NB-LRRs Not Responding Consecutively to Fusarium oxysporum Proliferation Caused Replant Disease Formation of Rehmannia glutinosa

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Changes in the Numbers of Pseudomonas spp. and Fusarium oxysporum in Rhizosphere Soils of Replant Disease R. glutinosa
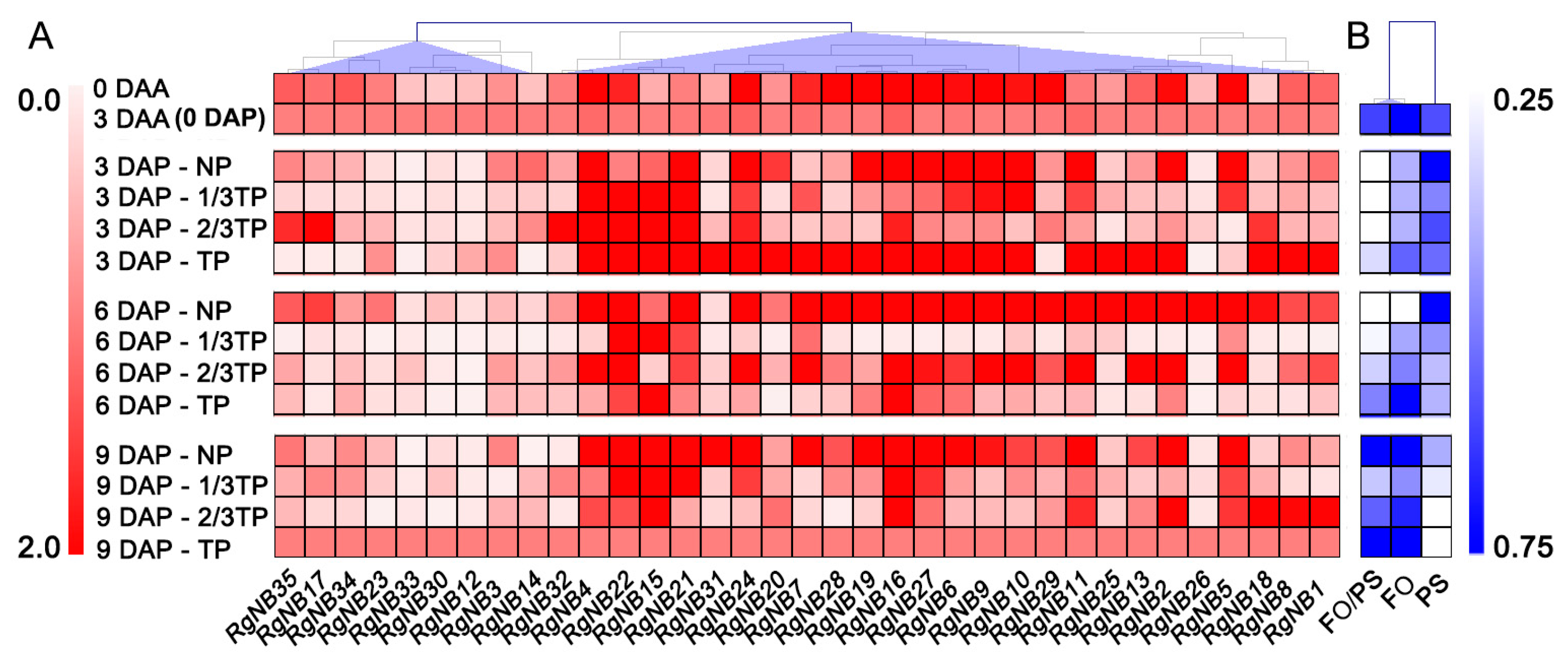
2.2. NB-LRR Lists Response to Replant Disease Stresses in R. glutinosa
2.3. Plant Hormones Response to Replant Disease Stresses in R. glutinosa
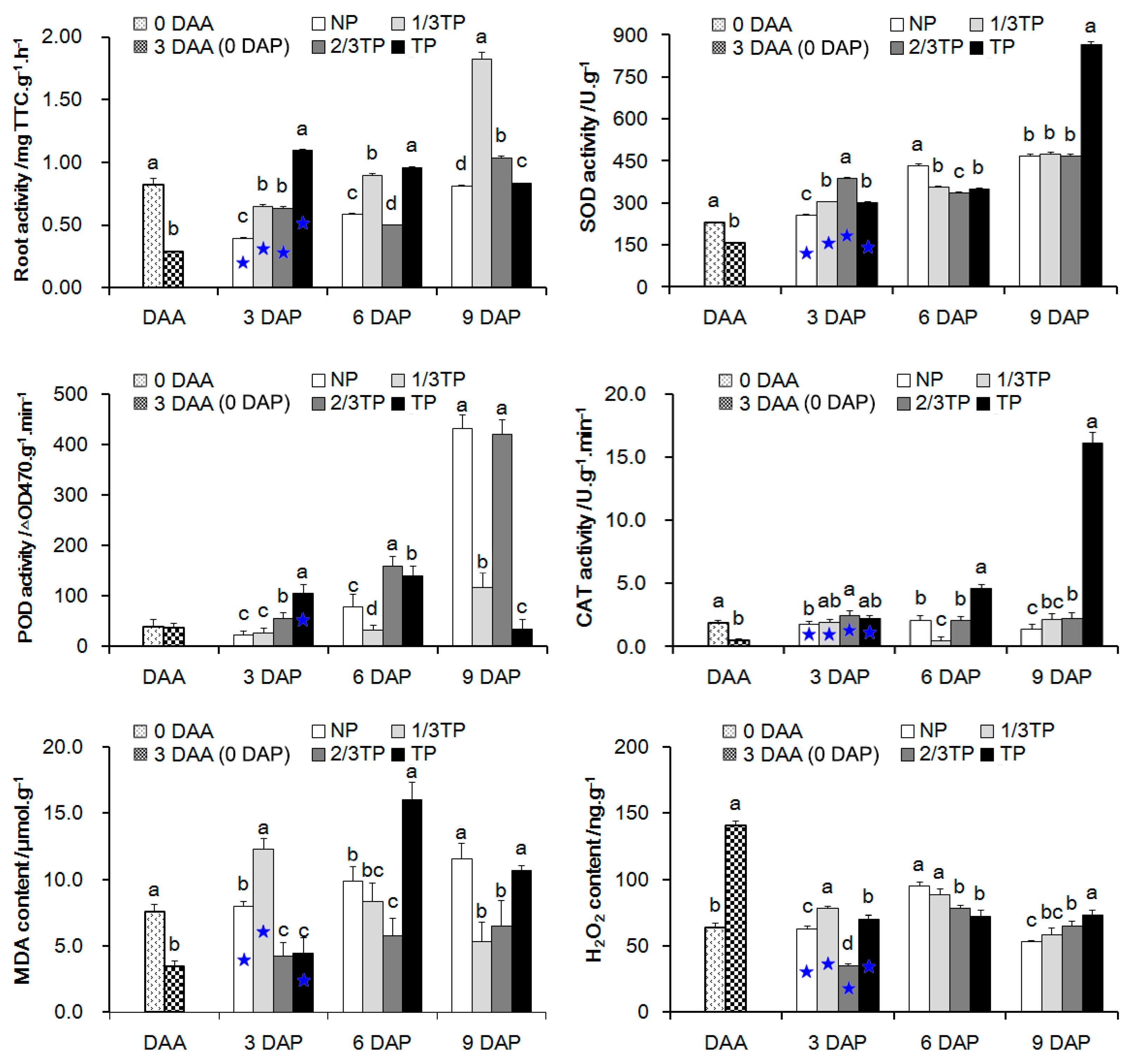
2.4. Physiological Response to Replant Disease Stresses in R. glutinosa
3. Discussion
3.1. Replant Disease Promotes Proliferation of Fusarium oxysporum in R. glutinosa Rhizospheres Soil
3.2. NB-LRRs Failed to Respond Timely and Effectively to Pathogenic Fusarium oxysporum in Replanted R. glutinosa
3.3. Lower Level of SA Biosynthesis Stimulated by Fusarium oxysporum Might Be Cloesly Related to the Formation of R. glutinosa Replant Disease
4. Materials and Methods
4.1. Plant Growth and Treatments
4.2. The Collection of Fresh Root and Rhizosphere Soil Samples
4.3. The Extraction of Soil DNA and Its Method Comparison
4.4. Absolute Quantification of Pseudomonas spp. and Fusarium oxysporum
4.4.1. Construction of the Recombinant Plasmid
4.4.2. Establishment of Standard Curve
4.4.3. Determination of AQ-PCR
4.5. qRT-PCR Analysis of NB-LRRs
4.6. Measurement of ABA, SA, ET and JA
4.7. Measurement of Root Activity and the Physiological Index
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DAA | Days after acclimatization |
| DAP | Days after planting |
| NP | Soil that was never planted with R. glutinosa for at least 10 years |
| TP | Soil that was consecutively planted with R. glutinosa in the same soils for three years |
| FO | Fusarium oxysporum f.sp. R. glutinosa |
| PS | Pseudomonas spp. |
| PR | Pathogenesis-related |
| PTI | Pathogen-associated molecular patterns (PAMP)-triggered immunity |
| ETI | Effector-triggered immunity |
| PRRs | Pattern recognition receptors |
| NB-LRR | Nucleotide-binding-leucine-rich repeat |
| NB-ARC | Nucleotide-binding adaptor shared by APAF-1, R proteins, and CED-4 |
| TIR | Toll/interleukin-1 receptor |
| CC | Coiled-coil |
| NPR1 | Non-expressor of pathogenesis-related proteins 1 |
| EDS1 | Enhanced disease susceptibility 1 |
| NDR1 | Non-race-specific disease resistance 1 |
| R1A | Resistance in linkage group 1A |
| R1B | Resistance in linkage group 1B |
| RPS2 | Resistance to Pseudomonas syringae 2 |
| TAO1 | Target of AvrB operation 1 |
| RPW8 | Resistance to Powdery mildew 8 |
| MPK4 | Mitogen-activated protein kinase 4 |
| ELISA | Enzyme-linked immunosorbent assay |
| SA | Salicylic acid |
| JA | Jasmonic acid |
| ET | Ethylene |
| ABA | Abscisic acid |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| CAT | Catalase |
| MDA | Malondialdehyde |
| H2O2 | Hydrogen peroxide |
| AQ-PCR | Absolute quantification PCR |
| IPTG | Isopropyl-l-d-thiogalactopyranoside |
