Polymorphism in the Promoter Region of NFE2L2 Gene Is a Genetic Marker of Susceptibility to Cirrhosis Associated with Alcohol Abuse

 , ,
, ,
Abstract
:1. Introduction
2. Results
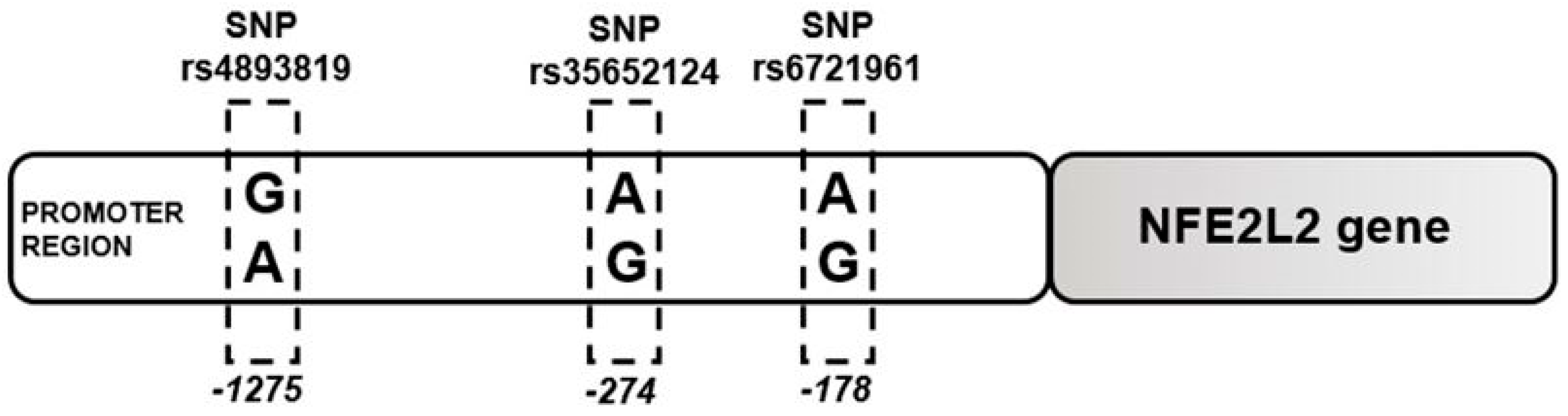
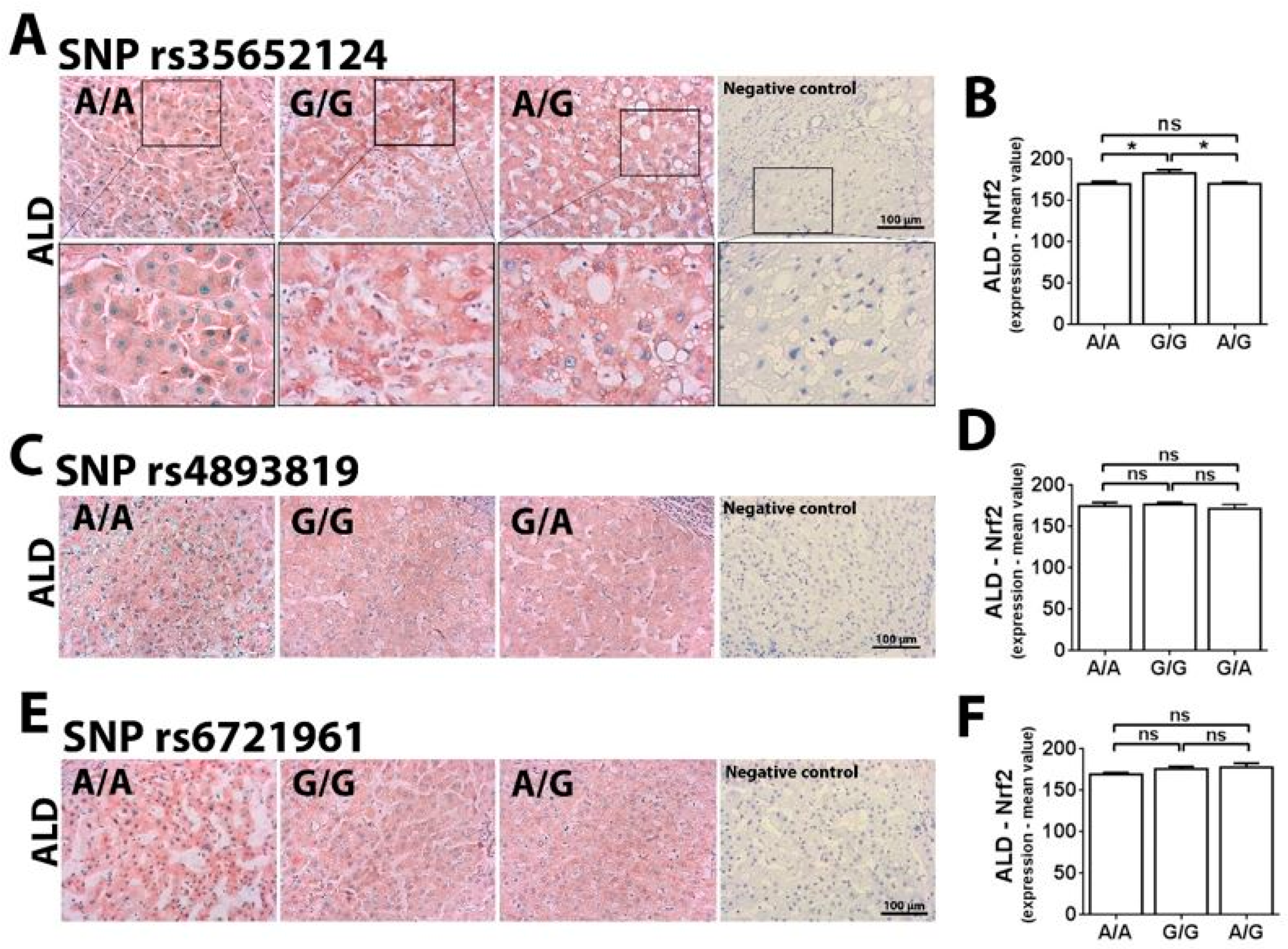
2.1. Single Nucleotide Polymorphism at -274 of the NFE2L2 Promoter Region Is a Genetic Marker for ALD Susceptibility
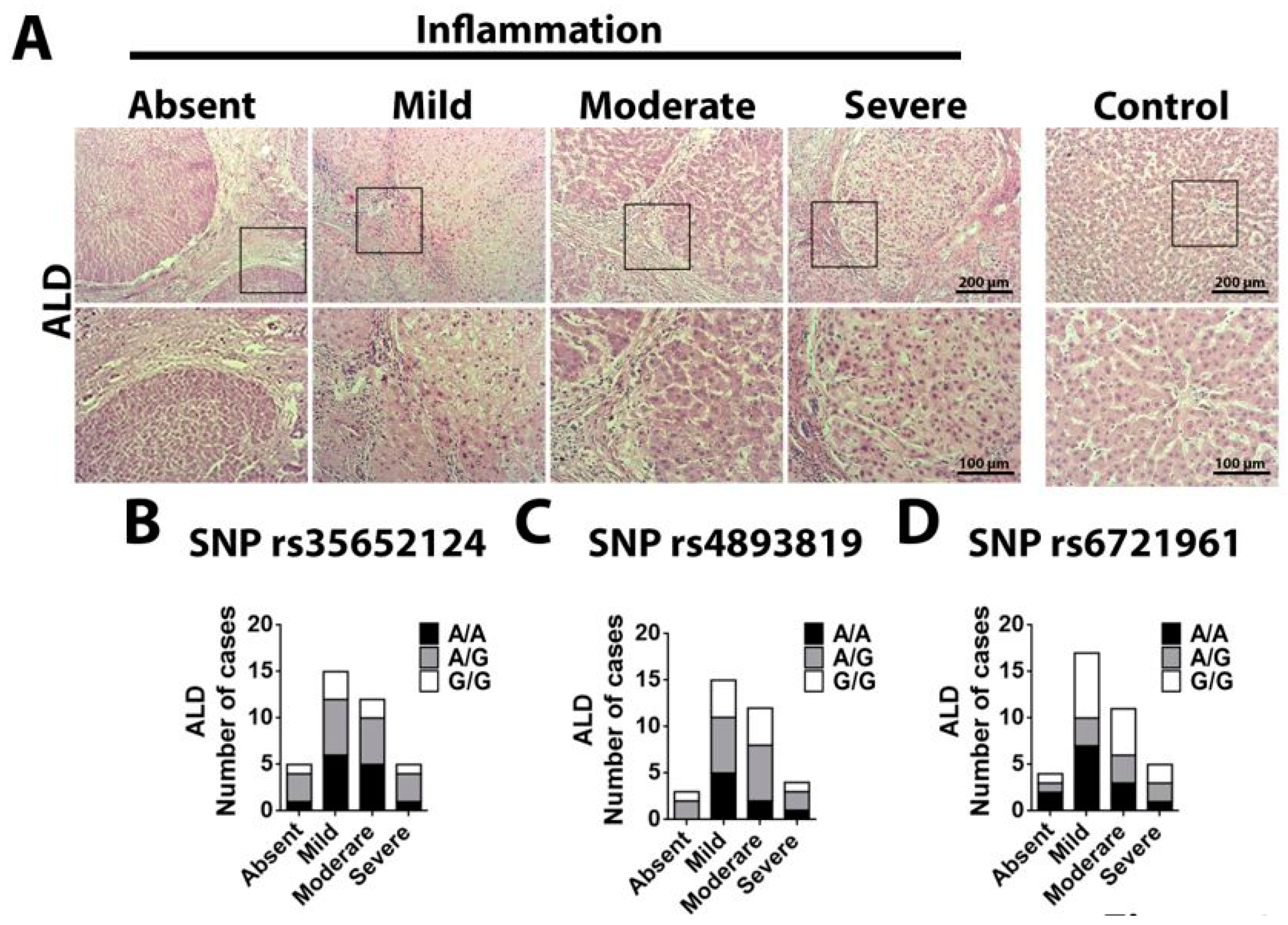
2.2. Single Nucleotide Polymorphism Rs35652124 in the NFE2L2 Gene Correlates With Inflammatory Score in ALD
3. Discussion
4. Materials and Methods
4.1. Human Specimens
4.2. Genotyping
4.3. Histopathological Analysis
4.4. Immunohistochemistry
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALD | Alcoholic Liver Disease |
| ROS | Reactive Oxygen Species |
| Nrf2 | Nuclear erythroid-related factor 2 |
| SNP | Single Nucleotide Polymorphism |
| NFE2L2 | Nuclear erythroid-related factor 2 gene |
| HCV | Hepatitis C virus |
| A | Adenine |
| G | Guanine |
| PNPLA3 | Patatin-like phospholipase domain-containing protein 3 |
| TM6SF2 | Transmembrane 6 superfamily member 2 |
| MBOAT7 | Membrane-bound O-acyltransferase domain-containing 7 |
| HCC | Hepatocellular carcinoma |
| NAFLD | Non-alcoholic fatty liver disease |
| VLDL | Very low-density lipoprotein |
| ARE | Antioxidant response element |
| GST | Glutathione-S-transferase |
| Q10 | Coenzyme Q10 |
| QR | NAD(P)Hiquinone oxidoreductase |
| SOD1 | Superoxide dismutase 1 |
| PD | Parkinson’s disease |
| M | Male |
| F | Female |
| MELD | Model for End-Stage Liver Disease |
| ALT | Alanine transaminase |
| AST | Aspartate transaminase |
| GGT | Gamma-glutamyltransferase |
| TB | Total bilirubin |
| DB | Direct bilirubin |
| IB | Indirect bilirubin |
| ALP | Alkaline phosphatase |
| LDH | Lactate dehydrogenase |
| ALB | Albumin |
| INR | International normalized ratio |
| UR | Urea |
| CR | Creatinine |
References
- Singal, A.K.; Bataller, R.; Ahn, J.; Kamath, P.S.; Shah, V.H. ACG clinical guideline: Alcoholic liver disease. Am. J. Gastroenterol. 2018, 113, 175–194. [Google Scholar] [CrossRef] [PubMed]
- Thursz, M.; Kamath, P.S.; Mathurin, P.; Szabo, G.; Shah, V.H.; Szabo, G.; Kamath, P.S.; Shah, V.H.; Thursz, M.; Mathurin, P.; et al. Alcohol-related liver disease: Areas of consensus, unmet needs and opportunities for further study. J. Hepatol. 2019, 70, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Tsukamoto, H. Inflammation in alcoholic and nonalcoholic fatty liver disease: Friend or foe? Gastroenterology 2016, 150, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Iranshahy, M.; Iranshahi, M.; Abtahi, S.R.; Karimi, G. The role of nuclear factor erythroid 2–related factor 2 in hepatoprotective activity of natural products: A review. Food Chem. Toxicol. 2018, 120, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Vomund, S.; Schafer, A.; Parnham, M.J.; Brune, B.; von Knethen, A. Nrf2, the master regulator of anti–oxidative responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I. Oxidative stress in age-related macular degeneration: Nrf2 as therapeutic target. Front. Pharmacol. 2018, 9, 1280. [Google Scholar] [CrossRef]
