Secretome of Adipose Tissue-Derived Stem Cells (ASCs) as a Novel Trend in Chronic Non-Healing Wounds: An Overview of Experimental In Vitro and In Vivo Studies and Methodological Variables


 ,
, 
 ,
,
Abstract
:
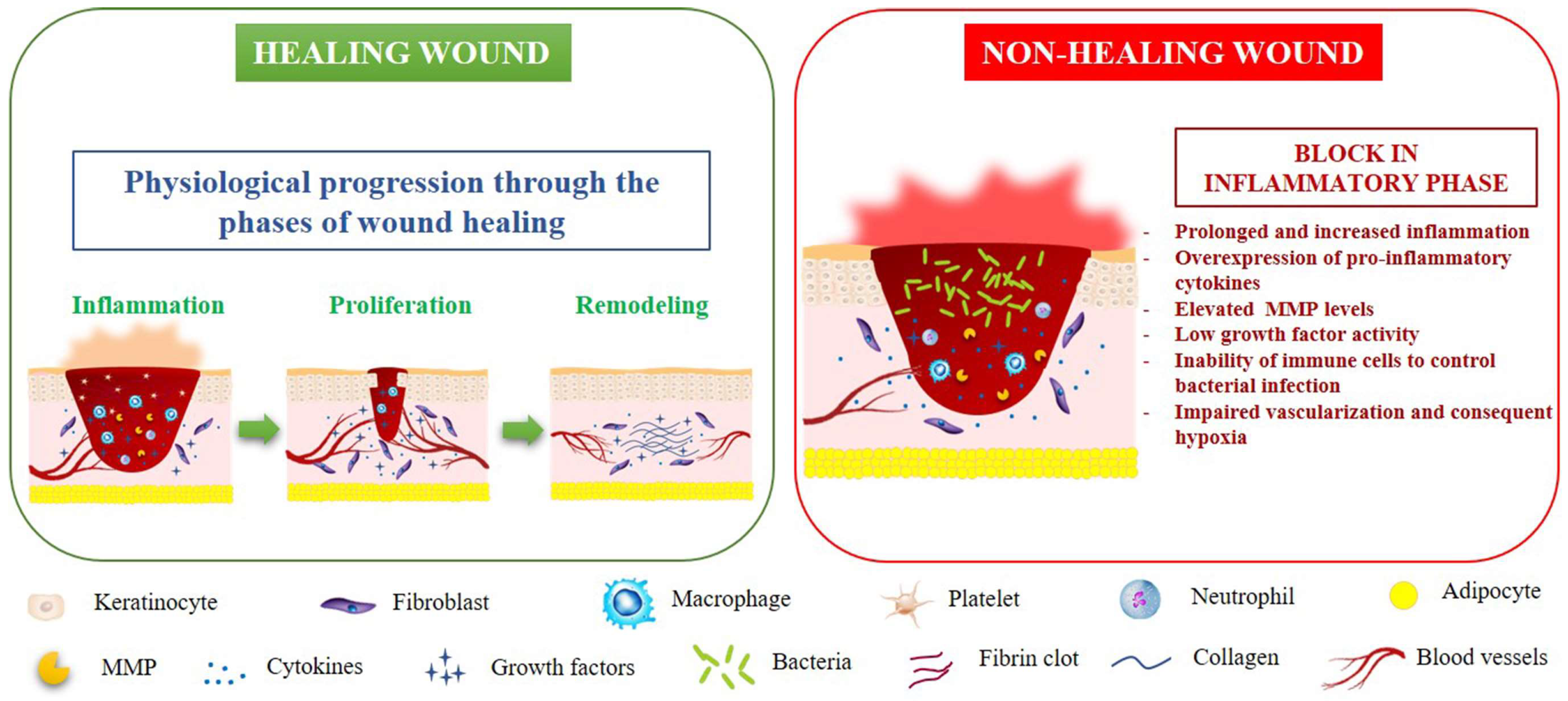
1. Introduction
2. Effects of ASC-Conditioned Medium/Secretome on Wound Healing
2.1. In Vitro Studies
2.2. In Vivo Experimental Studies
3. Effects of ASC-Secreted Extracellular Vesicles on Wound Healing
3.1. In Vitro Studies
3.2. In Vivo Studies
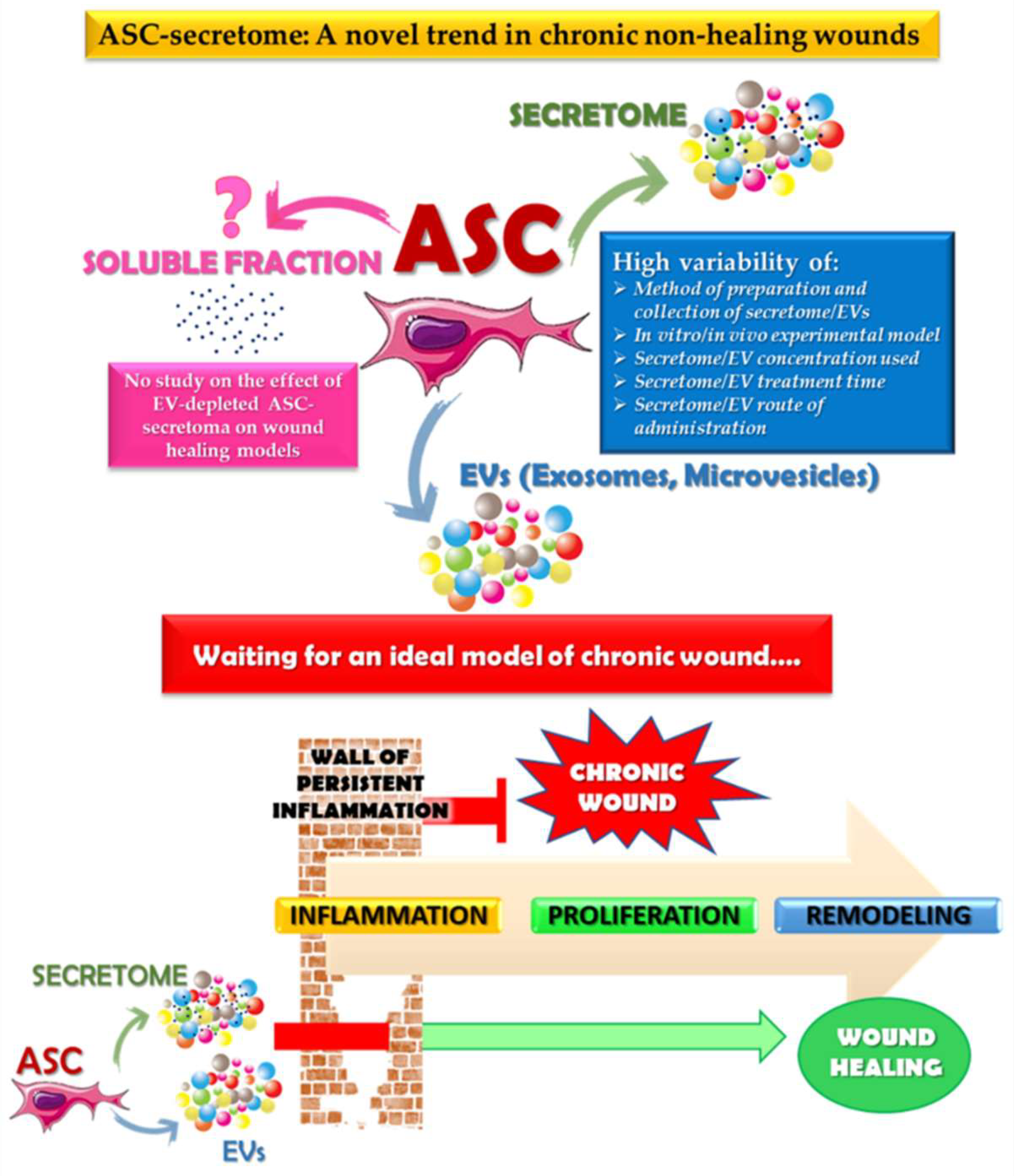
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MMP | Metalloproteinase |
| ECM | Extracellular matrix |
| MSCs | Mesenchymal stem cells |
| ASCs | Adipose tissue-derived stem cells |
| CM | Conditioned medium |
| EVs | Extracellular vesicles |
| HDFs | Human dermal fibroblasts |
| HUVECs | Umbilical vein endothelial cells |
| MVs | Microvesicles |
References
- Chan, B.; Cadarette, S.; Wodchis, W.; Wong, J.; Mittmann, N.; Krahn, M. Cost-of-illness studies in chronic ulcers: A systematic review. J. Wound Care 2017, 26, S4–S14. [Google Scholar] [CrossRef] [PubMed]
- Jarbrink, K.; Ni, G.; Sonnergren, H.; Schmidtchen, A.; Pang, C.; Bajpai, R.; Car, J. Prevalence and incidence of chronic wounds and related complications: A protocol for a systematic review. Syst. Rev. 2016, 5, 152. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, S.R.; Carter, M.J.; Fife, C.E.; DaVanzo, J.; Haught, R.; Nusgart, M.; Cartwright, D. An economic evaluation of the impact, cost, and medicare policy implications of chronic nonhealing wounds. Value Health J. Int. Soc. Pharm. Outcomes Res. 2018, 21, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Gould, L.; Abadir, P.; Brem, H.; Carter, M.; Conner-Kerr, T.; Davidson, J.; DiPietro, L.; Falanga, V.; Fife, C.; Gardner, S.; et al. Chronic wound repair and healing in older adults: Current status and future research. J. Am. Geriatr. Soc. 2015, 63, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Wicke, C.; Bachinger, A.; Coerper, S.; Beckert, S.; Witte, M.B.; Konigsrainer, A. Aging influences wound healing in patients with chronic lower extremity wounds treated in a specialized wound care center. Wound Repair Regen. 