| X-Gal | 5-Bromo-4-chloro-3-indolyl β-d-galactoside |
| Amp | Ampicillin |
| PBS | Phosphate-buffered saline |
| TTC | Triphenyl tetrazolium chloride |
References
- Zhang, Z.Y.; Yin, W.J.; Li, J.; Du, J.F.; Yang, Y.H.; Chen, X.J.; Lin, W.X. Physio-ecological properties of continuous cropping Rehmannia glutinosa. Chin. J. Plant Ecol. 2010, 34, 547–554. [Google Scholar]
- Yang, M.; Zhang, X.D.; Xu, Y.G.; Mei, X.Y.; Jiang, B.B.; Liao, J.J.; Yin, Z.B.; Zheng, J.F.; Zhao, Z.; Fan, L.M.; et al. Autotoxic ginsenosides in the rhizosphere contribute to the replant failure of Panax notoginseng. PLoS ONE 2015, 10, e0118555. [Google Scholar] [CrossRef] [PubMed]
- Rumberger, A.; Merwin, I.A.; Thies, J.E. Microbial community development in the rhizosphere of apple trees at a replant disease site. Soil Biol. Biochem. 2007, 39, 1645–1654. [Google Scholar] [CrossRef]
- Yang, J.I.; Ruegger, P.M.; McKenry, M.V.; Becker, J.O.; Borneman, J. Correlations between root-associated microorganisms and peach replant disease symptoms in a California soil. PLoS ONE 2012, 7, e46420. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Liu, Q.Z.; Chen, P. Effect of long-term continuous cropping of strawberry on soil bacterial community structure and diversity. J. Integr. Agric. 2018, 17, 2570–2582. [Google Scholar] [CrossRef]
- Bai, L.; Cui, J.Q.; Jie, W.G.; Cai, B.Y. Analysis of the community compositions of rhizosphere fungi in soybeans continuous cropping fields. Microbiol. Res. 2015, 180, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.G.; Li, M.J.; Zhang, B.; Wang, F.J.; Gu, L.; Feng, F.J.; Liu, H.Y.; Zhang, Z.Y. Review on catastrophe mechanism of medicinal plant and its rhizosphere microecosystem mediated by consecutive monoculture. Mod. Chin. Med. 2016, 18, 239–245. [Google Scholar]
- Zhang, B.; Li, X.Z.; Wang, F.Q.; Li, M.J.; Zhang, J.Y.; Gu, L.; Zhang, L.J.; Tu, W.Q.; Zhang, Z.Y. Assaying the potential autotoxins and microbial community associated with Rehmannia glutinosa replant problems based on its ‘autotoxic circle’. Plant Soil 2016, 407, 307–322. [Google Scholar] [CrossRef]
- Wen, X.S.; Yang, S.L.; Wei, J.H.; Zheng, J.H. Textual research on planting history of Rehmannia glutinosa and its cultivated varieties. Chin. Tradit. Herb. Drugs 2002, 33, 946–949. [Google Scholar]
- Wu, L.K.; Wang, J.Y.; Huang, W.M.; Wu, H.M.; Chen, J.; Yang, Y.Q.; Zhang, Z.Y.; Lin, W.X. Plant-microbe rhizosphere interactions mediated by Rehmannia glutinosa root exudates under consecutive monoculture. Sci. Rep. 2015, 5, 15871. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Lin, W.X. Continuous cropping obstacle and allelopathic autotoxicity of medicinal plants. Chin. J. Eco-Agric. 2009, 17, 189–196. [Google Scholar] [CrossRef]
- Kuijken, R.C.P.; Snel, J.F.H.; Heddes, M.H.; Bouwmeester, H.J.; Marcelis, L.F.M. The importance of a sterile rhizosphere when phenotyping for root exudation. Plant Soil 2015, 387, 131–142. [Google Scholar] [CrossRef]