- Hennig, P.; Garstkiewicz, M.; Grossi, S.; Di Filippo, M.; French, L.E.; Beer, H.D. The crosstalk between Nrf2 and inflammasomes. Int. J. Mol. Sci. 2018, 19, 562. [Google Scholar] [CrossRef]
- Li, N.; Nel, A.E. Role of the Nrf2-mediated signaling pathway as a negative regulator of inflammation: Implications for the impact of particulate pollutants on asthma. Antioxid Redox Signal 2006, 8, 88–98. [Google Scholar] [CrossRef]
- Lu, C.; Xu, W.; Zhang, F.; Shao, J.; Zheng, S. Nrf2 knockdown disrupts the protective effect of curcumin on alcohol-induced hepatocyte necroptosis. Mol. Pharm. 2016, 13, 4043–4053. [Google Scholar] [CrossRef] [PubMed]
- Lamle, J.; Marhenke, S.; Borlak, J.; von Wasielewski, R.; Eriksson, C.J.; Geffers, R.; Manns, M.P.; Yamamoto, M.; Vogel, A. Nuclear factor-eythroid 2-related factor 2 prevents alcohol-induced fulminant liver injury. Gastroenterology 2008, 134, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Eun, J.W.; Cho, H.J.; Lee, H.Y.; Seo, C.W.; Noh, C.K.; Shin, S.J.; Lee, K.M.; Cho, S.W.; Cheong, J.Y. Effect of fibroblast growth factor-2 and its receptor gene polymorphisms on the survival of patients with hepatitis B virus-associated hepatocellular carcinoma. Anticancer Res. 2019, 39, 2217–2226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gong, X.; Yin, Z.; Cui, L.; Yang, J.; Wang, P.; Zhou, Y.; Jiang, X.; Wei, S.; Wang, F.; et al. Association between NRGN gene polymorphism and resting-state hippocampal functional connectivity in schizophrenia. BMC Psychiatry 2019, 19, 108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, H.; Liu, J.; Ouyang, Y.; Wang, D.; Bao, W.; Liu, L. Association between the NF-E2 related factor 2 gene polymorphism and oxidative stress, anti-oxidative status, and newly-diagnosed type 2 diabetes mellitus in a chinese population. Int. J. Mol. Sci. 2015, 16, 16483–16496. [Google Scholar] [CrossRef] [PubMed]
- Weerachayaphorn, J.; Amaya, M.J.; Spirli, C.; Chansela, P.; Mitchell-Richards, K.A.; Ananthanarayanan, M.; Nathanson, M.H. Nuclear factor, Erythroid 2-Like 2 regulates expression of type 3 Inositol 1,4,5-trisphosphate receptor and calcium signaling in cholangiocytes. Gastroenterology 2015, 149, 211–222.e10. [Google Scholar] [CrossRef] [PubMed]
- Purdom-Dickinson, S.E.; Sheveleva, E.V.; Sun, H.; Chen, Q.M. Translational control of nrf2 protein in activation of antioxidant response by oxidants. Mol. Pharmacol. 2007, 72, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- Covas, G.; Marinho, H.S.; Cyrne, L.; Antunes, F. Activation of Nrf2 by H2O2: De Novo Synthesis Versus Nuclear Translocation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2013; Volume 528, pp. 157–171. [Google Scholar]