2009, 17, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.L.; Liang, H.L.N.; Clarke, E.; Jackson, C.; Xue, M.L. Inflammation in chronic wounds. Int. J. Mol. Sci. 2016, 17, 2085. [Google Scholar] [CrossRef]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound healing: A cellular perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune regulation of skin wound healing: Mechanisms and novel therapeutic targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Mustoe, T.A.; O’Shaughnessy, K.; Kloeters, O. Chronic wound pathogenesis and current treatment strategies: A unifying hypothesis. Plast. Reconstr. Surg. 2006, 117, 35S–41S. [Google Scholar] [CrossRef]
- Han, G.; Ceilley, R. Chronic wound healing: A review of current management and treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef]
- Opdenakker, G.; Van Damme, J.; Vranckx, J.J. Immunomodulation as rescue for chronic atonic skin wounds. Trends Immunol. 2018, 39, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, V.K.; Angra, D.; Rahimi, H.; McNish, S. Vasculitic and autoimmune wounds. J. Vasc. Surg. Venous Lymphat. Disord. 2017, 5, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Steen, V.; Denton, C.P.; Pope, J.E.; Matucci-Cerinic, M. Digital ulcers: Overt vascular disease in systemic sclerosis. Rheumatology 2009, 48, iii19–iii24. [Google Scholar] [CrossRef] [PubMed]
- Frykberg, R.G.; Banks, J. Challenges in the treatment of chronic wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef] [PubMed]
- Kirby, G.T.S.; Mills, S.J.; Cowin, A.J.; Smith, L.E. Stem cells for cutaneous wound healing. BioMed Res. Int. 2015, 2015, 285869. [Google Scholar] [CrossRef]
- Kucharzewski, M.; Rojczyk, E.; Wilemska-Kucharzewska, K.; Wilk, R.; Hudecki, J.; Los, M.J. Novel trends in application of stem cells in skin wound healing. Eur. J. Pharm. 2019, 843, 307–315. [Google Scholar] [CrossRef]
- Leavitt, T.; Hu, M.S.; Marshall, C.D.; Barnes, L.A.; Longaker, M.T.; Lorenz, H.P. Stem cells and chronic wound healing: State of the art. Chronic Wound Care Manag. Res. 2016, 3, 7–27. [Google Scholar]
- Marfia, G.; Navone, S.E.; Di Vito, C.; Ughi, N.; Tabano, S.; Miozzo, M.; Tremolada, C.; Bolla, G.; Crotti, C.; Ingegnoli, F.; et al. Mesenchymal stem cells: Potential for therapy and treatment of chronic non-healing skin wounds. Organogenesis 2015, 11, 183–206. [Google Scholar] [CrossRef]
