Tandem 13-Lipoxygenase Genes in a Cluster Confers Yellow-Green Leaf in Cucumber



Abstract
:1. Introduction
2. Result
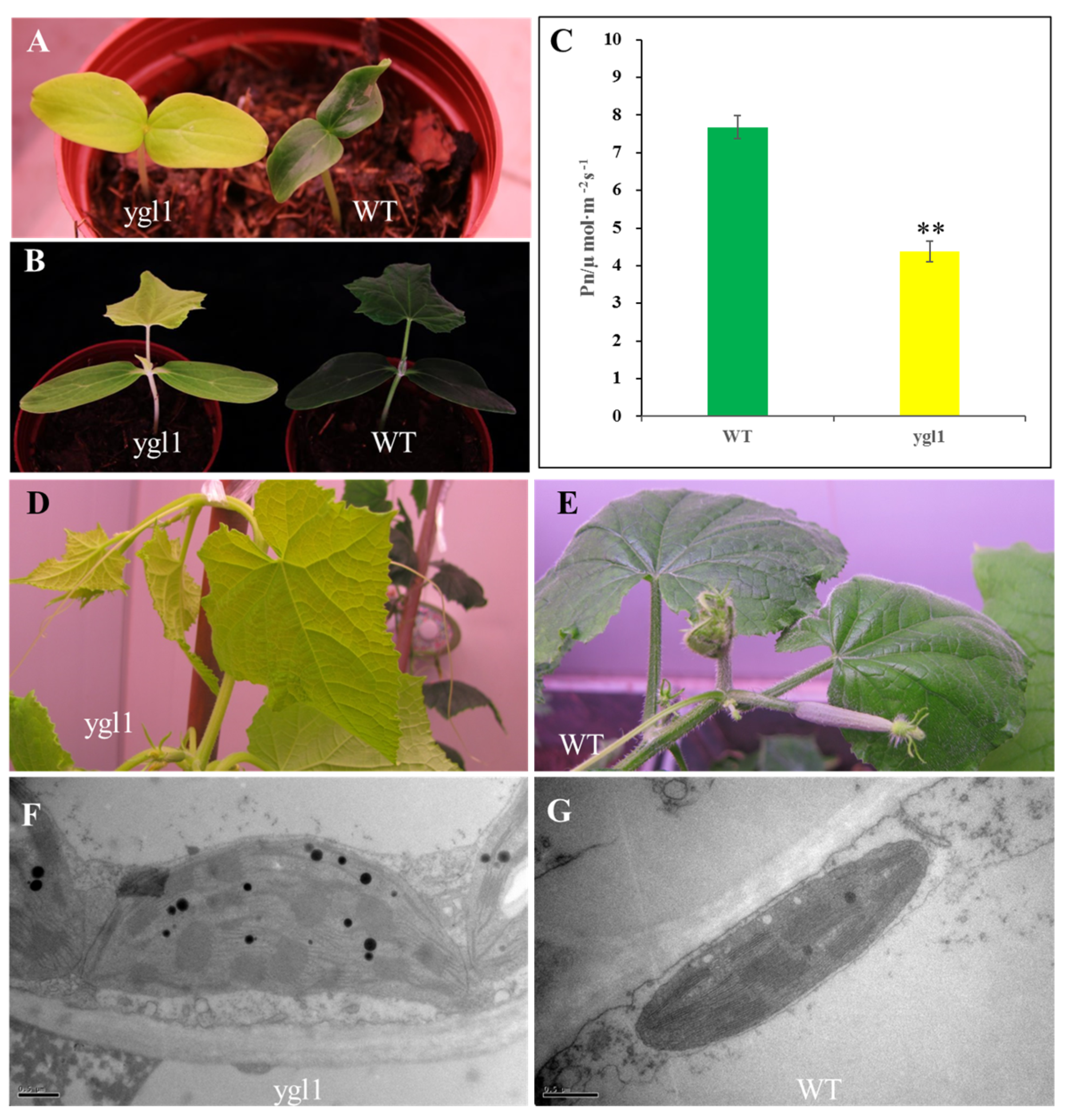
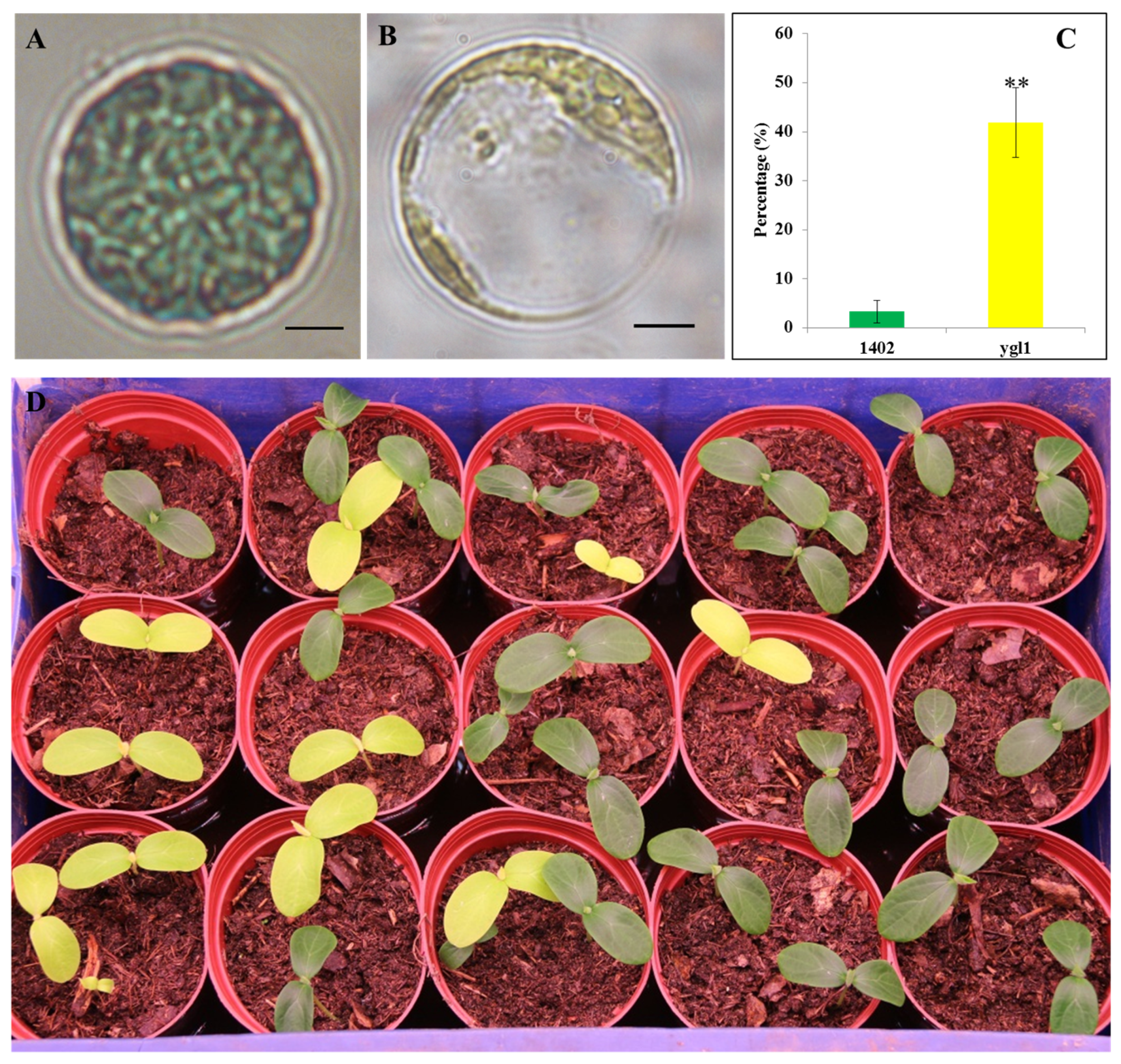
2.1. ygl1 Mutant Has Reduced Chl Accumulation and Abnormal Chloroplast Development
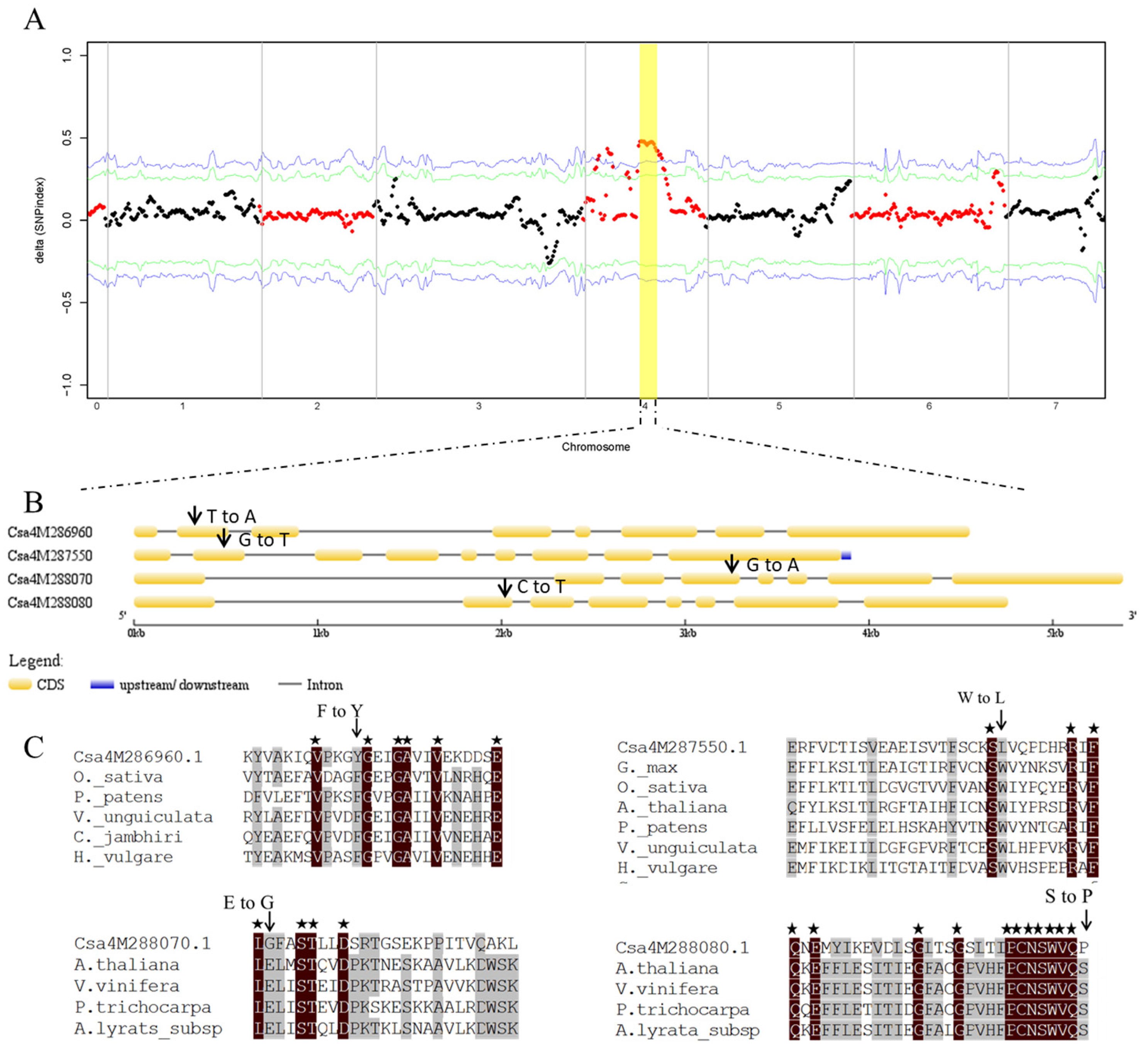
2.2. Identification of the Candidate Gene for ygl1
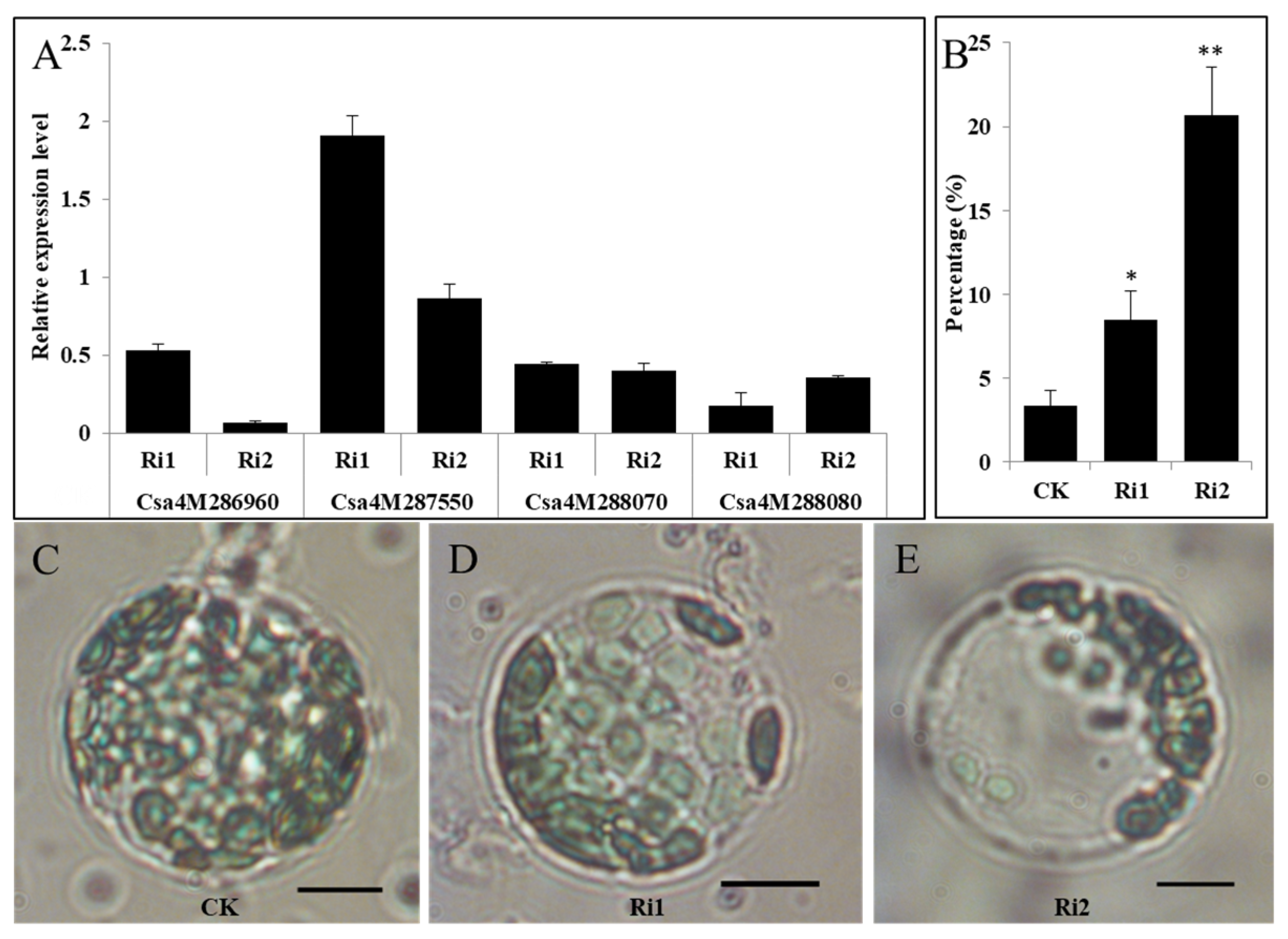
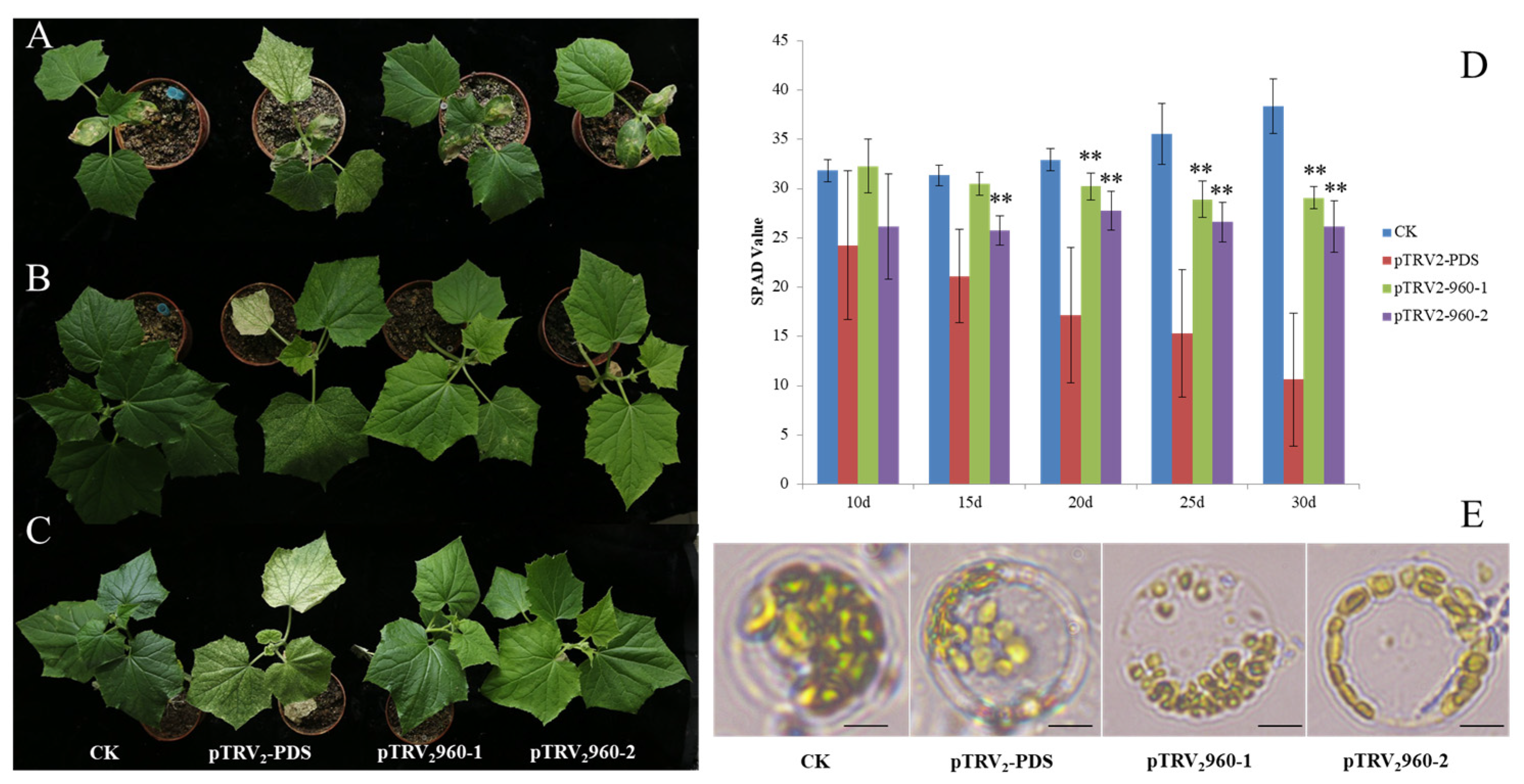
2.3. Abnormal Chloroplast Morphology in LOX Genes Knockdown Cucumber
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Net Photosynthetic Rate
4.3. Transmission Electron Microscopy (TEM) Analysis
4.4. Chloroplast Phenotype Analysis
4.5. Whole Genome Resequencing
4.6. Sequence Data Analysis
4.7. Sequence Alignments and Phylogenetic Analysis
4.8. Development of PCR-Based SNP Markers
4.9. Transient Gene Silencing in Cucumber Cotyledons
4.10. Chl Content and SPAD Assay
4.11. Plasmid Construction and Agro-infiltration
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ming, M.; Ying, W.; Qibing, L.; Yihua, W.; Zhijun, C. Identification and gene mapping of a yellow-leaf rice mutant ys94. Crops 2016, 4, 41–46. [Google Scholar]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? PlantCell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef]
- Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci. 1998, 3, 147–151. [Google Scholar] [CrossRef]
- Fromme, P.; Melkozernov, A.; Jordan, P.; Krauss, N. Structure and function of photosystem i: Interaction with its soluble electron carriers and external antenna systems. FEBS Lett 2003, 555, 40–44. [Google Scholar] [CrossRef]
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Fei, G.-L.; Wu, C.-Y.; Wu, F.-Q.; Sun, Y.-Y.; Chen, M.-J.; Ren, Y.-L.; Zhou, K.-N.; Cheng, Z.-J.; Wang, J.-L. A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants. Plant Physiol. 2013, 162, 1867–1880. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, Y.; Wang, P.; Li, C.; Xiao, F.; Chen, N.; Li, N.; Li, C.; Sun, C.; Li, L. A single nucleotide mutation of ispf gene involved in the mep pathway for isoprenoid biosynthesis causes yellow-green leaf phenotype in rice. Plant Mol. Biol. 2018, 96, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-p.; Yang, C.; Han, F.-q.; Fang, Z.-y.; Yang, L.-m.; Zhuang, M.; Lv, H.-h.; Liu, Y.-m.; Li, Z.-s.; Zhang, Y.-y. Genetics and fine mapping of a yellow-green leaf gene (ygl-1) in cabbage (brassica oleracea var. Capitata l.). Mol. Breed. 2016, 36, 1–8. [Google Scholar] [CrossRef]