- Milani, P.; Gagliardi, S.; Cova, E.; Cereda, C. SOD1 transcriptional and posttranscriptional regulation and its potential implications in ALS. Neurol. Res. Int. 2011, 2011, 458427. [Google Scholar] [CrossRef]
- Shasthry, S.M.; Rastogi, A.; Bihari, C.; Vijayaraghavan, R.; Arora, V.; Sharma, M.K.; Sarin, S.K. Histological activity score on baseline liver biopsy can predict non-response to steroids in patients with severe alcoholic hepatitis. Virchows Arch. 2018, 472, 667–675. [Google Scholar] [CrossRef]
- Brookes, A.J. The essence of SNPs. Gene 1999, 234, 177–186. [Google Scholar] [CrossRef]
- Stickel, F.; Datz, C.; Hampe, J.; Bataller, R. Pathophysiology and management of alcoholic liver disease: Update 2016. Gut Liver 2017, 11, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Magdaleno, F.; Blajszczak, C.C.; Nieto, N. Key events participating in the pathogenesis of alcoholic liver disease. Biomolecules 2017, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Reed, T.; Page, W.F.; Viken, R.J.; Christian, J.C. Genetic predisposition to organ-specific endpoints of alcoholism. Alcohol. Clin. Exp. Res. 1996, 20, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Toth, R.; Fiatal, S.; Petrovski, B.; McKee, M.; Adany, R. Combined effect of ADH1B RS1229984, RS2066702 and ADH1C RS1693482/ RS698 alleles on alcoholism and chronic liver diseases. Dis. Markers 2011, 31, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, T.; Yang, F.; Mao, Y.; Li, L.; Liu, C.; Sun, Q.; Li, Y.; Huang, J. Single-nucleotide rs738409 polymorphisms in the PNPLA3 gene are strongly associated with alcoholic liver disease in Han Chinese males. Hepatol. Int. 2018, 12, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Anstee, Q.M.; Daly, A.K.; Day, C.P. Genetics of alcoholic liver disease. Semin. Liver Dis. 2015, 35, 361–374. [Google Scholar] [PubMed]
- BasuRay, S.; Wang, Y.; Smagris, E.; Cohen, J.C.; Hobbs, H.H. Accumulation of PNPLA3 on lipid droplets is the basis of associated hepatic steatosis. Proc. Natl. Acad. Sci. USA 2019, 116, 9521–9526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liangpunsakul, S.; Puri, P.; Shah, V.H.; Kamath, P.; Sanyal, A.; Urban, T.; Ren, X.; Katz, B.; Radaeva, S.; Chalasani, N.; et al. Effects of age, sex, body weight, and quantity of alcohol consumption on occurrence and severity of alcoholic hepatitis. Clin. Gastroenterol. Hepatol. 2016, 14, 1831–1838.e3. [Google Scholar] [CrossRef]
