Determination of the MiRNAs Related to Bean Pyralid Larvae Resistance in Soybean Using Small RNA and Transcriptome Sequencing

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Small RNA Sequencing Data
2.2. Identification and Conservation Analyses of the MiRNAs
2.3. Analyses of the Differentially Expressed MiRNAs
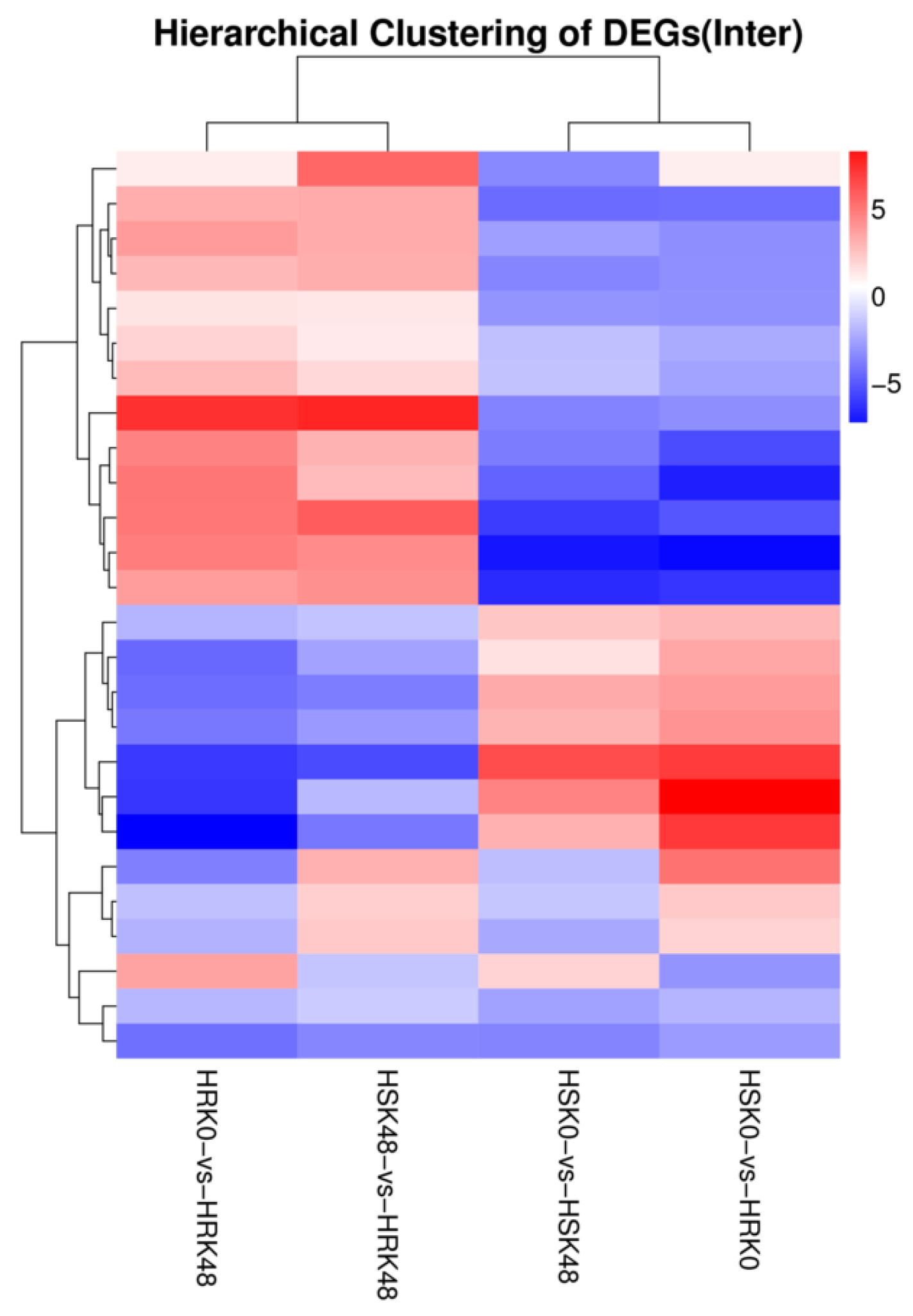
2.4. Cluster Analyses of the Differentially Expressed MiRNAs
2.5. Analysis of the qRT-PCR of the Differential MiRNAs
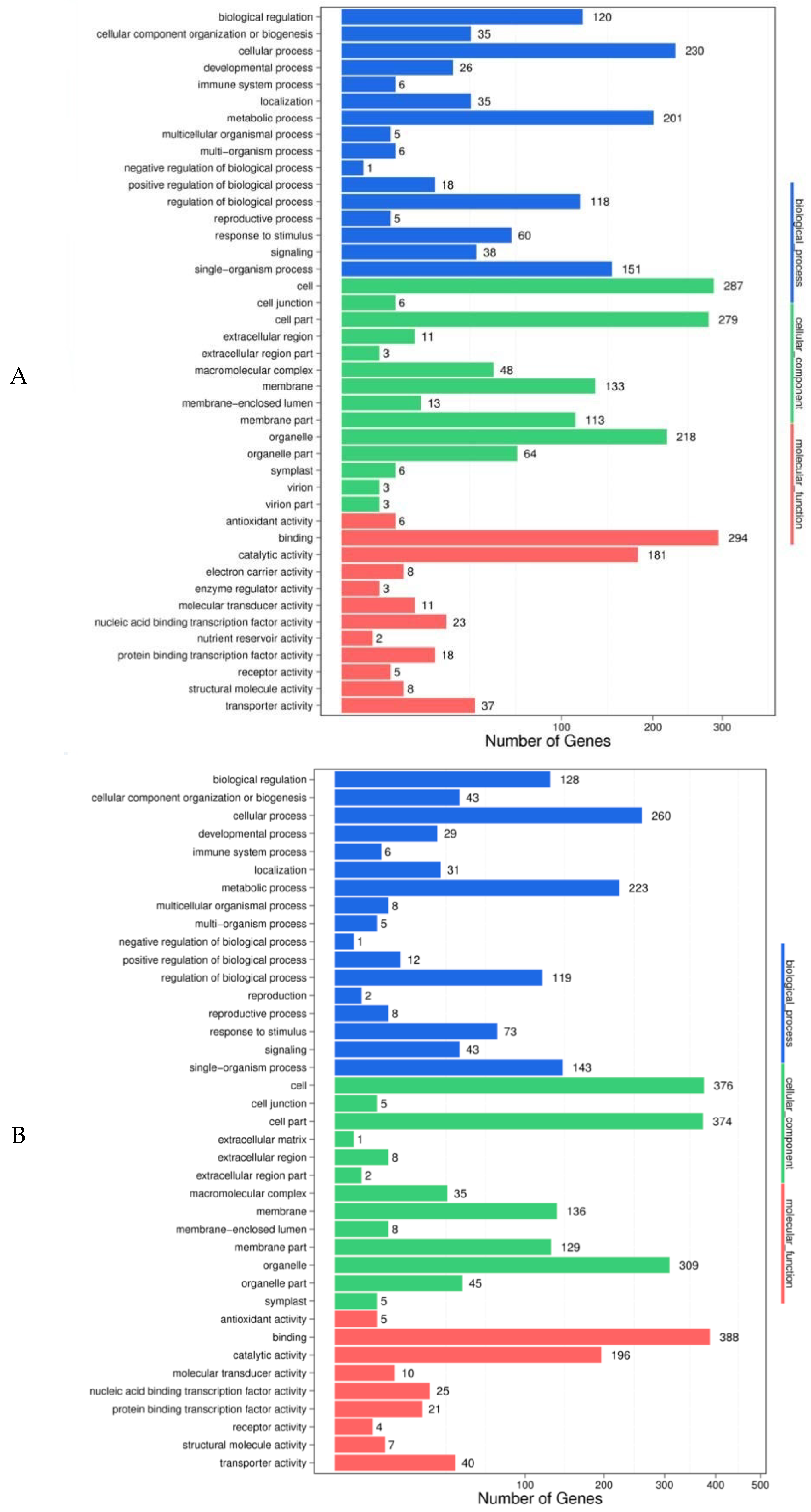
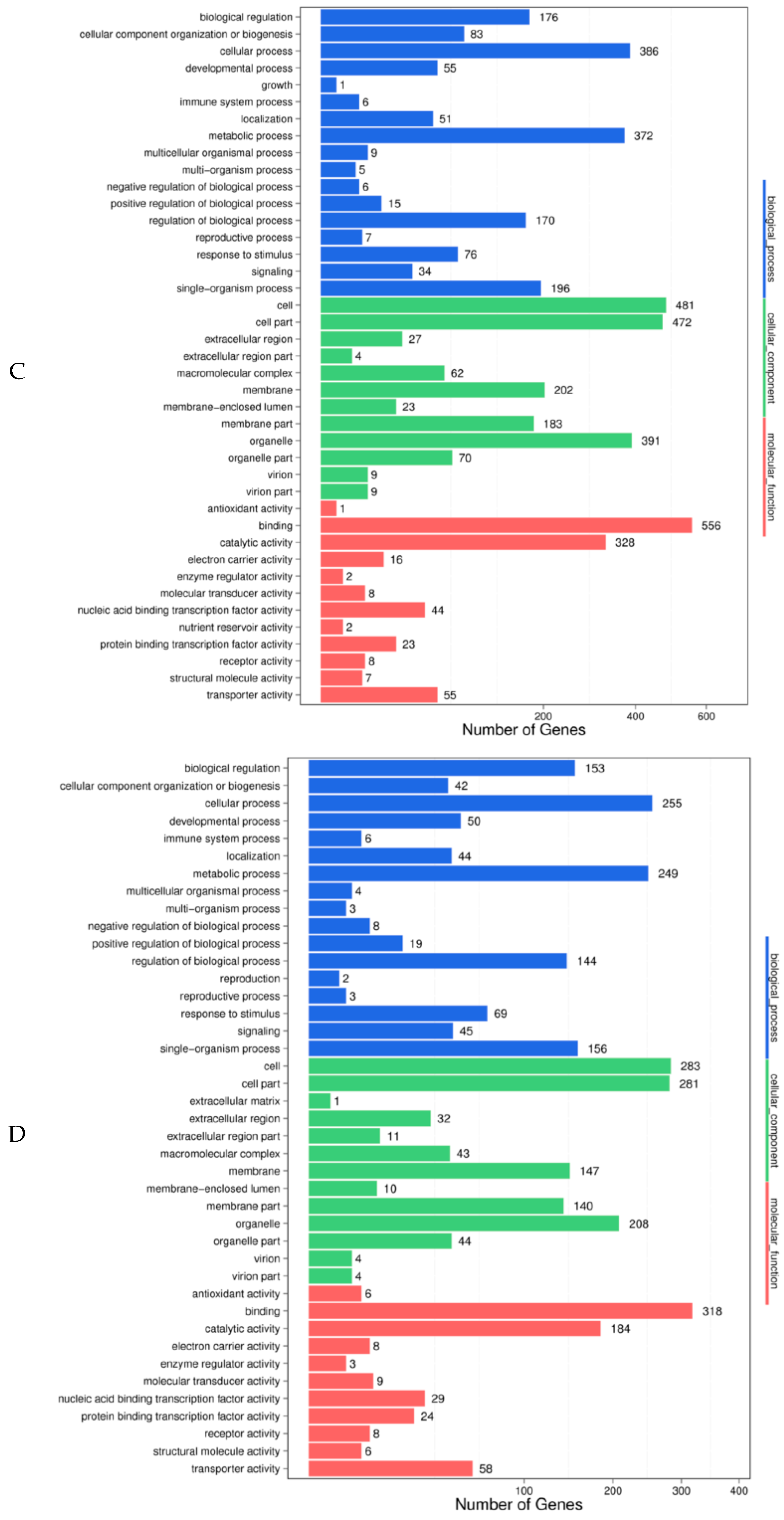
2.6. Target Gene Prediction and the Functional Analyses of the Differentially Expressed MiRNAs
2.7. Conjoint Analysis of the Negative Regulations of the MiRNA/mRNA under Bean Pyralid Larvae Stress
3. Discussion
4. Methods
4.1. Materials, Sample Collection, and Total RNA Extraction
4.2. Library Construction
4.3. Data Analysis of the Small RNA
4.4. Small RNA Predictions and Small RNA Expression Quantifications
4.5. Target Gene Predictions
4.6. Screening of the Differentially Expressed MiRNA
4.7. Hierarchical Cluster Analyses
4.8. Bioinformatics Analyses
4.9. Quantitative Real Time-PCR (qRT-PCR) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| qRT-PCR | Quantitative real time-PCR |
| miRNA | MicroRNA |
| sRNA | Small RNA |
| SPL | Squamosa promoter-binding protein-like |
| HD-Zip | Homodomain-leucine zipper |
| JA | Jasmonic acid |
| GRF | Growth regulating factor |
| MFE | Minimum folding free energy |
| MFEI | Minimum folding free energy index |
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Sunkar, R. MicroRNAs with macro-effects on plant stress responses. Semin. Cell Dev. Biol. 2010, 21, 805–811. [Google Scholar] [CrossRef]
- Meng, Y.J.; Shao, C.G.; Chen, M. Toward microRNA-mediated gene regulatory networks in plants. Brief. Bioinform. 2011, 12, 645–659. [Google Scholar] [CrossRef]
- Sunkar, R.; Li, Y.F.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Trends Plant Sci. 2012, 17, 196–203. [Google Scholar] [CrossRef]
- Zhang, B.H.; Wang, Q.L. MicroRNA-based biotechnology for plant improvement. J. Cell. Physiol. 2015, 230, 1–15. [Google Scholar] [CrossRef]
- Guo, N.; Ye, W.W.; Wu, X.L.; Shen, D.Y.; Wang, Y.C.; Xing, H.; Dou, D.L. Microarray profiling reveals microRNAs involving soybean resistance to Phytophthora sojae. Genome 2011, 54, 954–958. [Google Scholar] [CrossRef]
- Wong, J.; Gao, L.; Yang, Y.; Zhai, J.X.; Arikit, S.; Yu, Y.; Duan, S.Y.; Chan, V.; Xiong, Q.; Yan, J.; et al. Roles of small RNAs in soybean defense against Phytophthora sojae infection. Plant J. 2014, 79, 928–940. [Google Scholar] [CrossRef]
- Yin, X.C.; Wang, J.; Cheng, H.; Wang, X.L.; Yu, D.Y. Detection and evolutionary analysis of soybean miRNAs responsive to soybean mosaic virus. Planta 2013, 237, 1213–1225. [Google Scholar] [CrossRef]
