietz, R.D. Yeast Transformation by the LiAc/SS Carrier DNA/PEG Method. Methods Mol. Biol. 2014, 1163, 33–44. [Google Scholar] [CrossRef]
- Gao, X.Q.; Wheeler, T.; Li, Z.H.; Kenerley, C.M.; He, P.; Shan, L.B. Silencing GhNDR1 and GhMKK2 compromises cotton resistance to Verticillium wilt. Plant J. 2011, 66, 293–305. [Google Scholar] [CrossRef]
- An, J.; Hu, Z.M.; Che, B.N.; Chen, H.Y.; Yu, B.J.; Cai, W.M. Heterologous expression of Panax ginseng PgTIP1 confers enhanced salt tolerance of soybean cotyledon hairy roots, composite, and whole plants. Front. Plant Sci. 2017, 8, 1232. [Google Scholar] [CrossRef]
- Tian, F.; Jia, T.J.; Yu, B.J. Physiological regulation of seed soaking with soybean isoflavones on drought tolerance of Glycine max and Glycine soja. Plant Growth Regul. 2014, 74, 229–237. [Google Scholar] [CrossRef]
- Jouve, L.; Jacques, D.; Douglas, G.C.; Hoffmanna, L.; Hausman, J.F. Biochemical characterization of early and late bud flushing in common ash (Fraxinus excelsior L.). Plant Sci. 2007, 172, 962–969. [Google Scholar] [CrossRef]
- Chen, X.Q.; Yu, B.J. Ionic effects of Na+ and Cl− on photosynthesis in Glycine max seedlings under isoosmotic salt stress. J. Plant Physiol. Mol. Biol. 2007, 33, 294–300. [Google Scholar]
- Li, Y.; Takano, T.; Liu, S.K. Discovery and Characterization of Two Novel Salt-Tolerance Genes in Puccinellia tenuiflora. Int. J. Mol. Sci. 2014, 15, 16469–16483. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purposes | Genes | Primer Sequences (5′ to 3′) | |
|---|---|---|---|
| Plant VIGS | GhCLA1 | F | CTCCATGGGGATCCGGTACCCACAACATCGATGATTTAG |
| R | AGACGCGTGAGCTCGGTACCATGATGAGTAGATTGCAC | ||
| GhSOS1 | F | CTCCATGGGGATCCGGTACCTATTAGCTGTTTTTCTACCCGC | |
| R | AGACGCGTGAGCTCGGTACCATAAATCCAAGCCACAAAACAG | ||
| Plant expression | TrSOS1 | F | GCTTCTGCAGGGGCCCGGGATGGCAGCGGTGTCCGAATT |
| R | TTTAAATGTCGACCCCGGGTCAAGAAGCATGACGGAAAGATAGC | ||
| Semi-quantitative RT-PCR | GhCLA1 | F | TCTTACCCTCACAAAATCTTGAC |
| R | GCATGGATGGCAACAATATT | ||
| GhSOS1 | F | ATCCAGGCAGCATACTGGGA | |
| R | ATGAAGTTGCCGTCGTGCTA | ||
| Ghactin9 | F | TGAAATATCCCATTGAGCACG | |
| R | TGTAGTTTCATGGATTCCAGCAG | ||
| Yeast complementation | TrSOS1 | F | CCGCCAGTGTGCTGGAATTCATGGCAGCGGTGTCCGAATT |
| R | GATGGATATCTGCAGAATTCTCAAGAAGCATGACGGAAAGATAGC | ||
| GhSOS1 | F | CCGCCAGTGTGCTGGAATTCATGGAGGAAGTGAAAGAGTATC | |
| R | GATGGATATCTGCAGAATTCTTAAGAAGCCTGGTGGAAT | ||
| AtSOS1 | F | CCGCCAGTGTGCTGGAATTCATGACGACTGTAATCGACGC | |
| R | GATGGATATCTGCAGAATTCTCATAGATCGTTCCTGAAAACG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che, B.; Cheng, C.; Fang, J.; Liu, Y.; Jiang, L.; Yu, B. The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1. Int. J. Mol. Sci. 2019, 20, 2930. https://doi.org/10.3390/ijms20122930
Che B, Cheng C, Fang J, Liu Y, Jiang L, Yu B. The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1. International Journal of Molecular Sciences. 2019; 20(12):2930. https://doi.org/10.3390/ijms20122930
Chicago/Turabian StyleChe, Benning, Cong Cheng, Jiajia Fang, Yongmei Liu, Li Jiang, and Bingjun Yu. 2019. "The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1" International Journal of Molecular Sciences 20, no. 12: 2930. https://doi.org/10.3390/ijms20122930
APA StyleChe, B., Cheng, C., Fang, J., Liu, Y., Jiang, L., & Yu, B. (2019). The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1. International Journal of Molecular Sciences, 20(12), 2930. https://doi.org/10.3390/ijms20122930