Mangifera indica L. Leaf Extract Induces Adiponectin and Regulates Adipogenesis



 ,
, 

 and
and 
Abstract
:1. Introduction
2. Results
2.1. Polyphenols Determination
2.2. UPLC-MS/MS
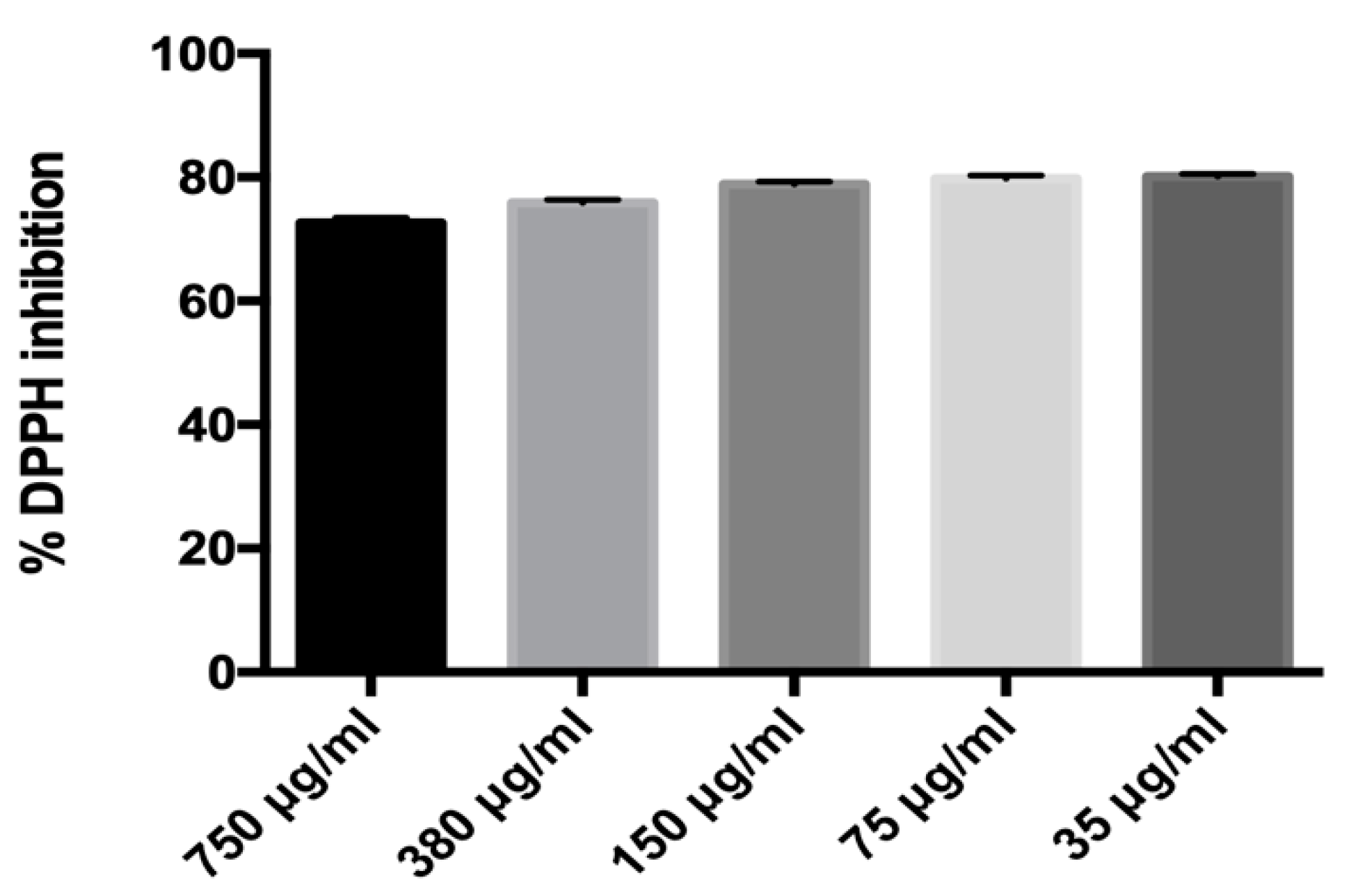
2.3. Radical Scavenging Activity of MLE
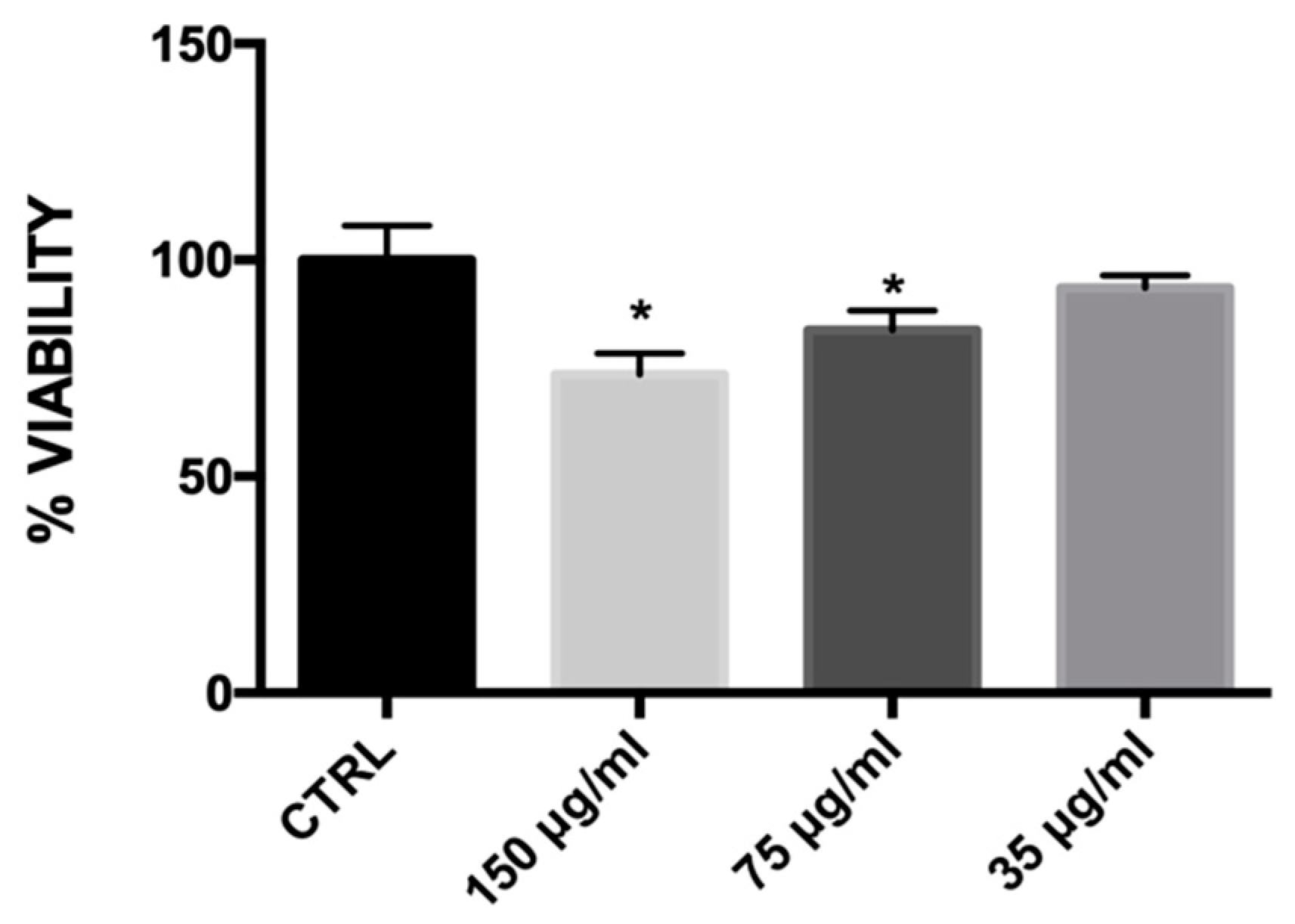
2.4. Effect of MLE on Cell Viability
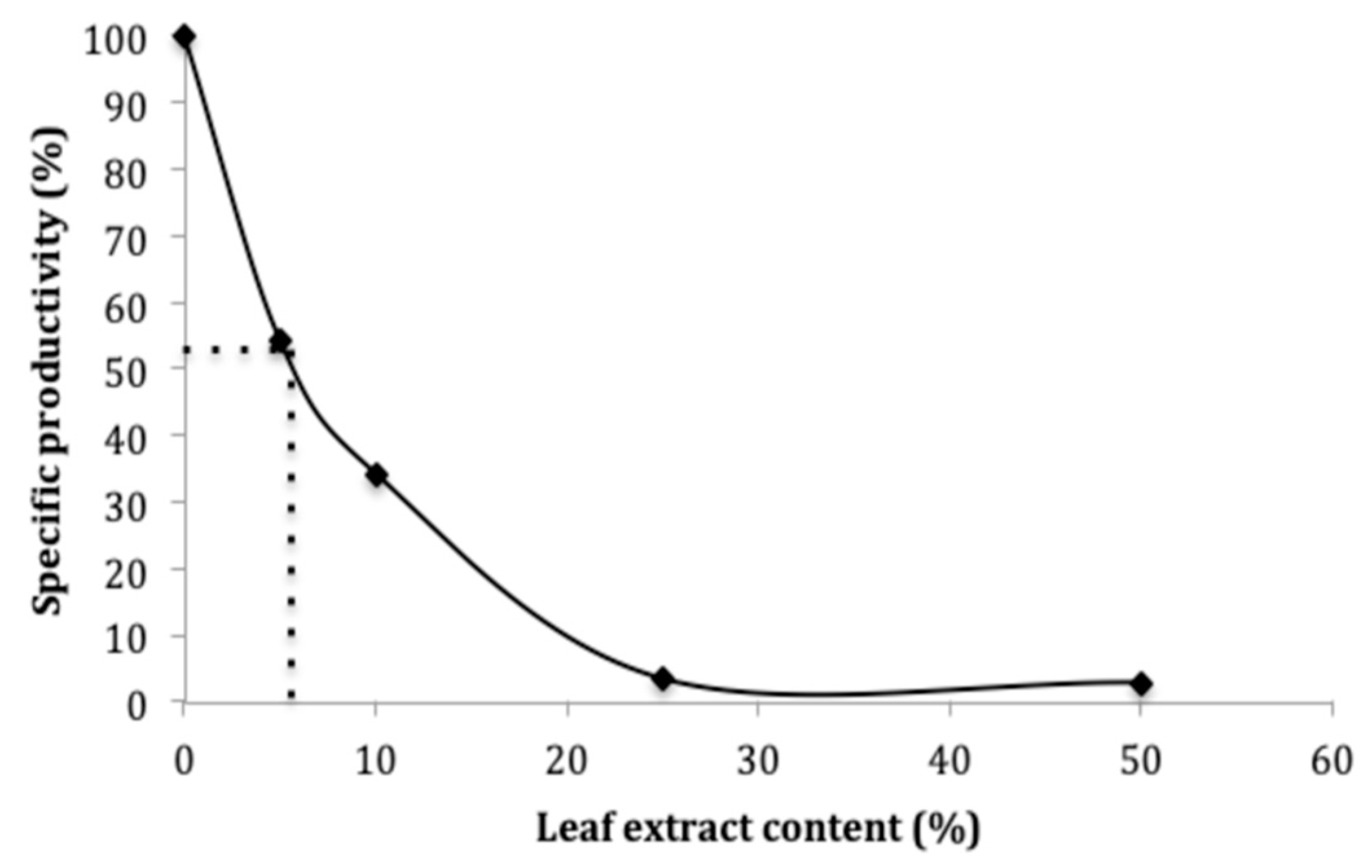
2.5. Inhibition of α-Glucosidase
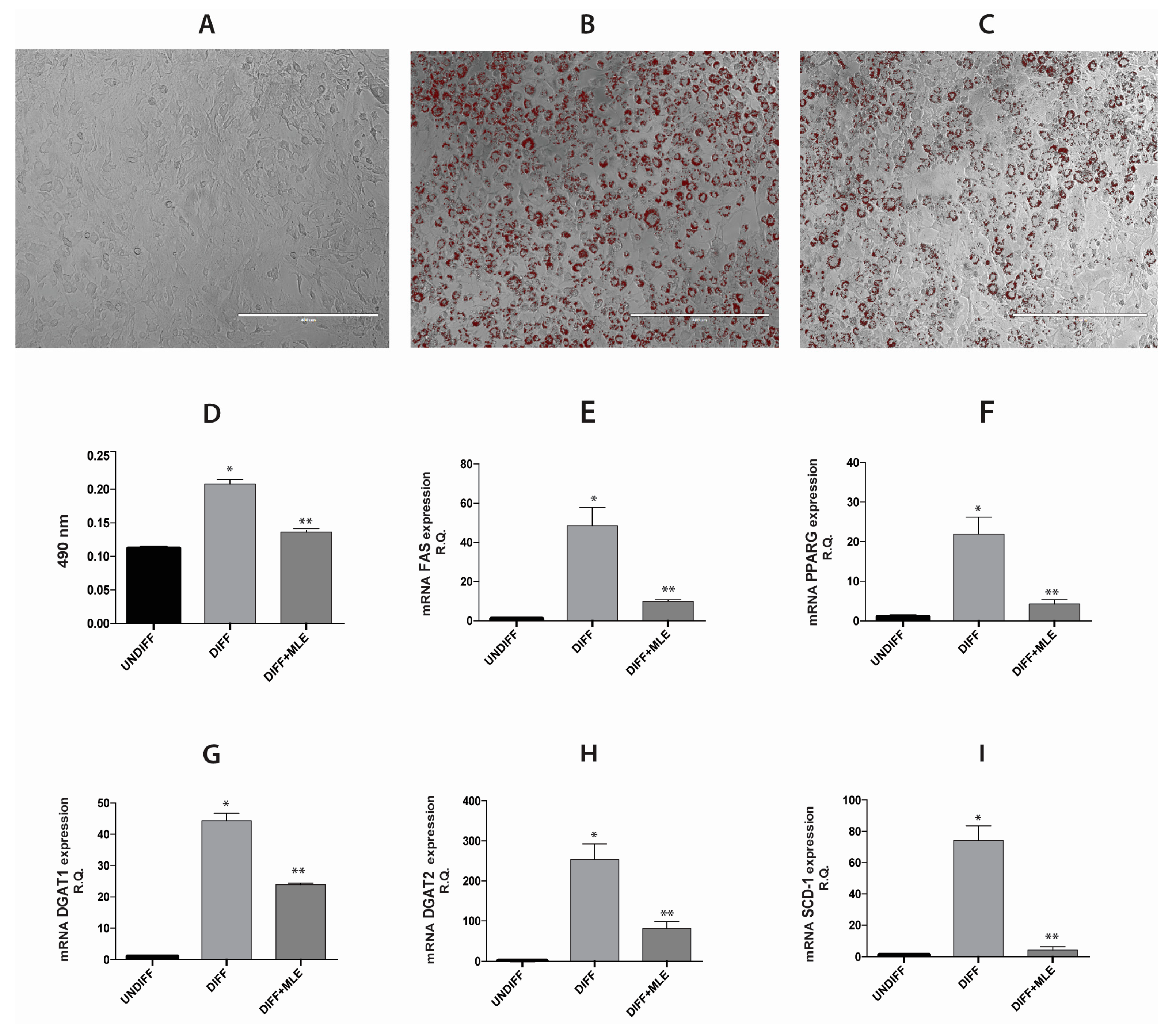
2.6. Analysis of Adipogenic Differentiation in 3T3-L1 Cells
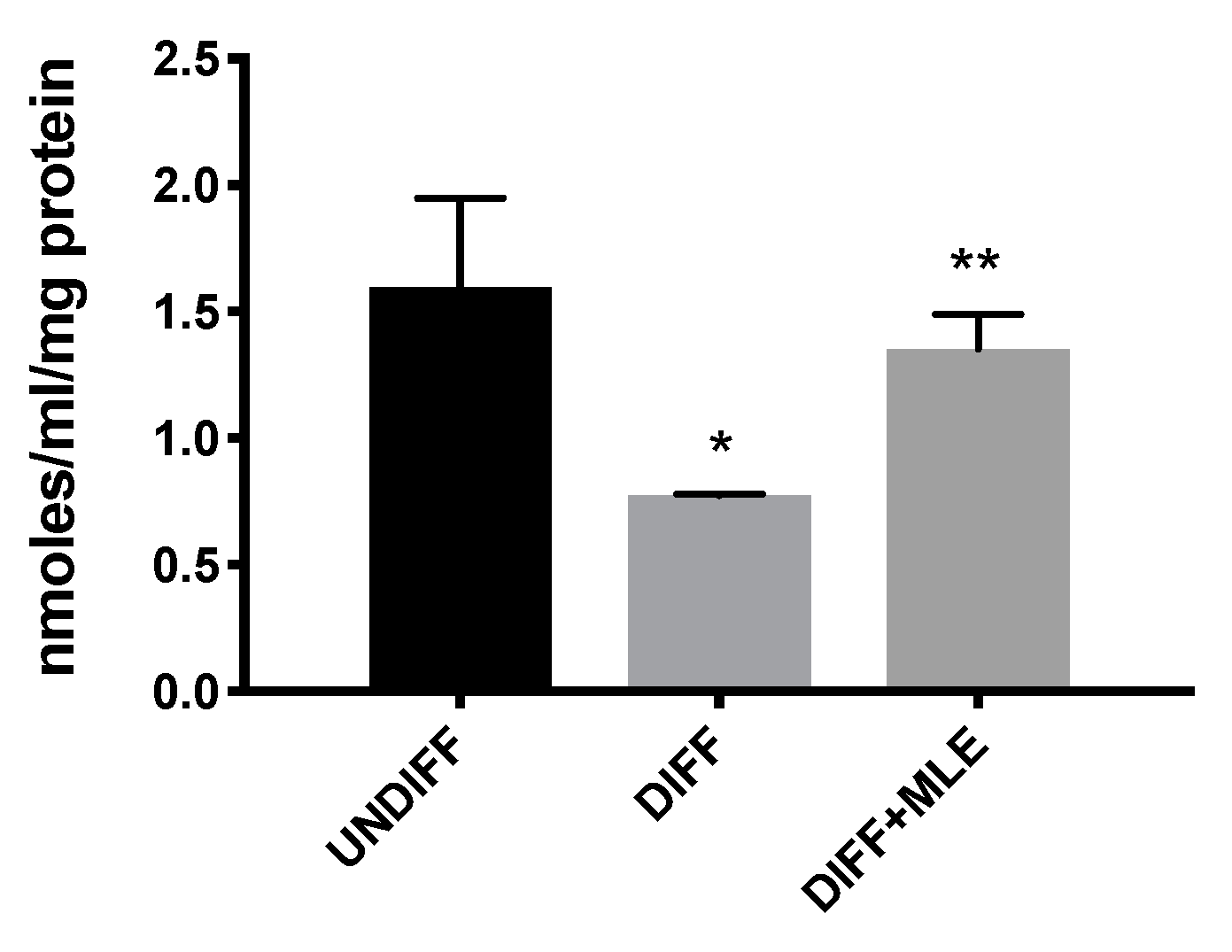
2.7. MLE Restores Glutathione Resources after Differentiation
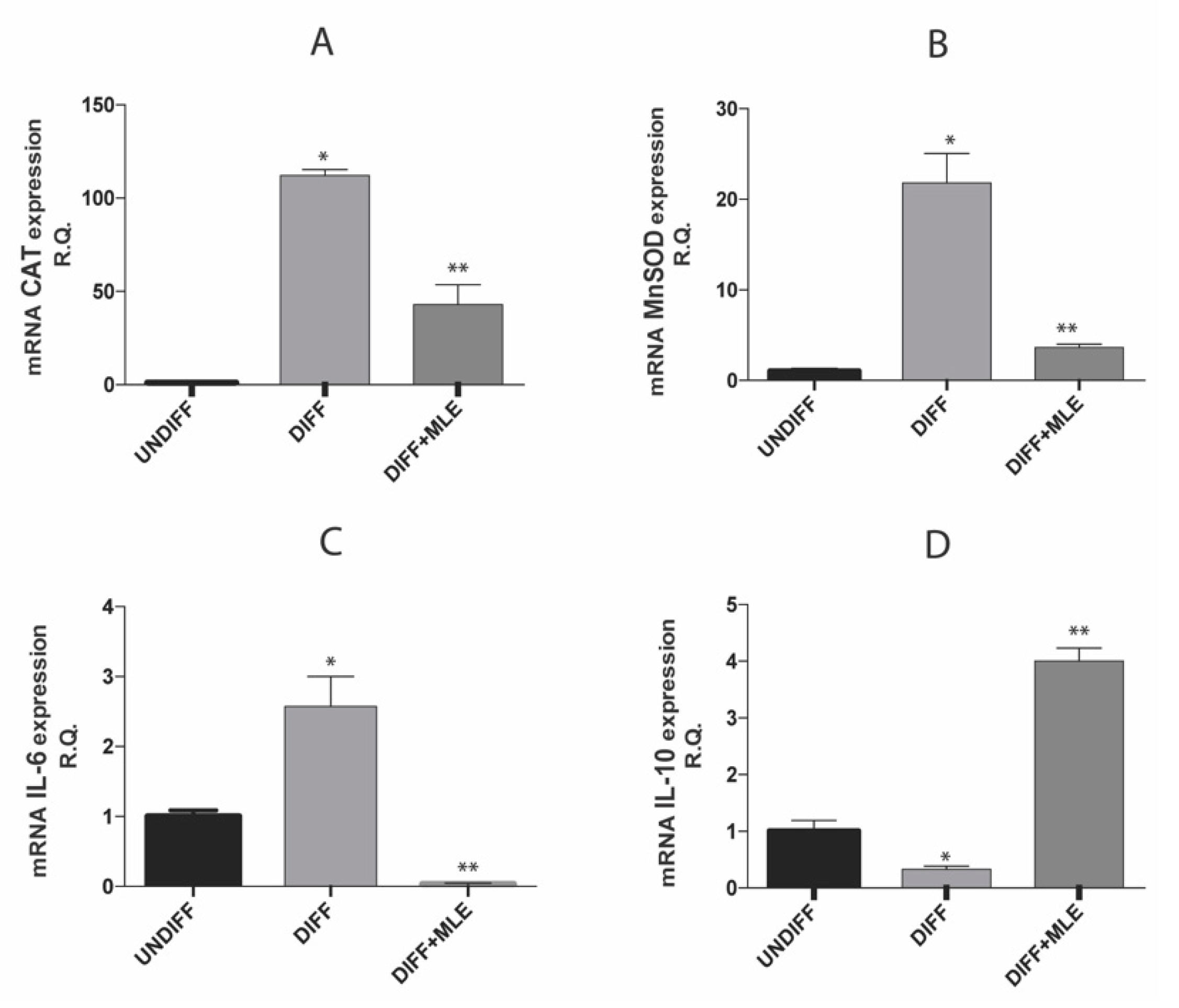
2.8. MLE Effects on Oxidative Stress and Inflammatory Markers Expression
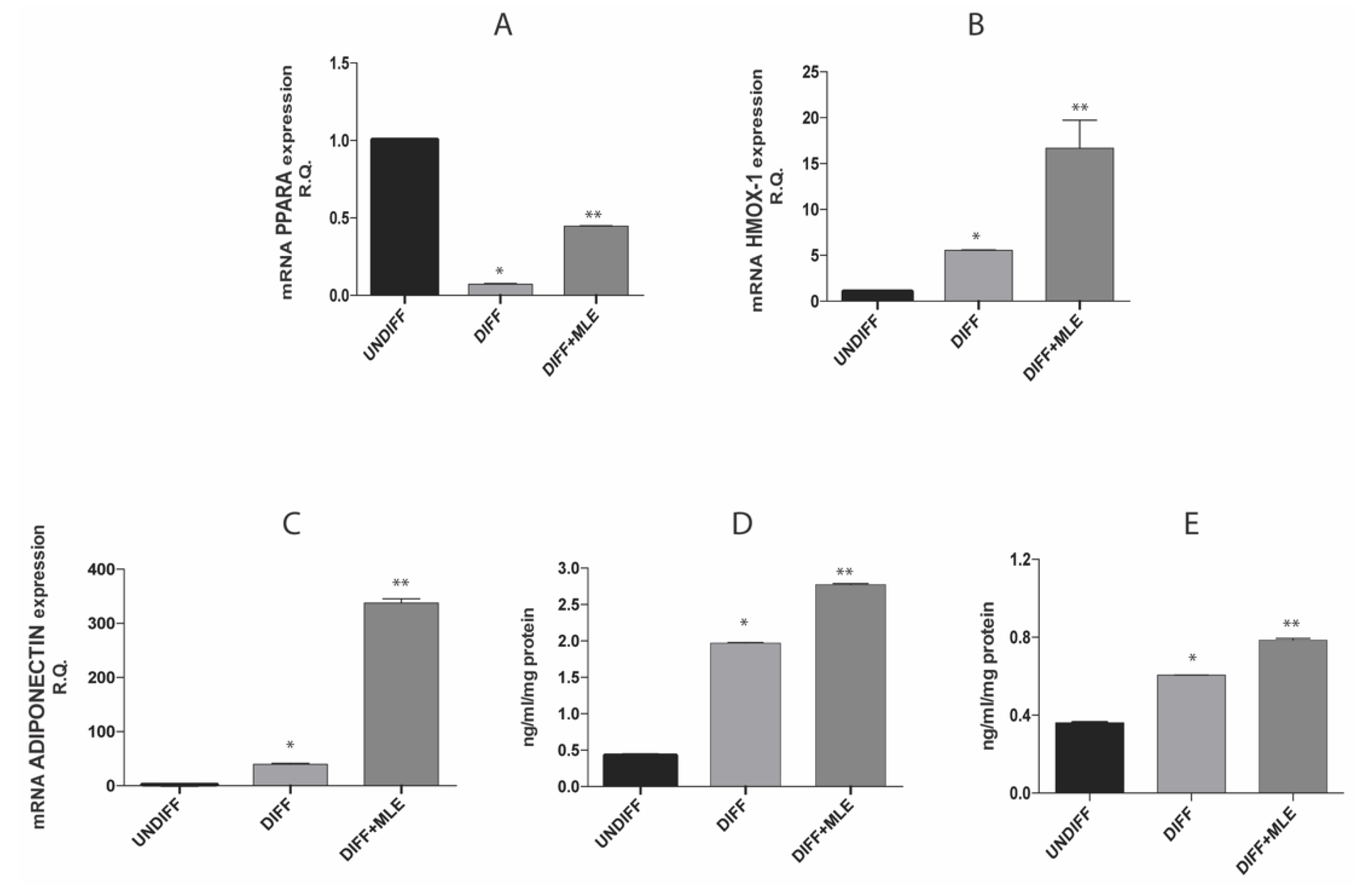
2.9. MLE Effect on PPAR-α, Heme oxygenase 1 (HO-1) and Adiponectin Expression
3. Discussion
4. Materials and Methods
4.1. Leaves Extract Preparation
4.2. Total Polyphenols Content
4.3. UPLC-Ms/Ms Analisys
4.4. Antioxidant Capacity
4.5. Measurement of Cell Viability: Bromide 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT) Assay
4.6. Alpha-Glucosidase Assay
4.7. Differentiation of 3T3-L1 into Adipocytes
4.8. Oil Red Staining
4.9. RNA Extraction and qRT-PCR
4.10. GSH Determination
4.11. Determination of Adiponectin by Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | Adenosine monophosphate-activated protein kinase |
| CAT | Catalase |
| DGAT-1 | Diacylglycerol-O-acyltransferase-1 |
| DGAT-2 | Diacylglycerol-O-acyltransferase-2 |
| DMSO | Dimethyl sulfoxide |
| DPPH | 1,1-Diphenyl-2-picryl-hydrazyl |
| FAS | Fatty Acid Synthase |
| FFA | Free fatty acids |
| GAPDH | Glyceraldeyde-3-phosphate dehydrogenase |
| GLUT4 | Glucose transporter 4 |
| GSH | Glutathione |
| HO-1 | Heme oxygenase-1 |
| IL-10 | Interleukin-10 |
| IL-6 | Interleukin 6 |
| LOD | Limit of detection |
| LOQ | Limit of quantification |
| MLE | Mangifera indica L. leaf extract |
| MnSOD | Manganese-dependent superoxide dismutase |
| OD | Optical density |
| ORZ | γ-oryzanol |
| PPAR-α | Peroxisome proliferator activated receptor-alpha |