- Atkinson, S.R.; Way, M.J.; McQuillin, A.; Morgan, M.Y.; Thursz, M.R. Homozygosity for rs738409:G in PNPLA3 is associated with increased mortality following an episode of severe alcoholic hepatitis. J. Hepatol. 2017, 67, 120–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buch, S.; Stickel, F.; Trépo, E.; Way, M.; Herrmann, A.; Nischalke, H.D.; Brosch, M.; Rosendahl, J.; Berg, T.; Ridinger, M.; et al. A genome-wide association study confirms PNPLA3 and identifies TM6SF2 and MBOAT7 as risk loci for alcohol-related cirrhosis. Nat. Genet. 2015, 47, 1443. [Google Scholar] [CrossRef] [PubMed]
- Salameh, H.; Raff, E.; Erwin, A.; Seth, D.; Nischalke, H.D.; Falleti, E.; Burza, M.A.; Leathert, J.; Romeo, S.; Molinaro, A.; et al. PNPLA3 gene polymorphism is associated with predisposition to and severity of alcoholic liver disease. Am. J. Gastroenterology 2015, 110, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, K.; Wannhoff, A.; Kattner, S.; Brune, M.; Hov, J.R.; Weiss, K.H.; Antoni, C.; Dollinger, M.; Neumann-Haefelin, C.; Seufferlein, T.; et al. PNPLA3 in end-stage liver disease: Alcohol consumption, hepatocellular carcinoma development, and transplantation-free survival. J. Gastroenterol. Hepatol. 2014, 29, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Falleti, E.; Cussigh, A.; Cmet, S.; Fabris, C.; Toniutto, P. PNPLA3 rs738409 and TM6SF2 rs58542926 variants increase the risk of hepatocellular carcinoma in alcoholic cirrhosis. Dig. Liver Dis. 2016, 48, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Goossens, N.; Hoshida, Y. Is hepatocellular cancer the same disease in alcoholic and nonalcoholic fatty liver diseases? Gastroenterology 2016, 150, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Castano, G.O.; Scian, R.; Mallardi, P.; Fernandez Gianotti, T.; Burgueno, A.L.; San Martino, J.; Pirola, C.J. Genetic variation in transmembrane 6 superfamily member 2 and the risk of nonalcoholic fatty liver disease and histological disease severity. Hepatology 2015, 61, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Goffredo, M.; Caprio, S.; Feldstein, A.E.; D’Adamo, E.; Shaw, M.M.; Pierpont, B.; Savoye, M.; Zhao, H.; Bale, A.E.; Santoro, N. Role of TM6SF2 rs58542926 in the pathogenesis of nonalcoholic pediatric fatty liver disease: A multiethnic study. Hepatology 2016, 63, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Mancina, R.M.; Dongiovanni, P.; Petta, S.; Pingitore, P.; Meroni, M.; Rametta, R.; Boren, J.; Montalcini, T.; Pujia, A.; Wiklund, O.; et al. The MBOAT7-TMC4 variant rs641738 increases risk of nonalcoholic fatty liver disease in individuals of European descent. Gastroenterology 2016, 150, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Thabet, K.; Chan, H.L.Y.; Petta, S.; Mangia, A.; Berg, T.; Boonstra, A.; Brouwer, W.P.; Abate, M.L.; Wong, V.W.; Nazmy, M.; et al. The membrane-bound O-acyltransferase domain-containing 7 variant rs641738 increases inflammation and fibrosis in chronic hepatitis B. Hepatology 2017, 65, 1840–1850. [Google Scholar] [CrossRef] [PubMed]