- Wu, L.K.; Li, Z.F.; Li, J.; Khan, M.A.; Huang, W.M.; Zhang, Z.Y.; Lin, W.X. Assessment of shifts in microbial community structure and catabolic diversity in response to Rehmannia glutinosa monoculture. Appl. Soil. Ecol. 2013, 67, 1–9. [Google Scholar] [CrossRef]
- Barto, E.K.; Cipollini, D. Half-lives and field soil concentrations of Alliaria petiolata secondary metabolites. Chemosphere 2009, 76, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Pan, L.P.; Li, H.H. Isolation, identification and characterization of soil microbes which degrade phenolic allelochemicals. J. Appl. Microbiol. 2010, 108, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Li, X.G.; Ding, C.F.; Hua, K.; Zhang, T.L.; Zhang, Y.N.; Zhao, L.; Yang, Y.R.; Liu, J.G.; Wang, X.X. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy. Soil Boil. Biochem. 2014, 78, 149–159. [Google Scholar] [CrossRef]
- Venturi, V.; Keel, C. Signaling in the rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Saleem, M.; Pervaiz, Z.H.; Traw, M.B. Theories, mechanisms and patterns of microbiome species coexistence in an era of climate change. In Microbiome Community Ecology; Springer: Cham, Switzerland, 2015; pp. 13–53. [Google Scholar]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver, L.T.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Lebeis, S.L. Greater than the sum of their parts: Characterizing plant microbiomes at the community-level. Curr. Opin. Plant Biol. 2015, 24, 82–86. [Google Scholar] [CrossRef]
- Biere, A.; Goverse, A. Plant-mediated systemic interactions between pathogens, parasitic nematodes, and herbivores above- and belowground. Annu. Rev. Phytopathol. 2016, 54, 499–527. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.S.; Zhen, W.C.; Qi, Y.Z.; Liu, X.J.; Yin, B.Z. Coordinated effects of root autotoxic substances and Fusarium oxysporum Schl. f. sp. fragariae on the growth and replant disease of strawberry. Front. Agric. China 2009, 3, 34. [Google Scholar] [CrossRef]
- Mazzola, M.; Manici, L.M. Apple replant disease: Role of microbial ecology in cause and control. Annu. Rev. Phytopathol. 2012, 50, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Spath, M.; Insam, H.; Peintner, U.; Kelderer, M.; Kuhnert, R.; Franke-Whittle, I. Linking soil biotic and abiotic factors to apple replant disease: A greenhouse approach. J. Phytopathol. 2015, 163, 287–299. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Chen, H.; Yang, Y.H.; Chen, T.; Lin, R.Y.; Chen, X.J.; Lin, W.X. Effects of continuous cropping on bacterial community diversity in rhizosphere soil of Rehmannia glutinosa. Chin. J. Appl. Ecol. 2010, 21, 2843–2848. [Google Scholar]
- Zhang, Z.Y.; Lin, W.X.; Yang, Y.H.; Chen, H.; Chen, X.J. Effects of consecutively monocultured Rehmannia glutinosa L. on diversity of fungal community in rhizospheric soil. Agric. Sci. China 2011, 10, 1374–1384. [Google Scholar] [CrossRef]