| PPAR-γ | Peroxisome proliferator activated receptor-gamma |
| SCD-1 | Stearoyl-coenzyme A desaturase 1 |
| TG | Triglycerides |
References
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef] [PubMed]
- Beckman, C.H. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant. Pathol. 2000, 57, 101–110. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Mangos and their bioactive components: Adding variety to the fruit plate for health. Food Funct. 2017, 8, 3010–3032. [Google Scholar] [CrossRef] [PubMed]
- Pokorski, M.; Pozdzik, M.; Mazzatenta, A. Antioxidant treatment for impaired hypoxic ventilatory responses in experimental diabetes in the rat. Respir. Physiol. Neurobiol. 2018, 255, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Illesca, P.; Valenzuela, R.; Espinosa, A.; Echeverria, F.; Soto-Alarcon, S.; Ortiz, M.; Videla, L.A. Hydroxytyrosol supplementation ameliorates the metabolic disturbances in white adipose tissue from mice fed a high-fat diet through recovery of transcription factors Nrf2, SREBP-1c, PPAR-gamma and NF-kappaB. Biomed. Pharm. 2019, 109, 2472–2481. [Google Scholar] [CrossRef] [PubMed]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R. A Review on Ethnopharmacological Applications, Pharmacological Activities, and Bioactive Compounds of Mangifera indica (Mango). Evid.-Based Complement. Altern. Med. 2017, 2017, 6949835. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Ferrante, P.; Vitale, A.; Polizzi, G.; Scortichini, M.; Cirvilleri, G. Characterization of Pseudomonas syringae pv. syringae isolated from mango in Sicily and occurrence of copper-resistant strains. J. Plant. Pathol. 2015, 97, 273–282. [Google Scholar]
- Schieber, A.; Ullrich, W.; Carle, R. Characterization of polyphenols in mango puree concentrate by HPLC with diode array and mass spectrometric detection. Innov. Food Sci. Emerg. Technol. 2000, 1, 161–166. [Google Scholar] [CrossRef]
- Meneses, M.A.; Caputo, G.; Scognamiglio, M.; Reverchon, E.; Adami, R. Antioxidant phenolic compounds recovery from Mangifera indica L. by-products by supercritical antisolvent extraction. J. Food Eng. 2015, 163, 45–53. [Google Scholar] [CrossRef]
