Single Mild Traumatic Brain Injury Induces Persistent Disruption of the Blood-Brain Barrier, Neuroinflammation and Cognitive Decline in Hypertensive Rats

 , , ,
, , , 
 , , ,
, , , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
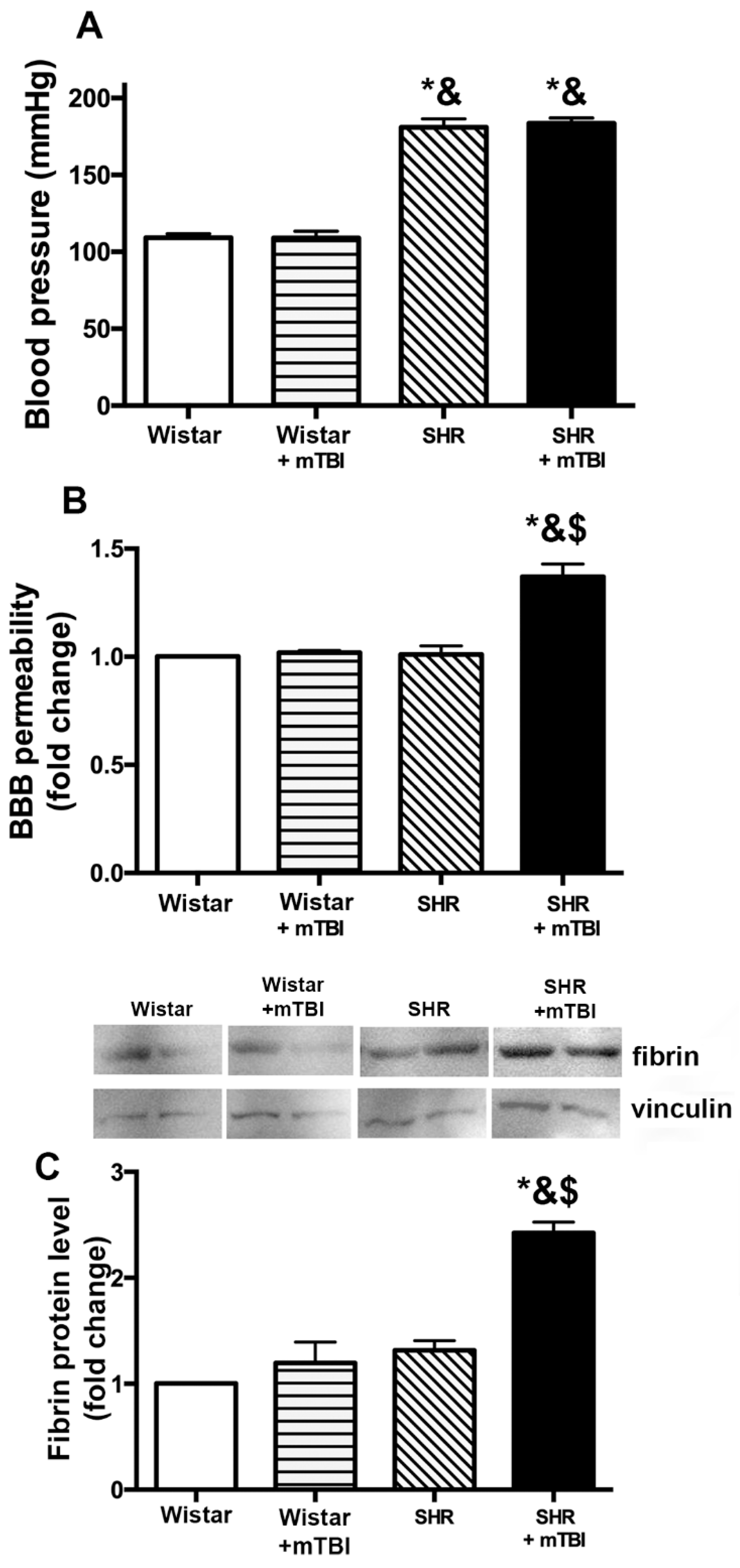
2.1. Mild TBI Induces Persistent Disruption of the Blood-Brain Barrier Which is Associated with Extravasation and Accumulation of Blood Borne Fibrin in Cerebral Tissue of Spontaneously Hypertensive Rats
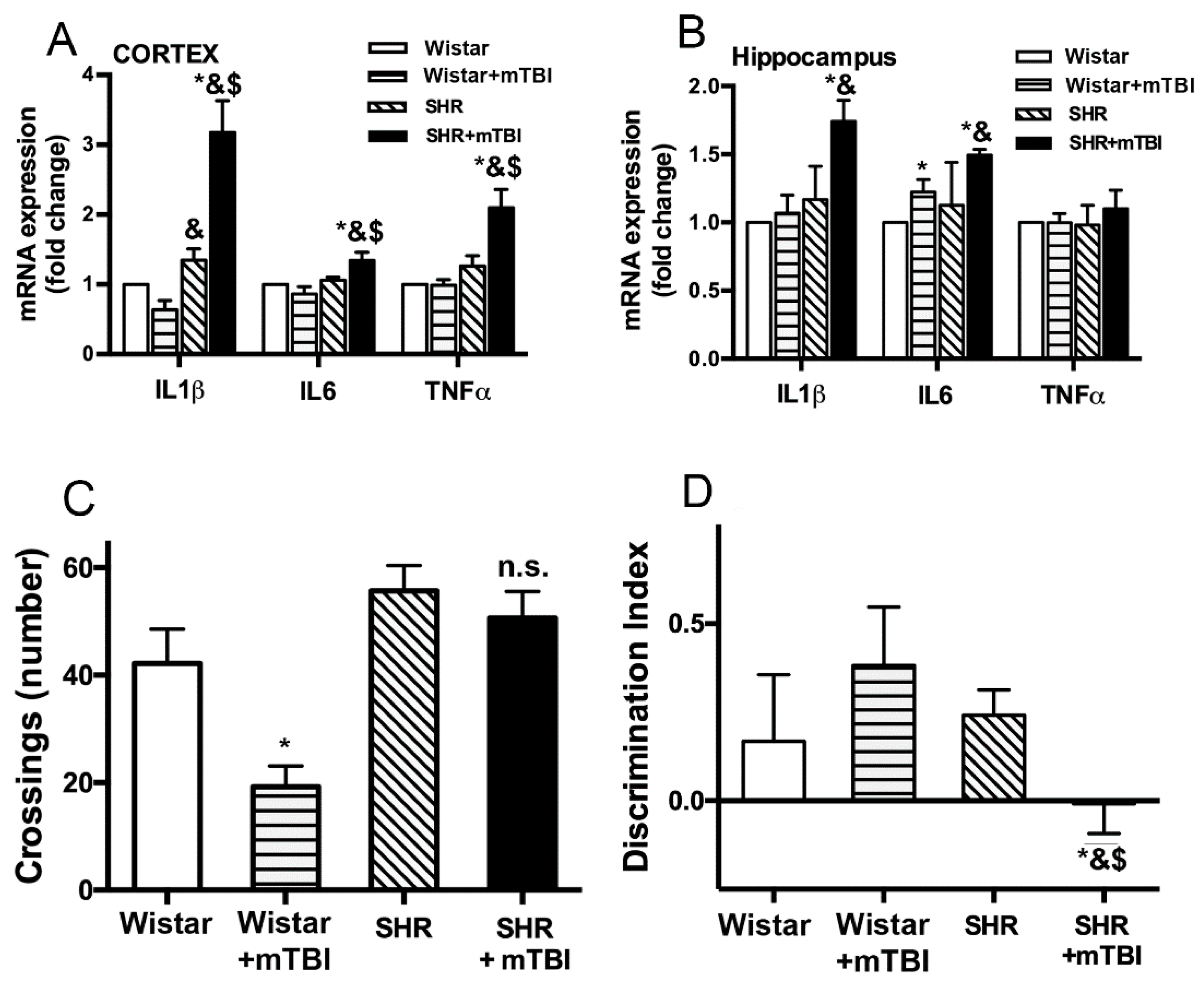
2.2. Mild TBI Induces Persistent Neuroinflammation and Cognitive Decline in Spontaneously Hypertensive Rats
3. Discussion
Limitations of the Study
4. Materials and Methods
4.1. Mild Traumatic Brain Injury (mTBI) in Normotensive and Hypertensive Rats
4.2. Behavioral Studies
4.2.1. Open Field Test (OFT)
4.2.2. Novel Object Recognition Test (NOR)
4.3. Blood-Brain Barrier Disruption
4.4. Western Blotting
4.5. Quantitative Real-Time RT-PCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TBI | traumatic brain injury |
| mTBI | mild traumatic brain injury |
| BBB | blood-brain barrier |
| IL | interleukin |
| TNF | tumor necrosis factor |
| SHR | spontaneously hypertensive rat |
| ICU | intensive care unit |
| DI | discrimination index |