- Li, Z.F.; Yang, Y.Q.; Wu, L.K.; Shu, Y.; Zhao, Y.P.; Huang, W.M.; Zhang, Z.Y.; Lin, W.X. Isolation of highly pathogenic pathogens and identification of formae speciales of Rehmannia glutinosa L. Chin. J. Eco-Agric. 2013, 21, 1426–1433. [Google Scholar] [CrossRef]
- Wu, L.K.; Huang, W.M.; Wang, J.Y.; Wu, H.M.; Chen, J.; Qin, X.J.; Zhang, Z.Y.; Lin, W.X. Diversity analysis of rhizosphere microflora of wild R. glutinosa grown in monocropping for different years. Acta Agron. Sin. 2015, 41, 308–317. [Google Scholar] [CrossRef]
- Wu, L.K.; Wang, J.Y.; Wu, H.M.; Chen, J.; Xiao, Z.G.; Qin, X.J.; Zhang, Z.Y.; Lin, W.X. Comparative metagenomic analysis of rhizosphere microbial community composition and functional potentials under Rehmannia glutinosa consecutive monoculture. Int. J. Mol. Sci. 2018, 19, 2394. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, R.; Luu, V.T.; Weinhold, A.; Goldberg, J.; Oh, Y.; Baldwin, I.T. Native root-associated bacteria rescue a plant from a sudden-wilt disease that emerged during continuous cropping. Proc. Natl. Acad. Sci. USA 2015, 112, 5013–5020. [Google Scholar] [CrossRef] [PubMed]
- Tena, G. Immunity: NLR population control. Nat. Plants 2016, 2, 16026. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.N.; Costa Alves, G.S.; van Sluys, M.A. Plant immunity: Unravelling the complexity of plant responses to biotic stresses. Ann. Bot. 2017, 119, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stael, S.; Kmiecik, P.; Willems, P.; Kelen, K.V.D.; Coll, N.S.; Teige, M.; van Breusegem, F. Plant innate immunity-sunny side up? Trends Plant Sci. 2015, 20, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nature 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Kapo, P.; Devendrakumar, K.T.; Li, X. Plant NLRs: From discovery to application. Plant Sci. 2019, 279, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Ji, H.; Amirullah, A.; Traw, M.B. Pseudomonas syringae pv. tomato DC3000 growth in multiple gene knockouts predicts interactions among hormonal, biotic and abiotic stress responses. Eur. J. Plant Pathol. 2017, 149, 779–786. [Google Scholar] [CrossRef]
- Xie, Z.M.; Yang, C.Y.; Chen, A.G.; Li, M.J.; Gu, L.; Zhang, J.Y.; Feng, F.J.; Zhang, B.; Chen, S.Q.; Zhang, Z.Y. Identification and expression analysis of leucine-rich repeat receptor-like kinase family reveals the roles of resistance proteins during formation of replant disease in Rehmannia glutinosa Libosch. Int. J. Agric. Biol. 2019. [Google Scholar] [CrossRef]
- Chen, A.G.; Gu, L.; Li, M.J.; Xu, N.; Feng, F.J.; Liu, G.L.; Zhang, B.; Gong, D.P.; Zhang, J.Y.; Liu, H.Y.; et al. Identifcation of Rehmannia glutinosa L. NB-ARC family proteins and their typical changes under consecutive monoculture stress. Acta Physiol. Plant. 2018, 40, 95. [Google Scholar] [CrossRef]