- Meroni, M.; Longo, M.; Rametta, R.; Dongiovanni, P. Genetic and epigenetic modifiers of alcoholic liver disease. Int. J. Mol. Sci. 2018, 19, 3857. [Google Scholar] [CrossRef] [PubMed]
- Bataille, A.M.; Manautou, J.E. Nrf2: A potential target for new therapeutics in liver disease. Clin. Pharmacol. Ther. 2012, 92, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Xu, M.; Jeong, S.; Qian, Y.; Wu, H.; Xia, Q.; Kong, X. The role of Nrf2 in liver disease: Novel molecular mechanisms and therapeutic approaches. Front. Pharmacol. 2018, 9, 1428. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Guo, F.F.; Xie, K.Q.; Zeng, T. Targeting Nrf-2 is a promising intervention approach for the prevention of ethanol-induced liver disease. Cell. Mol. Life Sci. 2018, 75, 3143–3157. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.M.; Yang, J.H.; Ki, S.H. Role of the Nrf2-ARE pathway in liver diseases. Oxid. Med. Cell. Longev. 2013, 2013, 763257. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.H.; Shenvi, S.V.; Dixon, B.M.; Liu, H.; Jaiswal, A.K.; Liu, R.M.; Hagen, T.M. Decline in transcriptional activity of Nrf2 causes age-related loss of glutathione synthesis, which is reversible with lipoic acid. Proc. Natl. Acad. Sci. USA 2004, 101, 3381–3386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Marzec, J.M.; Christie, J.D.; Reddy, S.P.; Jedlicka, A.E.; Vuong, H.; Lanken, P.N.; Aplenc, R.; Yamamoto, T.; Yamamoto, M.; Cho, H.Y.; et al. Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J. 2007, 21, 2237–2246. [Google Scholar] [CrossRef] [PubMed]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Hasegawa, S.; Takagi, T.; Wang, F.Y.; et al. The relationship between Helicobacter pylori infection and promoter polymorphism of the Nrf2 gene in chronic gastritis. Int. J. Mol. Med. 2007, 19, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Arima, Y.; Okubo, M.; et al. Association between promoter polymorphisms of nuclear factor-erythroid 2-related factor 2 gene and peptic ulcer diseases. Int. J. Mol. Med. 2007, 20, 849–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Okubo, M.; Sakata, M.; et al. Nrf2 gene promoter polymorphism is associated with ulcerative colitis in a Japanese population. Hepato-gastroenterology 2008, 55, 394–397. [Google Scholar]
- Hua, C.C.; Chang, L.C.; Tseng, J.C.; Chu, C.M.; Liu, Y.C.; Shieh, W.B. Functional haplotypes in the promoter region of transcription factor Nrf2 in chronic obstructive pulmonary disease. Dis. Markers 2010, 28, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Von Otter, M.; Bergstrom, P.; Quattrone, A.; De Marco, E.V.; Annesi, G.; Soderkvist, P.; Wettinger, S.B.; Drozdzik, M.; Bialecka, M.; Nissbrandt, H.; et al. Genetic associations of Nrf2-encoding NFE2L2 variants with Parkinson’s disease—A multicenter study. BMC Med. Genet. 