- Nothnagel, T.; Straka, P. Inheritance and mapping of a yellow leaf mutant of carrot (daucus carota). Plant Breed. 2003, 122, 339–342. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, Z.; Shan, X.; Li, C.; Tang, X.; Chi, M.; Feng, H. Physiological properties and chlorophyll biosynthesis in a pak-choi (brassica rapa l. Ssp. Chinensis) yellow leaf mutant, pylm. Acta Physiol. Plant. 2017, 39, 22. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, X.; He, B.; Diao, L.; Sheng, S.; Wang, J.; Guo, X.; Su, N.; Wang, L.; Jiang, L.; et al. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol 2007, 145, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Hu, L.; Li, Y.; Weng, Y. The chlorophyll-deficient golden leaf mutation in cucumber is due to a single nucleotide substitution in cschli for magnesium chelatase i subunit. Theor. Appl. Genet. 2016, 129, 1961–1973. [Google Scholar] [CrossRef] [PubMed]
- Beale, S.I. Green genes gleaned. Trends Plant Sci 2005, 10, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Nagata, N.; Tanaka, R.; Satoh, S.; Tanaka, A. Identification of a vinyl reductase gene for chlorophyll synthesis in arabidopsis thaliana and implications for the evolution of prochlorococcus species. Plant Cell 2005, 17, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using mutmap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Monde, R.A.; Zito, F.; Olive, J.; Wollman, F.A.; Stern, D.B. Post-transcriptional defects in tobacco chloroplast mutants lacking the cytochrome b6/f complex. Plant J. 2000, 21, 61–72. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, M.L.; Zhang, Y.Z.; Du, L.F.; Pan, T. A chlorophyll-reduced seedling mutant in oilseed rape, brassica napus, for utilization in f-1 hybrid production. Plant Breed. 2000, 119, 131–135. [Google Scholar] [CrossRef]
- Grechkin, A. Recent developments in biochemistry of the plant lipoxygenase pathway. Prog. Lipid Res. 1998, 37, 317–352. [Google Scholar] [CrossRef]
- Kuhn, H.; Saam, J.; Eibach, S.; Holzhütter, H.-G.; Ivanov, I.; Walther, M. Structural biology of mammalian lipoxygenases: Enzymatic consequences of targeted alterations of the protein structure. Biochem. Biophys. Res. Commun. 2005, 338, 93–101. [Google Scholar] [CrossRef]