- Andolfo, G.; Ercolano, M.R. Plant innate immunity multicomponent model. Front. Plant Sci. 2015, 6, 987. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, A.M.; Berens, M.L.; Tsuda, K.; Argueso, C.T. Towards engineering of hormonal crosstalk in plant immunity. Curr. Opin. Plant Biol. 2017, 38, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Derksen, H.; Rampitsch, C.; Daayf, F. Signaling cross-talk in plant disease resistance. Plant Sci. 2013, 207, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Lievens, L.; Pollier, J.; Goossens, A.; Beyaert, R.; Staal, J. Abscisic acid as pathogen effector and immune regulator. Front. Plant Sci. 2017, 8, 587. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, K.J.; Lee, Y.S. Isolation and characterization of the salicylic acid induced gene in Rehmannia glutinosa by differential display. Mycobiology 2002, 30, 88–92. [Google Scholar] [CrossRef]
- Wang, F.Q.; Zhi, J.Y.; Zhang, Z.Y.; Wang, L.N.; Suo, Y.F.; Xie, C.X.; Li, M.J.; Zhang, B.; Du, J.F.; Gu, L.; et al. Transcriptome analysis of salicylic acid treatment in Rehmannia glutinosa hairy roots using RNA-seq technique for identification of genes involved in acteoside biosynthesis. Front. Plant Sci. 2017, 8, 787. [Google Scholar] [CrossRef] [PubMed]
- Li, M.J.; Yang, Y.H.; Feng, F.J.; Zhang, B.; Chen, S.Q.; Yang, C.Y.; Gu, L.; Wang, F.Q.; Zhang, J.Y.; Chen, A.G.; et al. Differential proteomic analysis of replanted Rehmannia glutinosa roots by iTRAQ reveals molecular mechanisms for formation of replant disease. BMC Plant Biol. 2017, 17, 116. [Google Scholar] [CrossRef]
- Wu, L.K.; Chen, J.; Khan, M.U.; Wang, J.Y.; Wu, H.M.; Xiao, Z.G.; Zhang, Z.Y.; Lin, W.X. Rhizosphere fungal community dynamics associated with Rehmannia glutinosa replant disease in a consecutive monoculture regime. Phytopathology 2018, 108, 1493–1500. [Google Scholar] [CrossRef]
- Wu, L.K.; Chen, J.; Xiao, Z.G.; Zhu, X.C.; Wang, J.Y.; Wu, H.M.; Wu, Y.H.; Zhang, Z.Y.; Lin, W.X. Barcoded pyrosequencing reveals a shift in the bacterial community in the rhizosphere and rhizoplane of Rehmannia glutinosa under consecutive monoculture. Int. J. Mol. Sci. 2018, 19, 850. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; Voort, M.V.D.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Bouizgarne, B.; Brault, M.; Pennarun, A.M.; Rona, J.P.; Ouhdouch, Y.; Hadrami, E.I.; Bouteau, F. Electrophysiological responses to fusaric acid of root hairs from seedlings of date palm-susceptible and -resistant to Fusarium oxysporum f. sp. albedinis. J. Phytopathol. 2004, 152, 321–324. [Google Scholar] [CrossRef]
- Ye, S.F.; Yu, J.Q.; Peng, Y.H.; Zheng, J.H.; Zou, L.Y. Incidence of Fusarium wilt in Cucumis sativus L. is promoted by cinnamic acid, an autotoxin in root exudates. Plant Soil 2004, 263, 143–150. [Google Scholar] [CrossRef]