- Ling, L.T.; Yap, S.-A.; Radhakrishnan, A.K.; Subramaniam, T.; Cheng, H.M.; Palanisamy, U.D. Standardised Mangifera indica extract is an ideal antioxidant. Food Chem. 2009, 113, 1154–1159. [Google Scholar] [CrossRef]
- Abdullah, A.S.; Mohammed, A.S.; Rasedee, A.; Mirghani, M.E.; Al-Qubaisi, M.S. Induction of apoptosis and oxidative stress in estrogen receptor-negative breast cancer, MDA-MB231 cells, by ethanolic mango seed extract. BMC Complement. Altern. Med. 2015, 15, 45. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, N.M.; Toledo, R.C.L.; Moreira, M.E.C.; Martino, H.S.D.; Benjamin, L.D.A.; de Queiroz, J.H.; Ribeiro, A.Q.; Ribeiro, S.M.R. Anti-obesity effects of tea from Mangifera indica L. leaves of the Uba variety in high-fat diet-induced obese rats. Biomed. Pharmacother. 2017, 91, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Samaras, K.; Botelho, N.K.; Chisholm, D.J.; Lord, R.V. Subcutaneous and visceral adipose tissue gene expression of serum adipokines that predict type 2 diabetes. Obesity 2010, 18, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Fried, S.K.; Bunkin, D.A.; Greenberg, A.S. Omental and subcutaneous adipose tissues of obese subjects release interleukin-6: Depot difference and regulation by glucocorticoid. J. Clin. Endocrinol. Metab. 1998, 83, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Walsh, K. Obesity, adiponectin and vascular inflammatory disease. Curr. Opin. Lipidol. 2003, 14, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun. 1999, 257, 79–83. [Google Scholar] [CrossRef]
- Li, S.; Shin, H.J.; Ding, E.L.; van Dam, R.M. Adiponectin levels and risk of type 2 diabetes: A systematic review and meta-analysis. JAMA 2009, 302, 179–188. [Google Scholar] [CrossRef]
- Ronsisvalle, S.; Lissandrello, E.; Fuochi, V.; Petronio Petronio, G.; Straquadanio, C.; Crasci, L.; Panico, A.; Milito, M.; Cova, A.M.; Tempera, G.; et al. Antioxidant and antimicrobial properties of Casteanea sativa Miller chestnut honey produced on Mount Etna (Sicily). Nat. Prod. Res. 2017, 33, 843–850. [Google Scholar] [CrossRef]