- Ivanov, I.; Heydeck, D.; Hofheinz, K.; Roffeis, J.; O’Donnell, V.B.; Kuhn, H.; Walther, M. Molecular enzymology of lipoxygenases. Arch. Biochem. Biophys. 2010, 503, 161–174. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Mathew, G.; Jayne, D.G.; Pelengaris, S.; Khan, M. 15-lipoxygenase-1 in colorectal cancer: A review. Tumour Biol. 2009, 30, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Chawengsub, Y.; Gauthier, K.M.; Campbell, W.B. Role of arachidonic acid lipoxygenase metabolites in the regulation of vascular tone. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H495–H507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, A.P. Lipoxygenase--a versatile biocatalyst for biotransformation of endobiotics and xenobiotics. Cell Mol Life Sci 2001, 58, 1805–1825. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Liavonchanka, A.; Feussner, I. Lipoxygenases: Occurrence, functions and catalysis. J. Plant Physiol. 2006, 163, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Chedea, V.S.; Jisaka, M. Lipoxygenase and carotenoids: A co-oxidation story. Afr. J. Biotechnol. 2013, 12. [Google Scholar]
- Gayen, D.; Ali, N.; Sarkar, S.N.; Datta, S.K.; Datta, K. Down-regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta 2015, 242, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Robinson, D.S.; Hughes, R.K.; Casey, R.; Hardy, D.; West, S.I. Co-oxidation of β-carotene catalyzed by soybean and recombinant pea lipoxygenases. J. Agric. Food Chem. 1999, 47, 4899–4906. [Google Scholar] [CrossRef]
- Cabibel, M.; Nicolas, J. Lipoxygenase from tomato fruit (lycopersicon esculentum l.). Partial purification, some properties and in vitro cooxidation of some carotenoid pigments. Sci. Des Aliment. 1991, 11, 177–290. [Google Scholar]
- Biacs, P.; Daood, H. Lipoxygenase-catalysed degradation of carotenoids from tomato in the presence of antioxidant vitamins. Biochem. Soc. Trans. 2000, 28, 839–845. [Google Scholar] [CrossRef]
- Aziz, S.; Wu, Z.; Robinson, D.S. Potato lipoxygenase catalysed co-oxidation of β-carotene. Food Chem. 1999, 64, 227–230. [Google Scholar] [CrossRef]