- Scott, G.; Almasrahi, A.; Mansoorkhani, F.M.; Rupar, M.; Dickinson, M.; Shama, G. Hormetic UV-C seed treatments for the control of tomato diseases. Plant Pathol. 2019, 68, 700–707. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Bi, D.; Cheng, Y.T.; Li, X.; Zhang, Y. SRFR1 negatively regulates plant NB-LRR resistance protein accumulation to prevent autoimmunity. PLoS Pathog. 2010, 6, e1001111. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.H.; Xu, F.; Zhang, Y.X.; Cheng, Y.T.; Wiermer, M.; Li, X.; Zhang, Y.L. Arabidopsis resistance protein SNC1 activates immune response through association with a transcriptional corepressor. Proc. Natl. Acad. Sci. USA 2010, 107, 13960–13965. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Lin, X.L.; Kong, X.X.; Qu, G.P.; Cai, B.; Lee, J.Y.; Jin, J.B. SIZ1-mediated SUMOylation of TPR1 suppresses plant Immunity in Arabidopsis. Mol. Plant 2019, 12, 215–228. [Google Scholar] [CrossRef]
- Richard, M.M.S.; Takken, F.L.W. Plant autoimmunity: When good things go bad. Curr. Biol. 2017, 27, 361–363. [Google Scholar] [CrossRef]
- Wang, J.L.; Grubb, L.E.; Wang, J.Y.; Liang, X.X.; Li, L.; Gao, C.L.; Ma, M.M.; Feng, F.; Li, M.; Li, L.; et al. A regulatory module controlling homeostasis of a plant immune kinase. Mol. Cell 2018, 69, 493–504. [Google Scholar] [CrossRef]
- Cai, X.Z.; Xu, Q.F.; Wang, C.C.; Zheng, Z. Development of a virus-induced gene-silencing system for functional analysis of the RPS2-dependent resistance signalling pathways in Arabidopsis. Plant Mol. Biol. 2006, 62, 223–232. [Google Scholar] [CrossRef]
- Eitas, T.K.; Nimchuk, Z.L.; Dangl, J.L. Arabidopsis TAO1 is a TIR-NB-LRR protein that contributes to disease resistance induced by the Pseudomonas syringae effector AvrB. Proc. Natl. Acad. Sci. USA 2008, 105, 6475–6480. [Google Scholar] [CrossRef]
- Slootweg, E.; Roosien, J.; Spiridon, L.N.; Petrescu, A.J.; Tameling, W.; Joosten, M.; Pomp, R.; van Schaik, C.; Dees, R.; Borst, J.W.; et al. Nucleocytoplasmic distribution is required for activation of resistance by the potato NB-LRR receptor Rx1 and is balanced by its functional domains. Plant Cell 2010, 22, 4195–4215. [Google Scholar] [CrossRef] [PubMed]
- Castel, B.; Ngou, P.M.; Cevik, V.; Redkar, A.; Kim, D.S.; Yang, Y.; Ding, P.T.; Jones, J.D.G. Diverse NLR immune receptors activate defence via the RPW8-NLR NRG1. New Phytol. 2019, 222, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef] [PubMed]
- An, C.F.; Mou, Z.L. Salicylic acid and its function in plant immunity. J. Integr. Plant Biol. 2011, 53, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.R.; Jaleel, H. Jasmonates counter plant stress: A review. Environ. Exp. Bot. 2015, 115, 49–57. [Google Scholar] [CrossRef]
- Wu, L.M.; Huang, Z.Y.; Li, X.; Ma, L.M.; Gu, Q.; Wu, H.J.; Liu, J.; Borriss, R.; Wu, Z.; Gao, X.W. Stomatal closure and SA-, JA/ET-signaling pathways are essential for Bacillus amyloliquefaciens FZB42 to restrict leaf disease caused by Phytophthora nicotianae in Nicotiana benthamiana. Front. Microbiol. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Adie, B.A.; Pérez-Pérez, J.; Pérez-Pérez, M.M.; Godoy, M.; Sánchez-Serrano, J.J.; Schmelz, E.A.; Solano, R. ABA is an essential signal for plant resistance to pathogens affecting JA biosynthesis and the activation of defense in Arabidopsis. Plant Cell 2007, 19, 1665–1681. [Google Scholar] [CrossRef]