- Li, X.Y.; Wang, X.; Zhang, S.P.; Liu, D.W.; Duan, Y.X.; Dong, W. Identification of soybean microRNAs involved in soybean cyst nematode infection by deep sequencing. PLoS ONE 2012, 7, e39650. [Google Scholar] [CrossRef]
- Xu, M.Y.; Li, Y.H.; Zhang, Q.X.; Xu, T.; Qiu, L.J.; Fan, Y.L.; Wang, L. Novel miRNA and phasiRNA biogenesis networks in soybean roots from two sister lines that are resistant and susceptible to SCN race 4. PLoS ONE 2014, 9, e110051. [Google Scholar] [CrossRef]
- Xu, F.; Liu, Q.; Chen, L.Y.; Kuang, J.B.; Walk, T.; Wang, J.X.; Liao, H. Genome-wide identification of soybean microRNAs and their targets reveals their organ-specificity and responses to phosphate starvation. BMC Genom. 2013, 14, 66. [Google Scholar] [CrossRef]
- Sha, A.H.; Chen, Y.H.; Ba, H.P.; Shan, Z.H.; Wu, X.J.; Qiu, D.Z.; Chen, S.L.; Zhou, X.N. Identification of Glycine max microRNAs in response to phosphorus deficiency. J. Plant Biol. 2012, 55, 268–280. [Google Scholar] [CrossRef]
- Fang, X.L.; Zhao, Y.Y.; Ma, Q.B.; Huang, Y.; Wang, P.; Zhang, J.; Nian, H.; Yang, C.Y. Identification and comparative analysis of cadmium tolerance-associated miRNAs and their targets in two soybean genotypes. PLoS ONE 2013, 8, e81471. [Google Scholar] [CrossRef]
- Zeng, Q.Y.; Yang, C.Y.; Ma, Q.B.; Li, X.P.; Dong, W.W.; Nian, H. Identification of wild soybean miRNAs and their target genes responsive to aluminum stress. BMC Plant Biol. 2012, 12, 182. [Google Scholar] [CrossRef]
- Choi, K.H.; Hong, Y.K.; Chang, Y.J.; Moon, J.S.; Kim, C.S.; Choi, D.C.; Kim, T.H. Development under constants temperatures and seasonal prevalence in soybean field of the bean pyralid, Omiodes indicates (Lepidoptera: Crambidae). Korean J. Appl. Entomol. 2008, 47, 353–358. [Google Scholar] [CrossRef]
- Editorial Committee of Plate of Chinese Diseases and Insects on Crop. Plate of Chinese Diseases and Insects on Crop, Fifth Fascicule, Diseases and Insectson Oil Crop (First); Agricultural Press: Beijing, China, 1982; pp. 136–137. [Google Scholar]
- Sun, Z.D.; Yang, S.Z.; Chen, H.Z.; Li, C.Y.; Long, L.P. Identification of soybean resistance to bean pyralid (Lamprosema indicata Fabricicus) and oviposition preference of bean pyralid on soybean varieties. Chin. J. Oil Crop Sci. 2005, 27, 69–71. [Google Scholar]
- Long, L.P.; Yang, S.Z.; Chen, H.Z.; Qin, J.L.; Li, C.Y.; Sun, Z.D. Effects of different genotypes of soybean varieties on the experimental population of bean pyralid (Lamprosema indicata Fabricicus). Oil Crop Sci. 2004, 26, 67–70. [Google Scholar]
- Ding, X.L. Identification and Functional Study of Differential miRNA in Cytoplasmic-Nuclear Male Sterile Line NJCMS1A and Its Maintainer, Restorer of Soybean. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2017. [Google Scholar]