- Sarde, S.J.; Kumar, A.; Remme, R.N.; Dicke, M. Genome-wide identification, classification and expression of lipoxygenase gene family in pepper. Plant Mol. Biol. 2018, 98, 375–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.W.; Pan, J.S.; He, H.L.; Zhang, C.; Li, Z.; Zhao, J.L.; Yuan, X.J.; Zhu, L.H.; Huang, S.W.; Cai, R. Construction of a high density integrated genetic map for cucumber (Cucumis sativus L.). Appl. Genet. 2012, 124, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, X.; Guo, L.; Xu, Q.; Zhao, S.; Li, F.; Yan, X.; Liu, S.; Wei, C. Characterization and alternative splicing profiles of lipoxygenase gene family in tea plant (camellia sinensis). Plant Cell Physiol. 2018, 59, 1765–1781. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, X.; Yan, H.; Li, W.; Li, Y.; Cai, R.; Xiang, Y. The lipoxygenase gene family in poplar: Identification, classification, and expression in response to meja treatment. PLoS ONE 2015, 10, e0125526. [Google Scholar] [CrossRef] [PubMed]
- Umate, P. Genome-wide analysis of lipoxygenase gene family in arabidopsis and rice. Plant Signal. Behav. 2011, 6, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Melan, M.A.; Dong, X.; Endara, M.E.; Davis, K.R.; Ausubel, F.M.; Peterman, T.K. An arabidopsis thaliana lipoxygenase gene can be induced by pathogens, abscisic acid, and methyl jasmonate. Plant Physiol. 1993, 101, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar] [CrossRef]
- Grebner, W.; Stingl, N.; Oenel, A.; Mueller, M.J.; Berger, S. Lipoxygenase 6-dependent oxylipin synthesis in roots is required for abiotic and biotic stress resistance of arabidopsis thaliana. Plant Physiol. 2013, 161, 2159–2170. [Google Scholar] [CrossRef]
- Mochizuki, S.; Sugimoto, K.; Koeduka, T.; Matsui, K. Arabidopsis lipoxygenase 2 is essential for formation of green leaf volatiles and five-carbon volatiles. FEBS Lett. 2016, 590, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Caldelari, D.; Wang, G.; Farmer, E.E.; Dong, X. Arabidopsis lox3 lox4 double mutants are male sterile and defective in global proliferative arrest. Plant Mol. Biol. 2011, 75, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Vellosillo, T.; Martínez, M.; López, M.A.; Vicente, J.; Cascón, T.; Dolan, L.; Hamberg, M.; Castresana, C. Oxylipins produced by the 9-lipoxygenase pathway in arabidopsis regulate lateral root development and defense responses through a specific signaling cascade. Plant Cell 2007, 19, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, A.; Barry, C.; Alpuche-Solis, A.G.; Grierson, D. Ethylene and developmental signals regulate expression of lipoxygenase genes during tomato fruit ripening. J. Exp. Bot. 1999, 50, 793–798. [Google Scholar] [CrossRef]