- Niu, M.M.; Li, J.; Du, J.F.; Yin, W.J.; Yang, Y.H.; Chen, X.J.; Zhang, Z.Y. Changes in source-sink relationship of photosynthate in Rehmannia glutinosa Libosch. and their relations with continuous cropping obstacle. Chin. J. Ecol. 2011, 30, 248–254. [Google Scholar]
- Coyne, K.J.; Handy, S.M.; Demir, E.; Whereat, E.B.; Hutchins, D.A.; Portune, K.J.; Doblin, M.A.; Cary, C. Improved quantitative real-time PCR assays for enumeration of harmful algal species in field samples using an exogenous DNA reference standard. Limnol. Oceanogr. Methods 2005, 3, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Lou, J.; Wang, H.Z.; Wu, L.S.; Xu, J.M. Use of an improved high-throughput absolute abundance quantification method to characterize soil bacterial community and dynamics. Sci. Total Environ. 2018, 633, 360–371. [Google Scholar] [CrossRef]
- Shirima, R.R.; Maeda, D.G.; Kanju, E.; Ceasar, G.; Tibazarwa, F.; Legg, J.P. Absolute quantification of cassava brown streak virus mRNA by real-time qPCR. J. Virol. Methods 2017, 245, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, G.; Yi, G.X.; Wang, B.M.; Deng, A.X.; Nan, T.G.; Li, Z.H.; Li, Q.X. Comparison between conventional indirect competitive enzyme-linked immunosorbent assay (icELISA) and simplified icELISA for small molecules. Anal. Chim. Acta 2006, 571, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, B.; Du, M.W.; Eneji, A.E.; Wang, B.M.; Duan, L.S.; Li, Z.H.; Tian, X.L. Mechanism of phytohormone involvement in feedback regulation of cotton leaf senescence induced by potassium deficiency. J. Exp. Bot. 2012, 63, 5887–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Huang, G.; Bian, X.; Zhao, Q. Effects of root interaction and nitrogen fertilization on the chlorophyll content, root activity, photosynthetic characteristics of intercropped soybean and microbial quantity in the rhizosphere. Plant Soil Environ. 2013, 59, 80–88. [Google Scholar] [CrossRef]
- Deenamo, N.; Kuyyogsuy, A.; Khompatara, K.; Chanwun, T.; Ekchaweng, K.; Churngchow, N. Salicylic acid induces resistance in rubber tree against Phytophthora palmivora. Int. J. Mol. Sci. 2018, 19, 1883. [Google Scholar] [CrossRef] [PubMed]
- Chakrabrty, D.; Datta, S.K. Micropropagation of gerbera: Lipid peroxidation and antioxidant enzyme activities during acclimatization process. Acta Physiol. Plant. 2008, 30, 325–331. [Google Scholar] [CrossRef]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. TM4 microarray software suite. Methods Enzymol. 2006, 411, 134–193. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ∆FO/PS | ∆PS | ∆FO | |
|---|---|---|---|
| Levels of replant disease stresses | 0.7918 ** | −0.5611 | 0.6809 * |
| PS | FO | |
|---|---|---|
| FO/PS | −0.2488 | 0.9208 ** |
| PS | 0.1331 |
| RgNB1 | RgNB2 | RgNB3 | RgNB4 | RgNB5 | RgNB6 | RgNB7 | |
| PS | −0.1156 | 0.0187 | 0.6833 | −0.1978 | 0.5612 | −0.0038 | −0.3908 |