- Saleh Gazwi, H.S.; Mahmoud, M.E. Restorative activity of aqueous extract Mangifera indica leaves against CCl4 induced hepatic damage in rats. J. Pharm. Biomed. Anal. 2019, 164, 112–118. [Google Scholar] [CrossRef]
- Rosalie, R.; Lechaudel, M.; Dhuique-Mayer, C.; Dufosse, L.; Joas, J. Antioxidant and enzymatic responses to oxidative stress induced by cold temperature storage and ripening in mango (Mangifera indica L. cv. ‘Cogshall’) in relation to carotenoid content. J. Plant. Physiol. 2018, 224–225, 75–85. [Google Scholar] [CrossRef]
- Selles, A.J.; Villa, D.G.; Rastrelli, L. Mango polyphenols and its protective effects on diseases associated to oxidative stress. Curr. Pharm. Biotechnol. 2015, 16, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Li, S.; Na, L.; Feng, R.; Liu, L.; Li, Y.; Sun, C. Mangiferin decreases plasma free fatty acids through promoting its catabolism in liver by activation of AMPK. PLoS ONE 2012, 7, e30782. [Google Scholar] [CrossRef] [PubMed]
- Na, L.; Zhang, Q.; Jiang, S.; Du, S.; Zhang, W.; Li, Y.; Sun, C.; Niu, Y. Mangiferin supplementation improves serum lipid profiles in overweight patients with hyperlipidemia: A double-blind randomized controlled trial. Sci. Rep. 2015, 5, 10344. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Zi, T.; Liu, L.; Feng, R.; Sun, C. A 1H-NMR based metabolomics study of the intervention effect of mangiferin on hyperlipidemia hamsters induced by a high-fat diet. Food Funct. 2017, 8, 2455–2464. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Zheng, D.; Zhong, G.; Hu, Y. Mangiferin mitigates diabetic cardiomyopathy in streptozotocin-diabetic rats. Can. J. Physiol Pharm. 2013, 91, 759–763. [Google Scholar] [CrossRef]
- Unfer, V.; Carlomagno, G.; Dante, G.; Facchinetti, F. Effects of myo-inositol in women with PCOS: A systematic review of randomized controlled trials. Gynecol. Endocrinol. 2012, 28, 509–515. [Google Scholar] [CrossRef]