- Del Papa, N.; Caviggioli, F.; Sambataro, D.; Zaccara, E.; Vinci, V.; Di Luca, G.; Parafioriti, A.; Armiraglio, E.; Maglione, W.; Polosa, R.; et al. Autologous fat grafting in the treatment of fibrotic perioral changes in patients with systemic sclerosis. Cell Transpl. 2015, 24, 63–72. [Google Scholar] [CrossRef]
- Del Papa, N.; Di Luca, G.; Andracco, R.; Zaccara, E.; Maglione, W.; Pignataro, F.; Minniti, A.; Vitali, C. Regional grafting of autologous adipose tissue is effective in inducing prompt healing of indolent digital ulcers in patients with systemic sclerosis: Results of a monocentric randomized controlled study. Arthritis Res. 2019, 21, 7. [Google Scholar] [CrossRef]
- Toyserkani, N.M.; Christensen, M.L.; Sheikh, S.P.; Sorensen, J.A. Adipose-derived stem cells: New treatment for wound healing? Ann. Plast. Surg. 2015, 75, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Kocan, B.; Maziarz, A.; Tabarkiewicz, J.; Ochiya, T.; Banas-Zabczyk, A. Trophic activity and phenotype of adipose tissue-derived mesenchymal stem cells as a background of their regenerative potential. Stem Cells Int. 2017, 2017, 1653254. [Google Scholar] [CrossRef] [PubMed]
- Brzoska, M.; Geiger, H.; Gauer, S.; Baer, P. Epithelial differentiation of human adipose tissue-derived adult stem cells. Biochem. Biophys. Res. Commun. 2005, 330, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Chiou, J.F.; Wu, A.T.H.; Tsai, C.Y.; Leu, J.D.; Ting, L.L.; Wang, M.F.; Chen, H.Y.; Lin, C.T.; Williams, D.F.; et al. The effect of diminished osteogenic signals on reduced osteoporosis recovery in aged mice and the potential therapeutic use of adipose-derived stem cells. Biomaterials 2012, 33, 6105–6112. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Scutaru, T.T.; Ghetu, N.; Carasevici, E.; Lupascu, C.D.; Ferariu, D.; Pieptu, D.; Coman, C.G.; Danciu, M. The effects of adipose-derived stem cell-differentiated adipocytes on skin burn wound healing in rats. J. Burn Care Res. 2017, 38, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cherubino, M.; Rubin, J.P.; Miljkovic, N.; Kelmendi-Doko, A.; Marra, K.G. Adipose-derived stem cells for wound healing applications. Ann. Plast. Surg. 2011, 66, 210–215. [Google Scholar] [CrossRef]
- Kim, W.S.; Park, B.S.; Sung, J.H. The wound-healing and antioxidant effects of adipose-derived stem cells. Expert Opin. Biol. Ther. 2009, 9, 879–887. [Google Scholar] [CrossRef]