- Ferrie, B.J.; Beaudoin, N.; Burkhart, W.; Bowsher, C.G.; Rothstein, S.J. The cloning of two tomato lipoxygenase genes and their differential expression during fruit ripening. Plant Physiol. 1994, 106, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Hackett, R.; Walker, D.; Taylor, A.; Lin, Z.; Grierson, D. Identification of a specific isoform of tomato lipoxygenase (tomloxc) involved in the generation of fatty acid-derived flavor compounds. Plant Physiol. 2004, 136, 2641–2651. [Google Scholar] [CrossRef]
- Yan, L.; Zhai, Q.; Wei, J.; Li, S.; Wang, B.; Huang, T.; Du, M.; Sun, J.; Kang, L.; Li, C.-B. Role of tomato lipoxygenase d in wound-induced jasmonate biosynthesis and plant immunity to insect herbivores. PLoS Genet. 2013, 9, e1003964. [Google Scholar] [CrossRef]
- Mariutto, M.; Duby, F.; Adam, A.; Bureau, C.; Fauconnier, M.-L.; Ongena, M.; Thonart, P.; Dommes, J. The elicitation of a systemic resistance by pseudomonas putida btp1 in tomato involves the stimulation of two lipoxygenase isoforms. BMC Plant Biol. 2011, 11, 29. [Google Scholar] [CrossRef]
- Allmann, S.; Halitschke, R.; Schuurink, R.C.; Baldwin, I.T. Oxylipin channelling in nicotiana attenuata: Lipoxygenase 2 supplies substrates for green leaf volatile production. PlantCell Environ. 2010, 33, 2028–2040. [Google Scholar] [CrossRef]
- VanDoorn, A.; Kallenbach, M.; Borquez, A.A.; Baldwin, I.T.; Bonaventure, G. Rapid modification of the insect elicitor n-linolenoyl-glutamate via a lipoxygenase-mediated mechanism on nicotiana attenuata leaves. BMC Plant Biol. 2010, 10, 164. [Google Scholar] [CrossRef]
- León, J.; Royo, J.; Vancanneyt, G.; Sanz, C.; Silkowski, H.; Griffiths, G.; Sánchez-Serrano, J.J. Lipoxygenase h1 gene silencing reveals a specific role in supplying fatty acid hydroperoxides for aliphatic aldehyde production. J. Biol. Chem. 2002, 277, 416–423. [Google Scholar] [CrossRef]
- Hwang, I.S.; Hwang, B.K. The pepper 9-lipoxygenase gene calox1 functions in defense and cell death responses to microbial pathogens. Plant Physiol. 2010, 152, 948–967. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.; Kang, C.; Rustgi, S.; Von Wettstein, D.; Reinbothe, C.; Pollmann, S.; Reinbothe, S. Programmed chloroplast destruction during leaf senescence involves 13-lipoxygenase (13-lox). Proc. Natl. Acad. Sci. USA 2016, 113, 3383–3388. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Li, S.; Li, M.F.; Yang, X.L.; Wang, W.T.; Cao, Z.Y.; Li, W. Physiological characteristics and leaf ultrastructure of a novel chlorophyll-deficient chd6 mutant of vitis venifera cultured in vitro. J. Plant Growth Regul. 2012, 31, 124–135. [Google Scholar] [CrossRef]