2014, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y.; Mitsuda, Y.; Tsuruta, Y.; Hamajima, N.; Niwa, T. Polymorphism of Nrf2, an antioxidative gene, is associated with blood pressure and cardiovascular mortality in hemodialysis patients. Int. J. Med. Sci. 2014, 11, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Zhang, L.; Lv, W.; Zhang, W.; Zhao, J.; Hu, X. NFE2L2 variations reduce antioxidant response in patients with Parkinson disease. Oncotarget 2016, 7, 10756–10764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, R.L.; Chung, G.G.; Rimm, D.L. Automated subcellular localization and quantification of protein expression in tissue microarrays. Nat. Med. 2002, 8, 1323–1328. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Allele | ALD n (%) | Control n (%) | p Value | ||
|---|---|---|---|---|---|
| A/A | 18 | 39.1 | 5 | 23.8 | 0.1457 |
| G/G | 9 | 19.6 | 9 | 42.9 | |
| A/G | 19 | 41.3 | 7 | 33.3 | |
| Total | 46 | 100.0 | 21 | 100 | |
| A/A | 18 | 39.1 | 5 | 23.8 | 0.2192 |
| G/G_A/G | 28 | 60.9 | 16 | 76.2 | |
| Total | 46 | 100.0 | 21 | 100 | |
| A/A_A/G | 37 | 80.4 | 12 | 57.1 | 0.0460 |
| G/G | 9 | 19.6 | 9 | 42.9 | |
| Total | 46 | 100.0 | 21 | 100 | |
| A | 55 | 59.8 | 17 | 40.5 | 0.0387 |
| G | 37 | 40.2 | 25 | 59.5 | |
| Total | 92 | 100.0 | 42 | 100 | |
| Allele | ALD n (%) | HCV n (%) | p Value | ||
|---|---|---|---|---|---|
| A/A | 18 | 39.1 | 2 | 14.3 | 0.0668 |
| G/G | 9 | 19.6 | 7 | 50.0 | |
| A/G | 19 | 41.3 | 5 | 35.7 | |
| Total | 46 | 100.0 | 14 | 100.0 | |
| A/A | 18 | 39.1 | 2 | 14.3 | 0.0746 |
| G/G_A/G | 28 | 60.9 | 12 | 85.7 | |
| Total | 46 | 100.0 | 14 | 100.0 | |
| A/A_A/G | 37 | 80.4 | 7 | 50.0 | 0.0345 |
| G/G | 9 | 19.6 | 7 | 50.0 | |
| Total | 46 | 100.0 | 14 | 100.0 | |
| A | 55 | 59.8 | 9 | 32.1 | 0.0106 |
| G | 37 | 40.2 | 19 | 67.9 | |
| Total | 92 | 100.0 | 28 | 100.0 | |
| Allele | HCV n (%) | Control n (%) | p Value | ||
|---|---|---|---|---|---|
| A/A | 2 | 14.3 | 5 | 23.81 | 0.7898 |
| G/G | 7 | 50.0 | 9 | 42.86 | |
| A/G | 5 | 35.7 | 7 | 33.33 | |
| Total | 14 | 100.0 | 21 | 100 | |
| A/A | 2 | 14.3 | 5 | 23.81 | 0.4995 |
| G/G_A/G | 12 | 85.7 | 16 | 76.19 | |
| Total | 14 | 100.0 | 21 | 100 | |
| A/A_A/G | 7 | 50.0 | 12 | 57.14 | 0.6846 |
| G/G | 7 | 50.0 | 9 | 42.86 | |
| Total | 14 | 100.0 | 21 | 100 | |
| A | 9 | 32.1 | 17 | 40.48 | 0.4832 |
| G | 19 | 67.9 | 25 | 59.52 | |
| Total | 28 | 100.0 | 42 | 100 | |
| Allele | ALD n (%) | HCV n (%) | p Value | ||
|---|---|---|---|---|---|
| A/G | 16 | 34.8 | 10 | 55.6 | 0.0531 |
| A/A | 11 | 23.9 | 6 | 33.3 | |
| G/G | 19 | 41.3 | 2 | 11.1 | |
| Total | 46 | 100.0 | 18 | 100.0 | |
| A/A | 11 | 23.9 | 6 | 33.3 | 0.6510 |
| A/G_G/G | 35 | 76,1 | 12 | 66.7 | |
| Total | 46 | 100.0 | 18 | 100.0 | |