| ROS | reactive oxygen species |
| OFT | open field test |
| NOR | novel object recognition test |
| TCA | trichloroacetic acid |
References
- Schwarzmaier, S.M.; Terpolilli, N.A.; Dienel, A.; Gallozzi, M.; Schinzel, R.; Tegtmeier, F.; Plesnila, N. Endothelial nitric oxide synthase mediates arteriolar vasodilatation after traumatic brain injury in mice. J. Neurotrauma 2014, 32, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Sahuquillo, J.; Poca, M.A.; Amoros, S. Current aspects of pathophysiology and cell dysfunction after severe head injury. Curr. Pharm. Des. 2001, 7, 1475–1503. [Google Scholar] [CrossRef] [PubMed]
- Greve, M.W.; Zink, B.J. Pathophysiology of traumatic brain injury. Mt. Sinai J. Med. New York 2009, 76, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Alluri, H.; Wiggins-Dohlvik, K.; Davis, M.L.; Huang, J.H.; Tharakan, B. Blood-brain barrier dysfunction following traumatic brain injury. Metab. Brain Dis. 2015, 30, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Gardner, R.C.; Yaffe, K. Epidemiology of mild traumatic brain injury and neurodegenerative disease. Mol. Cell Neurosci. 2015, 66, 75–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Grande, B.; Ichkova, A.; Lemarchant, S.; Badaut, J. Early to long-term alterations of cns barriers after traumatic brain injury: Considerations for drug development. Aaps. J. 2017, 19, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Watts, L.; Long, J.; Zhou, W.; Shen, Q.; Jiang, Z.; Li, Y.; Duong, T.Q. Spatiotemporal changes in blood-brain barrier permeability, cerebral blood flow, t2 and diffusion following mild traumatic brain injury. Brain Res. 2016, 1646, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Johnson, V.E.; Weber, M.T.; Xiao, R.; Cullen, D.K.; Meaney, D.F.; Stewart, W.; Smith, D.H. Mechanical disruption of the blood-brain barrier following experimental concussion. Acta Neuropathol 2018, 135, 711–726. [Google Scholar] [CrossRef]
- Kumar, R.G.; Juengst, S.B.; Wang, Z.; Dams-O’Connor, K.; Dikmen, S.S.; O’Neil-Pirozzi, T.M.; Dahdah, M.N.; Hammond, F.M.; Felix, E.R.; Arenth, P.M.; et al. Epidemiology of comorbid conditions among adults 50 years and older with traumatic brain injury. J. Head Trauma Rehabil. 2018, 33, 15–24. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Report to Congress on Traumatic Brain Injury in the United States: Epidemiology and Rehabilitation; Division of Unintentional Injury Prevention; National Center for Injury Prevention and Control: Atlanta, GA, USA, 2015.