- Zhang, B.H.; Pan, X.P.; Cannon, C.H.; Cobb, G.P.; Anderson, T.A. Conservation and divergence of plant microRNA genes. Plant J. 2006, 46, 243–259. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Yang, X.Y.; Wang, L.C.; Yuan, D.J.; Lindsey, K.; Zhang, X.L. Small RNA and degradome sequencing reveal complex miRNA regulation during cotton somatic embryogenesis. J. Exp. Bot. 2013, 64, 1521–1536. [Google Scholar] [CrossRef]
- Zeng, W.Y.; Sun, Z.D.; Cai, Z.Y.; Chen, H.Z.; Lai, Z.G.; Yang, S.Z.; Tang, X.M. Comparative transcriptome analysis of soybean response to bean pyralid larvae. BMC Genom. 2017, 18, 871. [Google Scholar] [CrossRef]
- Ruiz-Ferrer, V.; Voinnet, O. Roles of plant small RNAs in biotic stress responses. Annu. Rev. Plant Biol. 2009, 60, 485–510. [Google Scholar] [CrossRef]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in arabidopsis. Plant Mol. Biol. 2008, 67, 183–195. [Google Scholar] [CrossRef]
- Chuck, G.; Cigan, A.M.; Saeteurn, K.; Hake, S. The heterochronic maize mutant Corngrass1 results from overexpression of a tandem microRNA. Nat. Genet. 2007, 39, 544–553. [Google Scholar] [CrossRef]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weiqel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef]
- Ge, Y.F.; Han, J.Y.; Zhou, G.X.; Xu, Y.M.; Ding, Y.; Shi, M.; Guo, C.K.; Wu, G. Silencing of miR156 confers enhanced resistance to brown planthopper in rice. Planta 2018, 248, 813–826. [Google Scholar] [CrossRef]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.R.; Bäurle, I. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef]
- Chuck, G.; Candela, H.; Hake, S. Big impacts by small RNAs in plant development. Curr. Opin. Plant Biol. 2009, 12, 81–86. [Google Scholar] [CrossRef]
- Liu, Q.L.; Yao, X.Z.; Pi, L.M.; Wang, H.; Cui, X.F.; Huang, H. The ARGONAUTE10 gene modulates shoot apical meristem maintenance and establishment of leaf polarity by repressing miR165/166 in arabidopsis. Plant J. 2009, 58, 27–40. [Google Scholar] [CrossRef]
- Wang, H.; Li, G.B.; Zhang, D.Y.; Lin, J.; Sheng, B.L.; Han, J.L.; Chang, Y.H. Biological functions of HD-Zip transcription factors. Hereditas 2013, 35, 1179–1188. [Google Scholar] [CrossRef]
- Ding, D.; Zhang, L.F.; Wang, H.; Liu, Z.J.; Zhang, Z.X.; Zheng, Y.L. Differential expression of miRNAs in response to salt stress in maize roots. Ann. Bot. 2009, 103, 29–38. [Google Scholar] [CrossRef]
- Kitazumi, A.; Kawahara, Y.; Onda, T.S.; De Koeyer, D.; de los Reyes, B.G. Implications of miR166 and miR159 induction to the basal response mechanisms of an andigena potato (Solanum tuberosum subsp. andigena) to salinity stress, predicted from network models in arabidopsis. Genome 2015, 58, 13–24. [Google Scholar] [CrossRef]
- Lv, D.K.; Bai, X.; Li, Y.; Ding, X.D.; Ge, Y.; Cai, H.; Ji, W.; Wu, N.; Zhu, Y.M. Profiling of cold-stress-responsive miRNAs in rice by microarrays. Gene 2010, 459, 39–47. [Google Scholar] [CrossRef]