| A/A_A/G | 27 | 58.7 | 16 | 88.9 | 0.0360 |
| G/G | 19 | 41.3 | 2 | 11.1 | |
| Total | 46 | 100.0 | 18 | 100.0 | |
| A | 38 | 41.3 | 22 | 61.1 | 0.0685 |
| G | 54 | 58.7 | 14 | 38.9 | |
| Total | 92 | 100.0 | 36 | 100.0 | |
| Allele | ALD n (%) | HCV n (%) | p Value | ||
|---|---|---|---|---|---|
| A/A | 10 | 22.7 | 4 | 28.6 | 0.8715 |
| G/G | 22 | 50.0 | 7 | 50.0 | |
| G/A | 12 | 27.3 | 3 | 21.4 | |
| Total | 44 | 100.0 | 14 | 100.0 | |
| A/A | 10 | 22.7 | 4 | 28.6 | 0.7245 |
| G/G_G/A | 34 | 77.3 | 10 | 71.4 | |
| Total | 44 | 100.0 | 14 | 100.0 | |
| A/A_G/A | 22 | 50.0 | 7 | 50.0 | 1.000 |
| G/G | 22 | 50.0 | 7 | 50.0 | |
| Total | 44 | 100.0 | 14 | 100.0 | |
| A | 32 | 36.4 | 11 | 39.3 | 0.9568 |
| G | 56 | 63.6 | 17 | 60.7 | |
| Total | 88 | 100.0 | 28 | 100.0 | |
| Allele | ALD - SNP rs35652124 (-214 A ã G) | |||||||||||||||
| M % | F % | Age | MELD | ALT | AST | GGT | TB | DB | IB | ALP | LDH | ALB | INR | UR | CR | |
| A/A | 94.45 | 5.55 | 53.61 (2.057) | 16.54 (1.101) | 46.25 (11.02) | 76.91 (19.03) | 114.75 (21.65) | 3.26 (0.3964) | 1.20 (0.2407) | 2.05 (0.3092) | 214.2 (39.73) | 460.23 (48.82) | 3.12 (0.1612) | 1.68 (0.0984) | 27.77 (2.699) | 1.04 (0.183) |
| G/G | 100 | 0 | 55.25 (3.304) | 16 (0.7559) | 51.71 (4.46) | 77.34 (13.1) | 212 (56.35) | 3.43 (0.5941) | 1.68 (0.505) | 1.79 (0.1932) | 191.3 (51.58) | 468.6 (57.1) | 3.07 (0.1686) | 1.48 (0.085) | 34.72 (4.387) | 0.95 (0.112) |
| A/G | 100 | 0 | 52.21 (2.924) | 17 (1.075) | 30.47 (5.188) | 39.93 (4.364) | 88.34 (27.72) | 3.36 (0.5608) | 1.42 (0.4408) | 2.04 (0.2102) | 125.63 (16.44) | 421.02 (46.45) | 3.25 (0.1566) | 1.82 (0.1526) | 38.04 (3.144) | 0.99 (0.068) |
| p value | - | - | 0.7761 | 0.8396 | 0.2224 | 0.1226 | 0.0514 | 0.9742 | 0.6619 | 0.7853 | 0.1928 | 0.8082 | 0.7287 | 0.2295 | 0.0643 | 0.9129 |
| Allele | ALD - SNP rs4893819 (-1275 G ã A) | |||||||||||||||
| M % | F % | Age | MELD | ALT | AST | GGT | TB | DB | IB | ALP | LDH | ALB | INR | UR | CR | |
| A/A | 88.89 | 11.11 | 48.33 (2.809) | 16.66 (0.8028) | 46.2 (5.275) | 60.05 (11.8) | 112.63 (39.07) | 3.26 (0.439) | 1.3 (0.2762) | 2 (0.2608) | 159.2 (23.81) | 421.44 (40.07) | 3.01 (0.04) | 1.56 (0.0898) | 26.76 (5.056) | 1.36 (0.321) |
| G/G | 100 | 0 | 53 (2.282) | 16.71 (1.029) | 44.87 (8.916) | 70.73 (15.51) | 118.7 (20.65) | 3.24 (0.417) | 1.2 (0.2856) | 2.03 (0.2483) | 151.06 (18.09) | 468.6 (47.46) | 3.10 (0.149) | 1.81 (0.1171) | 29.85 (3.431) | 0.9 (0.074) |
| G/A | 100 | 0 | 59 (2.967) | 16.86 (1.487) | 36 (10.75) | 55.67 (15.92) | 180 (79.48) | 3.71 (0.5114) | 1.63 (0.4826) | 1.89 (0.3667) | 268.3 (79.75) | 443.6 (75.63) | 3.47 (0.2327) | 1.56 (0.1429) | 35.45 (5.895) | 1.51 (0.107) |
| p value | - | - | 0.0563 | 0.9946 | 0.7944 | 0.8009 | 0.5107 | 0.7671 | 0.7181 | 0.9516 | 0.1054 | 0.8480 | 0.2425 | 0.2825 | 0.6952 | 0.1259 |