| FO | 0.9737 | 0.9344 | 0.4558 | −0.3259 | −0.4752 | 0.9219 | 0.9963 ** |
| RgNB8 | RgNB9 | RgNB10 | RgNB11 | RgNB12 | RgNB13 | RgNB14 | |
| PS | −0.2584 | 0.0342 | −0.1756 | 0.1503 | −0.4000 | −0.2856 | 0.7931 |
| FO | 0.9950 ** | 0.8935 | 0.5160 | 0.8193 | 0.9973 ** | 0.9984 ** | −0.7543 |
| RgNB15 | RgNB16 | RgNB17 | RgNB18 | RgNB19 | RgNB20 | RgNB21 | |
| PS | −0.3292 | −0.1421 | 0.0665 | −0.4500 | −0.0412 | −0.2643 | −0.3687 |
| FO | 0.0526 | 0.9794 * | −0.4775 | 0.8240 | 0.9452 | 0.9970 ** | 0.2326 |
| RgNB22 | RgNB23 | RgNB24 | RgNB25 | RgNB26 | RgNB27 | RgNB28 | |
| PS | −0.4505 | −0.4356 | −0.1776 | −0.3769 | −0.1168 | −0.0632 | −0.2509 |
| FO | 0.5924 | 0.8712 | 0.9858 * | 0.9845 * | −0.6370 | 0.9564 * | 0.9953 ** |
| RgNB29 | RgNB30 | RgNB31 | RgNB32 | RgNB33 | RgNB34 | RgNB35 | |
| PS | 0.4527 | −0.3214 | −0.3369 | −0.0666 | −0.4524 | 0.6505 | 0.3434 |
| FO | −0.8157 | 0.9787 * | 0.9965 ** | −0.3699 | −0.5155 | −0.7435 | −0.6044 |
| ∆FO | ∆JA | ∆ABA | ∆ET | ∆SA | |
|---|---|---|---|---|---|
| ∆PS | −0.7327 | 0.1328 | 0.0254 | 0.1123 | 0.2366 |
| ∆FO | −0.6524 | −0.5981 | −0.6560 | −0.7475 * | |
| ∆JA | 0.9752 ** | 0.9839 ** | 0.9182 ** | ||
| ∆ABA | 0.9650 ** | 0.9543 ** | |||
| ∆ET | 0.9009 ** |
| ∆Root Activity | ∆SOD | ∆POD | ∆CAT | ∆H2O2 | ∆MDA | ||
|---|---|---|---|---|---|---|---|
| 0~3 DAP | ∆PS | 0.0357 | 0.2189 | 0.3088 | −0.1421 | −0.3443 | −0.3313 |
| ∆FO | 0.4389 | −0.1874 | 0.2671 | 0.4416 | 0.1959 | −0.1724 | |
| 0~6 DAP | ∆PS | 0.1759 | −0.0713 | −0.0387 | −0.2458 | 0.1394 | −0.2020 |
| ∆FO | 0.2250 | 0.1574 | 0.2263 | 0.7464 * | −0.5732 | 0.7316 * | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, A.; Gu, L.; Xu, N.; Feng, F.; Chen, D.; Yang, C.; Zhang, B.; Li, M.; Zhang, Z. NB-LRRs Not Responding Consecutively to Fusarium oxysporum Proliferation Caused Replant Disease Formation of Rehmannia glutinosa. Int. J. Mol. Sci. 2019, 20, 3203. https://doi.org/10.3390/ijms20133203
Chen A, Gu L, Xu N, Feng F, Chen D, Yang C, Zhang B, Li M, Zhang Z. NB-LRRs Not Responding Consecutively to Fusarium oxysporum Proliferation Caused Replant Disease Formation of Rehmannia glutinosa. International Journal of Molecular Sciences. 2019; 20(13):3203. https://doi.org/10.3390/ijms20133203
Chicago/Turabian StyleChen, Aiguo, Li Gu, Na Xu, Fajie Feng, Dexin Chen, Chuyun Yang, Bao Zhang, Mingjie Li, and Zhongyi Zhang. 2019. "NB-LRRs Not Responding Consecutively to Fusarium oxysporum Proliferation Caused Replant Disease Formation of Rehmannia glutinosa" International Journal of Molecular Sciences 20, no. 13: 3203. https://doi.org/10.3390/ijms20133203
APA StyleChen, A., Gu, L., Xu, N., Feng, F., Chen, D., Yang, C., Zhang, B., Li, M., & Zhang, Z. (2019). NB-LRRs Not Responding Consecutively to Fusarium oxysporum Proliferation Caused Replant Disease Formation of Rehmannia glutinosa. International Journal of Molecular Sciences, 20(13), 3203. https://doi.org/10.3390/ijms20133203