- Agalou, A.; Purwantomo, S.; Övernäs, E.; Johannesson, H.; Zhu, X.Y.; Estiati, A.; de Kam, R.J.; Engstrom, P.; Slamet-Loedin, I.H.; Zhu, Z.; et al. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol. Biol. 2008, 66, 87–103. [Google Scholar] [CrossRef]
- Wang, Y.J.; Li, Y.D.; Luo, G.Z.; Tian, A.G.; Wang, H.W.; Zhang, J.S.; Chen, S.Y. Cloning and characterization of an HDZip I gene GmHZ1 from soybean. Planta 2005, 221, 831–843. [Google Scholar] [CrossRef]
- Cabello, J.V.; Arce, A.L.; Chan, R.L. The homologous HDZip I transcription factors HaHB1 and AtHB13 confer cold tolerance via the induction of pathogenesis-related and glucanase proteins. Plant J. 2011, 69, 141–153. [Google Scholar] [CrossRef]
- Manavella, P.A.; Dezar, C.A.; Ariel, F.D.; Drincovich, M.F.; Chan, R.L. The sunflower HD-Zip transcription factor HAHB4 is up-regulated in darkness, reducing the transcription of photosynthesis-related genes. J. Exp. Bot. 2008, 59, 3143–3155. [Google Scholar] [CrossRef] [Green Version]
- Manavella, P.A.; Arce, A.L.; Dezar, C.A.; Bitton, F.; Renou, J.P.; Crespi, M.; Chan, R.L. Cross-talk between ethylene and drought signalling pathways is mediated by the sunflower HaHB-4 transcription factor. Plant J. 2006, 48, 125–137. [Google Scholar] [CrossRef]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Gao, S.; Zhou, X.; Chellappan, P.; Chen, Z.; Zhou, X.; Zhang, X.; Fromuth, N.; Coutino, G.; Coffey, M.; et al. Bacteria-responsive microRNAs regulate plant innate immunity by modulating plant hormone networks. Plant Mol. Biol. 2011, 75, 205–206. [Google Scholar] [CrossRef]
- Schommer, C.; Palatnik, J.F.; Aggarwal, P.; Chétela, A.; Cubas, P.; Farmer, E.E.; Nath, U.; Weigel, D. Contron of jasmonate biosynthesis and senescence by miR319 targets. PLoS Biol. 2008, 6, e230. [Google Scholar] [CrossRef]
- Jin, W.B.; Wu, F.L. Characterization of miRNAs associated with Botrytis cinerea infection of tomato leaves. BMC Plant Biol. 2015, 15, 1. [Google Scholar] [CrossRef]
- Chen, H.Y.; Chen, X.L.; Chen, D.; Li, J.F.; Zhang, Y.; Wang, A.X. A comparison of the low temperature transcriptomes of two tomato genotypes that differ in freezing tolerance: Solanum lycopersicum and Solanum habrochaites. BMC Plant Biol. 2015, 15, 132. [Google Scholar] [CrossRef]
- Zhou, M.; Li, D.; Li, Z.; Yang, C.; Zhu, L.; Luo, H. Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass. Plant Physiol. 2013, 161, 1375–1391. [Google Scholar] [CrossRef]
- Reymond, P.; Farmer, E.E. Jasmonate and salicylate as global signals for defense gene expression. Curr. Opin. Plant Biol. 1998, 1, 404–411. [Google Scholar] [CrossRef]
- Reymond, P.; Weber, H.; Damond, M.; Fanner, E.E. Differential gene expression in response to mechanical wounding and insect feeding in arabidopsis. Plant Cell 2000, 12, 707–719. [Google Scholar] [CrossRef]