- Heimark, D.; McAllister, J.; Larner, J. Decreased myo-inositol to chiro-inositol (M/C) ratios and increased M/C epimerase activity in PCOS theca cells demonstrate increased insulin sensitivity compared to controls. Endocr. J. 2014, 61, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Shokrpour, M.; Foroozanfard, F.; Afshar Ebrahimi, F.; Vahedpoor, Z.; Aghadavod, E.; Ghaderi, A.; Asemi, Z. Comparison of myo-inositol and metformin on glycemic control, lipid profiles, and gene expression related to insulin and lipid metabolism in women with polycystic ovary syndrome: A randomized controlled clinical trial. Gynecol. Endocrinol. 2019, 35, 406–411. [Google Scholar] [CrossRef]
- Kozuka, C.; Yabiku, K.; Takayama, C.; Matsushita, M.; Shimabukuro, M. Natural food science based novel approach toward prevention and treatment of obesity and type 2 diabetes: Recent studies on brown rice and gamma-oryzanol. Obes. Res. Clin. Pract. 2013, 7, e165–e172. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhang, G.; Liao, Y.; Gong, D. Inhibitory kinetics and mechanism of kaempferol on alpha-glucosidase. Food Chem. 2016, 190, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Rudich, A.; Tirosh, A.; Potashnik, R.; Hemi, R.; Kanety, H.; Bashan, N. Prolonged oxidative stress impairs insulin-induced GLUT4 translocation in 3T3-L1 adipocytes. Diabetes 1998, 47, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Toita, R.; Kawano, T.; Murata, M.; Kang, J.H. Anti-obesity and anti-inflammatory effects of macrophage-targeted interleukin-10-conjugated liposomes in obese mice. Biomaterials 2016, 110, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, A.; Yamauchi, T.; Takekawa, S.; Hada, Y.; Ito, Y.; Maki, T.; Kadowaki, T. Peroxisome proliferator-activated receptor (PPAR)alpha activation increases adiponectin receptors and reduces obesity-related inflammation in adipose tissue: Comparison of activation of PPARα, PPARγ, and their combination. Diabetes 2005, 54, 3358–3370. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, N.; Jialal, I. Biochemistry, Adiponectin; StatPearls Publishing LLC: St. Petersburg, FL, USA, 2019. [Google Scholar]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed]