- Rohacek, K.; Bartak, M. Technique of the modulated chlorophyll fluorescence: Basic concepts, useful parameters, and some applications. Photosynthetica 1999, 37, 339–363. [Google Scholar] [CrossRef]
- Kodis, G.; Herrero, C.; Palacios, R.; Marino-Ochoa, E.; Gould, S.; De La Garza, L.; Van Grondelle, R.; Gust, D.; Moore, T.A.; Moore, A.L. Light harvesting and photoprotective functions of carotenoids in compact artificial photosynthetic antenna designs. J. Phys. Chem. B 2004, 108, 414–425. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef]
- Mayne, S. Carotenoids a Colorful and Timely Research Field; CRC Press, Taylor and Francisc Group: Boca Raton, FL, USA, 2010. [Google Scholar]
- Hulbert, S.H.; Webb, C.A.; Smith, S.M.; Sun, Q. Resistance gene complexes: Evolution and utilization. Annu Rev. Phytopathol. 2001, 39, 285–312. [Google Scholar] [CrossRef]
- Smith, L.M.; Bomblies, K.; Weigel, D. Complex evolutionary events at a tandem cluster of arabidopsis thaliana genes resulting in a single-locus genetic incompatibility. PLoS Genet. 2011, 7, e1002164. [Google Scholar] [CrossRef]
- Zhao, H.; Yu, L.; Huai, Z.X.; Wang, X.H.; Ding, G.D.; Chen, S.S.; Li, P.; Xu, F.S. Mapping and candidate gene identification defining bnchd1-1, a locus involved in chlorophyll biosynthesis in brassica napus. Acta Physiol. Plant. 2014, 36, 859–870. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, S.; Hu, B.; Chen, H.; Zhang, Z.; Huang, S. An accumulation and replication of chloroplasts 5 gene mutation confers light green peel in cucumber. J. Integr. Plant Biol. 2015, 57, 936–942. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version ii. Plant Mol. Biol. Report. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with burrows–wheeler transform. bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S. Qtl-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Ma, Y.; Zhou, Y.; Zhang, H.; Duan, L.; Chen, H.; Zeng, J.; Zhou, Q.; Wang, S.; Gu, W.; et al. Plant science. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148C, 350–382. [Google Scholar]