- Birkett, M.A.; Campbeel, C.A.; Chamberlain, K.; Guerrieri, E.; Hick, A.J.; Martn, J.L.; Mathes, M.; Napier, J.A.; Pickett, J.A.; Poppy, G.M.; et al. New roles for cis-jasmone as an insect semiochemical and in plant defense. Proc. Natl. Acad. Sci. USA 2000, 97, 9329–9334. [Google Scholar] [CrossRef]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Tian, X.; Song, L.P.; Wang, Y.; Jin, W.B.; Tong, F.D.; Wu, F.L. MiR394 acts as a negative regulator of arabidopsis resistance to B. cinerea infection by targeting LCR. Front. Plant Sci. 2018, 9, 903. [Google Scholar] [CrossRef]
- Li, H.S.; Lin, Y.L.; Chen, X.H.; Bai, Y.; Wang, C.Q.; Xu, X.P.; Wang, Y.; Lai, Z.X. Effects of blue light on flavonoid accumulation linked to the expression of miR393, miR394 and miR395 in longan embryogenic calli. PLoS ONE 2018, 13, e0191444. [Google Scholar] [CrossRef]
- Ghorecha, V.; Zheng, Y.; Liu, L.; Sunkar, R.; Krishnayya, N.S.R. MicroRNA dynamics in a wild and cultivated specie s of convolvulaceae exposed to drought stress. Physiol. Mol. Biol. Plants 2017, 23, 291–300. [Google Scholar] [CrossRef]
- Ferreira, T.H.; Gentile, A.; Vilela, R.D.; Costa, G.G.L.; Dias, L.I.; Endres, L.; Menossi, M. MicroRNAs associated with drought response in the bioenergy crop sugarcane (Saccharum spp.). PLoS ONE 2012, 7, e46703. [Google Scholar] [CrossRef]
- Ni, Z.Y.; Hu, Z.; Jiang, Q.Y.; Zhang, H. Overexpression of Gma-miR394a confers tolerance to drought in transgenic Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2012, 427, 330–335. [Google Scholar] [CrossRef]
- Sunkar, R.; Zhu, J.K. Novel and stress-regulated microRNAs and other small RNAs from arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-regulating factors (GRFs): A small transcription factor family with important functions in plant biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef]
- Liu, H.H.; Tian, X.; Li, Y.J.; Wu, C.A.; Zheng, C.C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef]
- Hewezi, T.; Howe, P.; Maier, T.R.; Baum, T.J. Arabidopsis small RNAs and their targets during cyst nematode parasitism. Mol. Plant Microbe Interact. 2008, 21, 1622–1634. [Google Scholar] [CrossRef]
- Hewezi, T.; Maier, T.R.; Nettleton, D.; Baum, T.J. The arabidopsis microRNA396-GRF1/GRF3 regulatory module acts as a developmental regulator in the reprogramming of root cells during cyst nematode infection. Plant Physiol. 2012, 159, 321–335. [Google Scholar] [CrossRef]
- Liu, D.M.; Song, Y.; Chen, Z.X.; Yu, D.Q. Ectopic expression of miR396 suppresses GRF target gene expression and alters leaf growth in arabidopsis. Physiol. Plant. 2009, 136, 223–236. [Google Scholar] [CrossRef]
- Chen, L.; Luan, Y.S.; Zhai, J.M. Sp-miR396a-5p acts as a stress-responsive genes regulator by conferring tolerance to abiotic stresses and susceptibility to Phytophthora nicotianae infection in transgenic tobacco. Plant Cell Rep. 2015, 34, 2013–2025. [Google Scholar] [CrossRef]