- Thompson, H.J.; Dikmen, S.; Temkin, N. Prevalence of comorbidity and its association with traumatic brain injury and outcomes in older adults. Res. Gerontol Nurs. 2012, 5, 17–24. [Google Scholar] [CrossRef]
- Toth, P.; Tucsek, Z.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Tarantini, S.; Deak, F.; Koller, A.; Sonntag, W.E.; Csiszar, A.; et al. Age-related autoregulatory dysfunction and cerebromicrovascular injury in mice with angiotensin ii-induced hypertension. J. Cereb Blood Flow Metab. 2013, 33, 1732–1742. [Google Scholar] [CrossRef] [PubMed]
- Tucsek, Z.; Toth, P.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Koller, A.; Szalai, G.; Sonntag, W.E.; Ungvari, Z.; Csiszar, A. Obesity in aging exacerbates blood-brain barrier disruption, neuroinflammation, and oxidative stress in the mouse hippocampus: Effects on expression of genes involved in beta-amyloid generation and alzheimer’s disease. J. Gerontol. A. Biol. Sci. Med. Sci. 2013, 69, 1212–1226. [Google Scholar] [CrossRef] [PubMed]
- Marcano, D.C.; Bitner, B.R.; Berlin, J.M.; Jarjour, J.; Lee, J.M.; Jacob, A.; Fabian, R.H.; Kent, T.A.; Tour, J.M. Design of poly(ethylene glycol)-functionalized hydrophilic carbon clusters for targeted therapy of cerebrovascular dysfunction in mild traumatic brain injury. J. Neurotrauma 2013, 30, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Szarka, N.; Pabbidi, M.R.; Amrein, K.; Czeiter, E.; Berta, G.; Pohoczky, K.; Helyes, Z.; Ungvari, Z.; Koller, A.; Buki, A.; et al. Traumatic brain injury impairs myogenic constriction of cerebral arteries: Role of mitochondria-derived h2o2 and trpv4-dependent activation of bkca channels. J. Neurotrauma 2018, 35, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.C.; Silva, K.C.; Biswas, S.K.; Martins, N.; De Faria, J.B.; De Faria, J.M. Arterial hypertension exacerbates oxidative stress in early diabetic retinopathy. Free Radic Res. 2007, 41, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Marklund, N.; Clausen, F.; Lewander, T.; Hillered, L. Monitoring of reactive oxygen species production after traumatic brain injury in rats with microdialysis and the 4-hydroxybenzoic acid trapping method. J. Neurotrauma 2001, 18, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Schreibelt, G.; Kooij, G.; Reijerkerk, A.; van Doorn, R.; Gringhuis, S.I.; van der Pol, S.; Weksler, B.B.; Romero, I.A.; Couraud, P.O.; Piontek, J.; et al. Reactive oxygen species alter brain endothelial tight junction dynamics via rhoa, pi3 kinase, and pkb signaling. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2007, 21, 3666–3676. [Google Scholar] [CrossRef]
- Haorah, J.; Ramirez, S.H.; Schall, K.; Smith, D.; Pandya, R.; Persidsky, Y. Oxidative stress activates protein tyrosine kinase and matrix metalloproteinases leading to blood-brain barrier dysfunction. J. Neurochem. 2007, 101, 566–576. [Google Scholar] [CrossRef]
- Sun, H.J.; Ren, X.S.; Xiong, X.Q.; Chen, Y.Z.; Zhao, M.X.; Wang, J.J.; Zhou, Y.B.; Han, Y.; Chen, Q.; Li, Y.H.; et al. Nlrp3 inflammasome activation contributes to vsmc phenotypic transformation and proliferation in hypertension. Cell Death Dis. 2017, 8, e3074. [Google Scholar] [CrossRef]
- Wang, Q.; Cui, Y.; Lin, N.; Pang, S. Correlation of cardiomyocyte apoptosis with duration of hypertension, severity of hypertension and caspase-3 expression in hypertensive rats. Exp. Ther. Med. 2019, 17, 2741–2745. [Google Scholar] [CrossRef]
- Montagne, A.; Nikolakopoulou, A.M.; Zhao, Z.; Sagare, A.P.; Si, G.; Lazic, D.; Barnes, S.R.; Daianu, M.; Ramanathan, A.; Go, A.; et al. Pericyte degeneration causes white matter dysfunction in the mouse central nervous system. Nat. Med. 2018, 24, 326. [Google Scholar] [CrossRef] [PubMed]
- Fiala, M.; Liu, Q.N.; Sayre, J.; Pop, V.; Brahmandam, V.; Graves, M.C.; Vinters, H.V. Cyclooxygenase-2-positive macrophages infiltrate the alzheimer’s disease brain and damage the blood-brain barrier. Eur. J. Clin. Investig. 2002, 32, 360–371. [Google Scholar] [CrossRef]