- Turer, A.T.; Scherer, P.E. Adiponectin: Mechanistic insights and clinical implications. Diabetologia 2012, 55, 2319–2326. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; Arita, Y. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nat. Med. 2002, 8, 731. [Google Scholar] [CrossRef]
- Nawrocki, A.R.; Rajala, M.W.; Tomas, E.; Pajvani, U.B.; Saha, A.K.; Trumbauer, M.E.; Pang, Z.; Chen, A.S.; Ruderman, N.B.; Chen, H. Mice lacking adiponectin show decreased hepatic insulin sensitivity and reduced responsiveness to peroxisome proliferator-activated receptor γ agonists. J. Biol. Chem. 2006, 281, 2654–2660. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Van De Wall, E.; Laplante, M.; Azzara, A.; Trujillo, M.E.; Hofmann, S.M.; Schraw, T.; Durand, J.L.; Li, H.; Li, G. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J. Clin. Investig. 2007, 117, 2621–2637. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Kim, D.H.; Tsenovoy, P.L.; Peterson, S.J.; Rezzani, R.; Rodella, L.F.; Aronow, W.S.; Ikehara, S.; Abraham, N.G. Treatment of obese diabetic mice with a heme oxygenase inducer reduces visceral and subcutaneous adiposity, increases adiponectin levels, and improves insulin sensitivity and glucose tolerance. Diabetes 2008, 57, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zang, P.; Han, F.; Hou, N.; Sun, X. Renal protective effects of induction of haem oxygenase-1 combined with increased adiponectin on the glomerular vascular endothelial growth factor-nitric oxide axis in obese rats. Exp. Physiol. 2015, 100, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, I.; Nicolosi, A.; Calabrese, G.; David, S.; Cimino, S.; Madonia, M.; Cappello, F. The role of the heme oxygenase system in the metabolic syndrome. Curr. Pharm. Des. 2014, 20, 4970–4974. [Google Scholar] [CrossRef] [PubMed]