- Tang, C.; Xie, Y.M.; Yan, W. AASRA: An anchor alignment-based small RNA annotation pipeline. BioRxiv 2017, 132918. [Google Scholar]
- Wang, K.; Liang, C.; Liu, J.D.; Xiao, H.M.; Huang, S.P.; Xu, J.H.; Li, F. Prediction of piRNAs using transposon interaction and a support vector machine. BMC Bioinform. 2014, 15, 419. [Google Scholar] [CrossRef]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, 451–454. [Google Scholar] [CrossRef]
- Wu, H.J.; Ma, Y.K.; Chen, T.; Wang, M.; Wang, X.J. PsRobot: A web-based plant small RNA metaanalysis toolbox. Nucleic Acids Res. 2012, 40, 22–28. [Google Scholar] [CrossRef]
- Fahlgren, N.; Carrington, J.C. MiRNA target prediction in plants. Methods Mol. Biol. 2009, 592, 51–57. [Google Scholar]
- Wong, W.H. Statistical inferences for isoform expression in RNASeq. Bioinformatics 2009, 25, 1026–1032. [Google Scholar]
- Wang, L.K.; Feng, Z.X.; Wang, X.; Wang, X.W.; Zhang, X.G. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2011, 26, 136–138. [Google Scholar] [CrossRef]
- Yang, Y.H.; Dudoit, S.; Luu, P.; Peng, V.; Ngai, J.; Speed, T.P. Normalization for cDNA microarray data: A robust composite method addressing single and multiple slide systematic variation. Nucleic Acids Res. 2002, 30, e15. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; Marcelino-Guimaraes, F.C.; Nepomuceno, A.L.; Abdelnoor, R.V.; Margis, R. The use of microRNAs as reference gene for quantitative polymerase chain reaction in soybean. Anal. Biochem. 2010, 406, 185–192. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | HRK0-1 | HRK0-2 | HRK48-1 | HRK48-2 | HSK0-1 | HSK0-2 | HSK48-1 | HSK48-2 |
|---|---|---|---|---|---|---|---|---|
| Total reads | 18187564 | 15645006 | 15811473 | 22213829 | 15877691 | 16418090 | 19154749 | 19705533 |
| Clean reads | 16725511 (91.96%) | 14594887 (93.29%) | 14335242 (90.66%) | 20230796 (91.07%) | 14697929 (92.57%) | 15060199 (91.73%) | 17541500 (91.58%) | 18523834 (94.00%) |
| Mapped reads | 15092682 (90.24%) | 13055191 (89.45%) | 12999357 (90.68%) | 18078009 (89.36%) | 13090959 (89.07%) | 13620528 (90.44%) | 15751832 (89.80%) | 16892630 (91.19%) |
| Intergenic | 9443266 (56.46%) | 7076994 (48.49%) | 7502556 (52.34%) | 10774382 (53.26%) | 8035260 (54.67%) | 7314000 (48.57%) | 9032395 (51.49%) | 9351270 (50.48%) |
| Exon | 2296806 (13.73%) | 2735655 (18.74%) | 2230122 (15.56%) | 3074662 (15.20%) | 2018611 (13.73%) | 2881758 (19.13%) | 2785970 (15.88%) | 3185485 (17.20%) |
| Intron | 2197086 (13.14%) | 2293967 (15.72%) | 2330038 (16.25%) | 2958235 (14.62%) | 2056682 (13.99%) | 2628373 (17.45%) | 2986540 (17.03%) | 3410640 (18.41%) |
| miRNA | 929540 (5.56%) | 607305 (4.16%) | 536353 (3.74%) | 959353 (4.74%) | 748416 (5.09%) | 528713 (3.51%) | 641115 (3.65%) | 584366 (3.15%) |