- Fu, D.Q.; Zhu, B.Z.; Zhu, H.L.; Jiang, W.B.; Luo, Y.B. Virus-induced gene silencing in tomato fruit. Plant J. 2005, 43, 299–308. [Google Scholar] [CrossRef]
- Igarashi, A.; Yamagata, K.; Sugai, T.; Takahashi, Y.; Sugawara, E.; Tamura, A.; Yaegashi, H.; Yamagishi, N.; Takahashi, T.; Isogai, M.; et al. Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among a broad range of plants including tobacco, tomato, arabidopsis thaliana, cucurbits, and legumes. Virology 2009, 386, 407–416. [Google Scholar] [CrossRef]
- Ratcliff, F.; Martin-Hernandez, A.M.; Baulcombe, D.C. Technical advance. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Stage | Genotype | Chl a | Chl b | Car | Car /Pig Percent |
|---|---|---|---|---|---|
| 3 weeks old | Wild type | 2.39 ± 0.20 | 1.31 ± 0.14 | 0.39 ± 0.04 | 9.66 ± 2.60 |
| ygl1 | 0.76 ± 0.03 | 0.25 ± 0.02 | 0.30 ± 0.01 | 22.87 ± 1.25 | |
| 12 weeks old | Wild type | 2.52 ± 0.15 | 1.17 ± 0.07 | 0.30 ± 0.02 | 7.54 ± 0.75 |
| ygl1 | 0.76 ± 0.06 | 0.23 ± 0.05 | 0.27 ± 0.01 | 21.91 ± 1.39 |
| Position | Ref | ygl1 | Gene | Translate | aa | Annotation |
|---|---|---|---|---|---|---|
| 11124523 | A | T | Csa4M286960.1 | TTT<->TAT | F<->Y | Lipoxygenase |
| 11126210 | T | G | Csa4M286960.1 | CCT<->CCG | P<->P | Lipoxygenase |
| 11126739 | A | G | Csa4M286960.1 | TTA<->TTG | L<->L | Lipoxygenase |
| 11135803 | T | C | Csa4M286980.1 | GAC<->GAT | D<->D | Lipoxygenase |
| 11135804 | A | G | Csa4M286980.1 | GAT<->AAT | D<->N | Lipoxygenase |
| 11135807 | T | G | Csa4M286980.1 | TGA<->GGA | *<->G | Lipoxygenase |
| 11141217 | A | G | Csa4M286990.1 | ACG<->ACA | T<->T | Lipoxygenase |
| 11144638 | C | G | Csa4M287010.1 | GCG<->GGG | A<->G | Lipoxygenase |
| 11166969 | T | G | Csa4M287550.1 | TGG<->TTG | W<->L | Lipoxygenase |
| 11167955 | A | C | Csa4M287550.1 | GCT<->GAT | A<->D | Lipoxygenase |
| 11168050 | C | G | Csa4M287550.1 | GTT<->CTT | V<->L | Lipoxygenase |
| 11168212 | T | C | Csa4M287550.1 | TAT<->TAC | Y<->Y | Lipoxygenase |
| 11168253 | T | C | Csa4M287550.1 | GTT<->GCT | V<->A | Lipoxygenase |
| 11173198 | A | G | Csa4M287570.1 | ACA<->ATA | T<->I | Unknown protein |
| 11178904 | A | G | Csa4M288070.1 | TGG<->TAG | W<->* | Lipoxygenase |
| 11179163 | T | G | Csa4M288070.1 | TTG<->TTT | L<->F | Lipoxygenase |
| 11181197 | T | C | Csa4M288070.1 | TCG<->TTG | S<->L | Lipoxygenase |