- Hou, N.; Liu, Y.; Han, F.; Wang, D.; Hou, X.; Hou, S.; Sun, X. Irisin improves perivascular adipose tissue dysfunction via regulation of the heme oxygenase-1/adiponectin axis in diet-induced obese mice. J. Mol. Cell Cardiol. 2016, 99, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Peterson, S.J.; Sodhi, K.; Vanella, L.; Barbagallo, I.; Rodella, L.F.; Schwartzman, M.L.; Abraham, N.G.; Kappas, A. Heme oxygenase gene targeting to adipocytes attenuates adiposity and vascular dysfunction in mice fed a high-fat diet. Hypertension 2012, 60, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Singh, S.P.; McClung, J.A.; Joseph, G.; Vanella, L.; Barbagallo, I.; Jiang, H.; Falck, J.R.; Arad, M.; Shapiro, J.I.; et al. EET intervention on Wnt1, NOV, and HO-1 signaling prevents obesity-induced cardiomyopathy in obese mice. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H368–H380. [Google Scholar] [CrossRef] [Green Version]
- Vanella, L.; Russo, G.I.; Cimino, S.; Fragala, E.; Favilla, V.; Li Volti, G.; Barbagallo, I.; Sorrenti, V.; Morgia, G. Correlation between lipid profile and heme oxygenase system in patients with benign prostatic hyperplasia. Urology 2014, 83, 1444.e7–1444.e13. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Han, L.; Gao, X.; Liu, E.; Wang, T. Regulation of lipid and glucose homeostasis by mango tree leaf extract is mediated by AMPK and PI3K/AKT signaling pathways. Food Chem. 2013, 141, 2896–2905. [Google Scholar] [CrossRef]
- Vázquez, A.J.C.; Janer, M.L. Determinación de los polifenoles totales del aceite de oliva. Grasas Aceites 1973, 24, 350–355. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Concentration (ng/mL) | LOD | LOQ | r2 |
|---|---|---|---|---|
| Mangiferin | 70,200 ±150 | 1875 | 7320 | 0.9985 |
| Myo-inositol | 21,600 ± 230 | 832 | 4250 | 0.9973 |
| γ-Orizanolo | 47,700 ± 190 | 1432 | 5265 | 0.9987 |
| Gene GenBank Code | Forward and Reverse Primers Sequence (5′→3′) | Tm °C | Amplicon Size |
|---|---|---|---|
| Adiponectin NM_009605 | GAAGCCGCTTATGTGTATCGC | 61.5 | 76 |
| GAATGGGTACATTGGGAACAGT | 60.0 | ||
| CAT NM_009804.2 | AAGATTGCCTTCTCCGGGTG | 60.04 | 430 |
| TGTGGAGAATCGAACGGCAA | 59.97 | ||
| DGAT1 NM_010046.3 | GTTTCCGTCCAGGGTGGTAG | 287 | 871 |
| GTTGGATCAGCCCCACTTGA | 1119 | ||
| DGAT2 NM_026384.3 | CAGGTGCCGTCTTGGGTTAT | 1932 | 100 |
| CAGGAGGATATGCGCCAGAG | 1993 | ||
| FAS NM_007988 | GGAGGTGGTGATAGCCGGTAT | 62.9 | 140 |