- Carrano, A.; Hoozemans, J.J.; van der Vies, S.M.; van Horssen, J.; de Vries, H.E.; Rozemuller, A.J. Neuroinflammation and blood-brain barrier changes in capillary amyloid angiopathy. Neuro-Degener. Dis. 2012, 10, 329–331. [Google Scholar] [CrossRef]
- Rochfort, K.D.; Cummins, P.M. The blood-brain barrier endothelium: A target for pro-inflammatory cytokines. Biochem. Soc. Trans. 2015, 43, 702–706. [Google Scholar] [CrossRef] [PubMed]
- Cortese, G.P.; Burger, C. Neuroinflammatory challenges compromise neuronal function in the aging brain: Postoperative cognitive delirium and alzheimer’s disease. Behav. Brain Res. 2017, 322, 269–279. [Google Scholar] [CrossRef]
- Jaworski, T.; Lechat, B.; Demedts, D.; Gielis, L.; Devijver, H.; Borghgraef, P.; Duimel, H.; Verheyen, F.; Kugler, S.; Van Leuven, F. Dendritic degeneration, neurovascular defects, and inflammation precede neuronal loss in a mouse model for tau-mediated neurodegeneration. Am. J. Pathol. 2011, 179, 2001–2015. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Trivedi, R.; Devi, M.M.; Tripathi, R.P.; Khushu, S. Longitudinal changes in the dti measures, anti-gfap expression and levels of serum inflammatory cytokines following mild traumatic brain injury. Exp. Neurol. 2016, 275 Pt 3, 427–435. [Google Scholar] [CrossRef]
- Singh, K.; Trivedi, R.; Haridas, S.; Manda, K.; Khushu, S. Study of neurometabolic and behavioral alterations in rodent model of mild traumatic brain injury: A pilot study. Nmr Biomed. 2016, 29, 1748–1758. [Google Scholar] [CrossRef]
- Szarka, N.; Amrein, K.; Horvath, P.; Ivic, I.; Czeiter, E.; Buki, A.; Koller, A.; Toth, P. Hypertension-induced enhanced myogenic constriction of cerebral arteries is preserved after traumatic brain injury. J. Neurotrauma 2017. [Google Scholar] [CrossRef]
- Muller, U.; Cristina, N.; Li, Z.W.; Wolfer, D.P.; Lipp, H.P.; Rulicke, T.; Brandner, S.; Aguzzi, A.; Weissmann, C. Behavioral and anatomical deficits in mice homozygous for a modified beta-amyloid precursor protein gene. Cell 1994, 79, 755–765. [Google Scholar] [CrossRef]
- Tarjanyi, O.; Berta, G.; Harci, A.; Bacsa, E.B.; Stark, B.; Pap, M.; Szeberenyi, J.; Setalo, G., Jr. The role of src protein in the process formation of pc12 cells induced by the proteasome inhibitor mg-132. Neurochem. Int. 2013, 63, 413–422. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szarka, N.; Toth, L.; Czigler, A.; Kellermayer, Z.; Ungvari, Z.; Amrein, K.; Czeiter, E.; Bali, Z.K.; Tadepalli, S.A.; Wahr, M.; et al. Single Mild Traumatic Brain Injury Induces Persistent Disruption of the Blood-Brain Barrier, Neuroinflammation and Cognitive Decline in Hypertensive Rats. Int. J. Mol. Sci. 2019, 20, 3223. https://doi.org/10.3390/ijms20133223
Szarka N, Toth L, Czigler A, Kellermayer Z, Ungvari Z, Amrein K, Czeiter E, Bali ZK, Tadepalli SA, Wahr M, et al. Single Mild Traumatic Brain Injury Induces Persistent Disruption of the Blood-Brain Barrier, Neuroinflammation and Cognitive Decline in Hypertensive Rats. International Journal of Molecular Sciences. 2019; 20(13):3223. https://doi.org/10.3390/ijms20133223
Chicago/Turabian StyleSzarka, Nikolett, Luca Toth, Andras Czigler, Zoltan Kellermayer, Zoltan Ungvari, Krisztina Amrein, Endre Czeiter, Zsolt Kristof Bali, Sai Ambika Tadepalli, Matyas Wahr, and et al. 2019. "Single Mild Traumatic Brain Injury Induces Persistent Disruption of the Blood-Brain Barrier, Neuroinflammation and Cognitive Decline in Hypertensive Rats" International Journal of Molecular Sciences 20, no. 13: 3223. https://doi.org/10.3390/ijms20133223
APA StyleSzarka, N., Toth, L., Czigler, A., Kellermayer, Z., Ungvari, Z., Amrein, K., Czeiter, E., Bali, Z. K., Tadepalli, S. A., Wahr, M., Hernadi, I., Koller, A., Buki, A., & Toth, P. (2019). Single Mild Traumatic Brain Injury Induces Persistent Disruption of the Blood-Brain Barrier, Neuroinflammation and Cognitive Decline in Hypertensive Rats. International Journal of Molecular Sciences, 20(13), 3223. https://doi.org/10.3390/ijms20133223