| Allele | ALD - SNP rs6721961 (-178 A ã G) | |||||||||||||||
| M % | F % | Age | MELD | ALT | AST | GGT | TB | DB | IB | ALP | LDH | ALB | INR | UR | CR | |
| A/A | 90 | 10 | 56.5 (3.212) | 16.63 (1.101) | 39.5 (7.058) | 72.75 (10.37) | 158.71 (60.2) | 3.64 (0.3599) | 1.63 (0.3632) | 2.03 (0.3692) | 267.62 (68.49) | 381.42 (58.18) | 3.3 (0.184) | 1.5 (0.1257) | 37 (4.687) | 1.32 (0.269) |
| G/G | 100 | 0 | 49.71 (1.957) | 15 (0.7977) | 40.18 (5.564) | 58.94 (8.13) | 115.7 (21.53) | 2.63 (0.3742) | 0.93 (0.1771) | 1.87 (0.2752) | 146.1 (16.06) | 471.01 (49.53) | 3.07 (0.1274) | 1.7 (0.107) | 32.04 (3.942) | 0.84 (0.051) |
| A/G | 100 | 0 | 57.14 (2.344) | 17.36 (1.002) | 43.73 (11.83) | 59.85 (21.59) | 120.5 (33.98) | 3.74 (0.5585) | 1.69 (0.4554) | 1.92 (0.181) | 126.5 (19.45) | 470.8 (47.57) | 3.18 (0.198) | 1.68 (1.1344) | 34.19 (2.451) | 0.97 (0.086) |
| p value | - | - | 0.0482 | 0.1959 | 0.9359 | 0.7991 | 0.7017 | 0.1677 | 0.2381 | 0.9141 | 0.0235 | 0.4332 | 0.6492 | 0.5050 | 0.6612 | 0.0623 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes dos Santos, K.; Florentino, R.M.; França, A.; Lima Filho, A.C.M.; Santos, M.L.d.; Missiaggia, D.; Fonseca, M.d.C.; Brasil Costa, I.; Vidigal, P.V.T.; Nathanson, M.H.; et al. Polymorphism in the Promoter Region of NFE2L2 Gene Is a Genetic Marker of Susceptibility to Cirrhosis Associated with Alcohol Abuse. Int. J. Mol. Sci. 2019, 20, 3589. https://doi.org/10.3390/ijms20143589
Nunes dos Santos K, Florentino RM, França A, Lima Filho ACM, Santos MLd, Missiaggia D, Fonseca MdC, Brasil Costa I, Vidigal PVT, Nathanson MH, et al. Polymorphism in the Promoter Region of NFE2L2 Gene Is a Genetic Marker of Susceptibility to Cirrhosis Associated with Alcohol Abuse. International Journal of Molecular Sciences. 2019; 20(14):3589. https://doi.org/10.3390/ijms20143589
Chicago/Turabian StyleNunes dos Santos, Kemper, Rodrigo M. Florentino, Andressa França, Antônio Carlos Melo Lima Filho, Marcone Loiola dos Santos, Dabny Missiaggia, Matheus de Castro Fonseca, Igor Brasil Costa, Paula Vieira Teixeira Vidigal, Michael H. Nathanson, and et al. 2019. "Polymorphism in the Promoter Region of NFE2L2 Gene Is a Genetic Marker of Susceptibility to Cirrhosis Associated with Alcohol Abuse" International Journal of Molecular Sciences 20, no. 14: 3589. https://doi.org/10.3390/ijms20143589
APA StyleNunes dos Santos, K., Florentino, R. M., França, A., Lima Filho, A. C. M., Santos, M. L. d., Missiaggia, D., Fonseca, M. d. C., Brasil Costa, I., Vidigal, P. V. T., Nathanson, M. H., Lemos, F. d. O., & Leite, M. F. (2019). Polymorphism in the Promoter Region of NFE2L2 Gene Is a Genetic Marker of Susceptibility to Cirrhosis Associated with Alcohol Abuse. International Journal of Molecular Sciences, 20(14), 3589. https://doi.org/10.3390/ijms20143589