| TGGGTAATCCATAGAGCCCAG | 60.4 | ||
| GAPDH NM_008085 | AGCTTCGGCACATATTTCATCTG | 61.0 | 89 |
| CGTTCACTCCCATGACAAACA | 60.5 | ||
| HMOX1 NM_010442.2 | CCTCACAGATGGCGTCACTT | 840 | 200 |
| TGGGGGCCAGTATTGCATTT | 1001 | ||
| Il-10 NM_010548 | GCTGGACAACATACTGCTAACC | 60.9 | 78 |
| ATTTCCGATAAGGCTTGGCAA | 60.0 | ||
| Il-6 NM_031168.2 | CCCCAATTTCCAATGCTCTCC | 598 | 141 |
| CGCACTAGGTTTGCCGAGTA | 699 | ||
| MnSOD NM_009127.4 | GCCCAAACCTATCGTGTCCA | 3102 | 70 |
| AGGGAACCCTAAATGCTGCC | 3133 | ||
| PPARα NM_011144.6 | CCGAACATTGGTGTTCGCAG | 135 | 161 |
| AGATACGCCCAAATGCACCA | 257 | ||
| PPARγ 19016 | TCGCTGATGCACTGCCTATG | 62.4 | 103 |
| GAGAGGTCCACAGAGCTGATT | 60.9 | ||
| SCD1 NM_009127.4 | GAGTAGCTGAGCTTTGGGCT | 1378 | 591 |
| ACTTCATCAGCGGGGACTTG | 1930 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sferrazzo, G.; Palmeri, R.; Vanella, L.; Parafati, L.; Ronsisvalle, S.; Biondi, A.; Basile, F.; Li Volti, G.; Barbagallo, I. Mangifera indica L. Leaf Extract Induces Adiponectin and Regulates Adipogenesis. Int. J. Mol. Sci. 2019, 20, 3211. https://doi.org/10.3390/ijms20133211
Sferrazzo G, Palmeri R, Vanella L, Parafati L, Ronsisvalle S, Biondi A, Basile F, Li Volti G, Barbagallo I. Mangifera indica L. Leaf Extract Induces Adiponectin and Regulates Adipogenesis. International Journal of Molecular Sciences. 2019; 20(13):3211. https://doi.org/10.3390/ijms20133211
Chicago/Turabian StyleSferrazzo, Giuseppe, Rosa Palmeri, Luca Vanella, Lucia Parafati, Simone Ronsisvalle, Antonio Biondi, Francesco Basile, Giovanni Li Volti, and Ignazio Barbagallo. 2019. "Mangifera indica L. Leaf Extract Induces Adiponectin and Regulates Adipogenesis" International Journal of Molecular Sciences 20, no. 13: 3211. https://doi.org/10.3390/ijms20133211
APA StyleSferrazzo, G., Palmeri, R., Vanella, L., Parafati, L., Ronsisvalle, S., Biondi, A., Basile, F., Li Volti, G., & Barbagallo, I. (2019). Mangifera indica L. Leaf Extract Induces Adiponectin and Regulates Adipogenesis. International Journal of Molecular Sciences, 20(13), 3211. https://doi.org/10.3390/ijms20133211