| rRNA | 211127 (1.26%) | 345408 (2.37%) | 395420 (2.76%) | 295084 (1.46%) | 233612 (1.59%) | 262057 (1.74%) | 294055 (1.68%) | 346254 (1.87%) |
| snoRNA | 5719 (0.03%) | 6779 (0.05%) | 5929 (0.04%) | 9232 (0.05%) | 4486 (0.03%) | 6169 (0.04%) | 7500 (0.04%) | 8553 (0.05%) |
| tRNA | 1767 (0.01%) | 2581 (0.02%) | 5226 (0.04%) | 6462 (0.03%) | 1742 (0.01%) | 1970 (0.01%) | 1915 (0.07%) | 3580 (0.02%) |
| sncRNA | 4662 (0.03%) | 5472 (0.04%) | 7182 (0.05%) | 7141 (0.04%) | 5164 (0.04%) | 5714 (0.04%) | 5719 (0.03%) | 5406 (0.03%) |
| Precursor | 30472 (0.18%) | 7206 (0.05%) | 6290 (0.04%) | 34022 (0.17%) | 5193 (0.04%) | 7113 (0.05%) | 2449 (0.01%) | 51321 (0.28%) |
| Unmap | 1604212 (9.59%) | 1518313 (10.40%) | 1312486 (9.17%) | 2110698 (10.43%) | 1588107 (10.80%) | 1423625 (9.45%) | 1771428 (10.10%) | 1574369 (8.50%) |
| Type | Highly-Conserved | Moderately-Conserved | Lowly-Conserved | Non-Conserved | Total |
|---|---|---|---|---|---|
| No. of known miRNAs | 188 | 23 | 32 | 185 | 428 |
| No. of known miRNAs families | 23 | 3 | 20 | 146 | 192 |
| Control Groups | miRNA | miRNA_log2Ratio (Fold Change) | mRNA | mRNA_log2Ratio (Fold Change) | NR Description |
|---|---|---|---|---|---|
| HRK48/HRK0 | Gma-miR1535a | 1.57 | Glyma.08G264900.2 | −2.25 | phototropin-2-like |
| Gma-miR166u | −1.70 | Glyma.18G204800.1 | 3.42 | ammonium transporter AMT2.2 | |
| Gma-miR394a-3p | −2.26 | Glyma.14G004300.1 | 3.71 | nudix hydrolase 2-like | |
| Gma-miR394a-3p | −2.26 | Glyma.05 G073500.2 | 7.12 | F-box protein SKIP14-like | |
| Gma-miR4996 | 1.77 | Glyma.U019800.1 | −7.57 | auxin response factor 4-like | |
| Gma-miR5374-3p | −2.20 | Glyma.17G128200.2 | 7.21 | cyclin-dependent kinase G-2-like | |
| Novel_miR2 | 1.19 | Glyma.05G167800.2 | −3.78 | guanosine nucleotide diphosphate dissociation inhibitor 2 | |
| HSK48/HSK0 | Gma-miR1512b | 3.10 | Glyma.09G019300.1 | −6.94 | protein FAR1-RELATED SEQUENCE 6 |
| Gma-miR1535a | 2.22 | Glyma.19G036600.2 | −7.41 | probable leucine-rich repeat receptor-like serine/threonine-protein kinase At5g15730 | |
| Gma-miR166j-3p | −1.25 | Glyma.07G016700.2 | 8.54 | homeobox-leucine zipper protein ATHB-15-like isoform X4 | |
| Gma-miR166u | −1.42 | Glyma.07G016700.2 | 8.54 | homeobox-leucine zipper protein ATHB-15-like isoform X4 | |
| Gma-miR395g | −1.21 | Glyma.14G005800.3 | 8.16 | nuclear pore complex protein NUP96 | |
| Gma-miR395g | −1.21 | Glyma.14G005800.4 | 8.10 | nuclear pore complex protein NUP96 | |
| Gma-miR399b | −2.31 | Glyma.03G021900.2 | 7.06 | uncharacterized protein LOC100306494 isoform X1 | |
| Gma-miR5761a | 2.34 | Glyma.11G210400.3 | −2.68 | glycinol 4-dimethylallyltransferase-like | |
| Gma-miR9749 | −1.30 | Glyma.09G236600.3 | 7.32 | lipoate-protein ligase LplJ | |
| Novel_miR2 | 3.13 | Glyma.05G167800.2 | −9.51 | guanosine nucleotide diphosphate dissociation inhibitor 2 | |