| 11181330 | A | G | Csa4M288070.1 | CAA<->CAG | Q<->Q | Lipoxygenase |
| 11181348 | A | G | Csa4M288070.1 | CAG<->CAA | Q<->Q | Lipoxygenase |
| 11181801 | T | G | Csa4M288070.1 | TCG<->TCT | S<->S | Lipoxygenase |
| 11181936 | T | C | Csa4M288070.1 | GAT<->GAC | D<->D | Lipoxygenase |
| 11181988 | A | G | Csa4M288070.1 | GAT<->AAT | D<->N | Lipoxygenase |
| 11182010 | T | C | Csa4M288070.1 | CCT<->CTT | P<->L | Lipoxygenase |
| 11182070 | A | G | Csa4M288070.1 | GAA<->GGA | E<->G | Lipoxygenase |
| 11182092 | A | G | Csa4M288070.1 | CAG<->CAA | Q<->Q | Lipoxygenase |
| 11182243 | T | C | Csa4M288070.1 | TTT<->TTC | F<->F | Lipoxygenase |
| 11190230 | T | C | Csa4M288080.1 | ATC<->ACC | I<->T | Lipoxygenase |
| 11192106 | T | C | Csa4M288080.1 | ACT<->ATT | T<->I | Lipoxygenase |
| 11192300 | A | G | Csa4M288080.1 | GTA<->GTG | V<->V | Lipoxygenase |
| 11192333 | A | G | Csa4M288080.1 | CAA<->CAG | Q<->Q | Lipoxygenase |
| 11192417 | T | C | Csa4M288080.1 | CTC<->CTT | L<->L | Lipoxygenase |
| 11193182 | T | C | Csa4M288080.1 | CCT<->TCT | P<->S | Lipoxygenase |
| 11203061 | A | G | Csa4M288110.1 | GCG<->GCA | A<->A | Lipoxygenase |
| 11203109 | T | C | Csa4M288110.1 | TCC<->TCT | S<->S | Lipoxygenase |
| 11203161 | A | T | Csa4M288110.1 | TAT<->AAT | Y<->N | Lipoxygenase |
| 11216771 | A | G | Csa4M288610.1 | ACG<->ACA | T<->T | Lipoxygenase |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.; Yang, W.; Su, C.; Ma, H.; Pan, Y.; Zhang, X.; Li, J. Tandem 13-Lipoxygenase Genes in a Cluster Confers Yellow-Green Leaf in Cucumber. Int. J. Mol. Sci. 2019, 20, 3102. https://doi.org/10.3390/ijms20123102
Ding Y, Yang W, Su C, Ma H, Pan Y, Zhang X, Li J. Tandem 13-Lipoxygenase Genes in a Cluster Confers Yellow-Green Leaf in Cucumber. International Journal of Molecular Sciences. 2019; 20(12):3102. https://doi.org/10.3390/ijms20123102
Chicago/Turabian StyleDing, Yin, Wei Yang, Chenggang Su, Huihui Ma, Yu Pan, Xingguo Zhang, and Jinhua Li. 2019. "Tandem 13-Lipoxygenase Genes in a Cluster Confers Yellow-Green Leaf in Cucumber" International Journal of Molecular Sciences 20, no. 12: 3102. https://doi.org/10.3390/ijms20123102
APA StyleDing, Y., Yang, W., Su, C., Ma, H., Pan, Y., Zhang, X., & Li, J. (2019). Tandem 13-Lipoxygenase Genes in a Cluster Confers Yellow-Green Leaf in Cucumber. International Journal of Molecular Sciences, 20(12), 3102. https://doi.org/10.3390/ijms20123102