| HRK0/HSK0 | Gma-miR156q | −1.10 | Glyma.06G238100.1 | 2.37 | squamosa promoter-binding protein 1-like |
| Gma-miR319d | 1.65 | Glyma.08G087400.1 | −5.22 | uncharacterized protein LOC100778760 | |
| Gma-miR394a-3p | 1.66 | Glyma.05G073500.2 | −7.96 | F-box protein SKIP14-like | |
| Gma-miR396e | 3.89 | Glyma.13G159700.1 | −6.85 | hypothetical protein GLYMA_13G159700 | |
| Gma-miR4996 | −3.14 | Glyma.06G040600.1 | 9.05 | THO complex subunit 5A-like isoform X1 | |
| Gma-miR5769 | −1.27 | Glyma.02G186100.2 | 8.96 | calcium-transporting ATPase 4, plasma membrane-type-like | |
| Novel_miR36 | 1.99 | Glyma.07G178800.2 | −6.80 | pentatricopeptide repeat-containing protein At2g15820, chloroplastic-like | |
| HRK48/HSK48 | Gma-miR166b | 4.63 | Glyma.07G016700.2 | −8.54 | homeobox-leucine zipper protein ATHB-15-like isoform X4 |
| Gma-miR166j-3p | 1.63 | Glyma.07G016700.2 | −8.54 | homeobox-leucine zipper protein ATHB-15-like isoform X4 | |
| Gma-miR395g | 1.38 | Glyma.14G005800.3 | −8.16 | nuclear pore complex protein NUP96 | |
| Gma-miR398b | −1.47 | Glyma.01G055700.2 | 3.19 | hypothetical protein GLYMA_01G055700 | |
| Gma-miR4416a | −1.15 | Glyma.19G136400.2 | 7.88 | putative calcium-transporting ATPase 11, plasma membrane-type | |
| Gma-miR4996 | −1.73 | Glyma.06G040600.1 | 8.90 | THO complex subunit 5A-like | |
| Gma-miR4996 | −1.73 | Glyma.07G002800.2 | 7.50 | ubiquitin receptor RAD23b-like | |
| Gma-miR5761a | −1.30 | Glyma.13G004400.1 | 2.45 | zinc transporter 8-like | |
| Novel_miR36 | 2.66 | Glyma.06G088200.3 | −7.97 | amino acid permease 6-like |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, W.; Sun, Z.; Lai, Z.; Yang, S.; Chen, H.; Yang, X.; Tao, J.; Tang, X. Determination of the MiRNAs Related to Bean Pyralid Larvae Resistance in Soybean Using Small RNA and Transcriptome Sequencing. Int. J. Mol. Sci. 2019, 20, 2966. https://doi.org/10.3390/ijms20122966
Zeng W, Sun Z, Lai Z, Yang S, Chen H, Yang X, Tao J, Tang X. Determination of the MiRNAs Related to Bean Pyralid Larvae Resistance in Soybean Using Small RNA and Transcriptome Sequencing. International Journal of Molecular Sciences. 2019; 20(12):2966. https://doi.org/10.3390/ijms20122966
Chicago/Turabian StyleZeng, Weiying, Zudong Sun, Zhenguang Lai, Shouzhen Yang, Huaizhu Chen, Xinghai Yang, Jiangrong Tao, and Xiangmin Tang. 2019. "Determination of the MiRNAs Related to Bean Pyralid Larvae Resistance in Soybean Using Small RNA and Transcriptome Sequencing" International Journal of Molecular Sciences 20, no. 12: 2966. https://doi.org/10.3390/ijms20122966
APA StyleZeng, W., Sun, Z., Lai, Z., Yang, S., Chen, H., Yang, X., Tao, J., & Tang, X. (2019). Determination of the MiRNAs Related to Bean Pyralid Larvae Resistance in Soybean Using Small RNA and Transcriptome Sequencing. International Journal of Molecular Sciences, 20(12), 2966. https://doi.org/10.3390/ijms20122966