- Li, P.; Guo, X.T. A review: Therapeutic potential of adipose-derived stem cells in cutaneous wound healing and regeneration. Stem Cell Res. Ther. 2018, 9, 302. [Google Scholar] [CrossRef]
- McLaughlin, M.M.; Marra, K.G. The use of adipose-derived stem cells as sheets for wound healing. Organogenesis 2013, 9, 79–81. [Google Scholar] [CrossRef] [Green Version]
- Zeppieri, M.; Salvetat, M.L.; Beltrami, A.; Cesselli, D.; Russo, R.; Alcalde, I.; Merayo-Lloves, J.; Brusini, P.; Parodi, P.C. Adipose derived stem cells for corneal wound healing after laser induced corneal lesions in mice. J. Clin. Med. 2017, 6, 115. [Google Scholar] [CrossRef]
- Hu, M.S.; Borrelli, M.R.; Lorenz, H.P.; Longaker, M.T.; Wan, D.C. Mesenchymal stromal cells and cutaneous wound healing: A comprehensive review of the background, role, and therapeutic potential. Stem Cells Int. 2018, 2018, 6901983. [Google Scholar] [CrossRef] [PubMed]
- Otero-Vinas, M.; Falanga, V. Mesenchymal stem cells in chronic wounds: The spectrum from basic to advanced therapy. Adv. Wound Care 2016, 5, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, P.; Lombardi, F.; Siragusa, G.; Cifone, M.G.; Cinque, B.; Giuliani, M. Methods of isolation, characterization and expansion of human adipose-derived stem cells (ascs): An overview. Int. J. Mol. Sci. 2018, 19, 1897. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, P.; Miconi, G.; Cinque, B.; La Torre, C.; Lombardi, F.; Zoccali, G.; Orsini, G.; Leocata, P.; Giuliani, M.; Cifone, M.G. In vitro evaluation of different methods of handling human liposuction aspirate and their effect on adipocytes and adipose derived stem cells. J. Cell. Physiol. 2015, 230, 1974–1981. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, J.; Sensebe, L. Mesenchymal stromal cells: Clinical challenges and therapeutic opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Neri, S. Genetic stability of mesenchymal stromal cells for regenerative medicine applications: A fundamental biosafety aspect. Int. J. Mol. Sci. 2019, 20, 2406. [Google Scholar] [CrossRef]
- Agrawal, H.; Shang, H.L.; Sattah, A.P.; Yang, N.; Peirce, S.M.; Katz, A.J. Human adipose-derived stromal/stem cells demonstrate short-lived persistence after implantation in both an immunocompetent and an immunocompromised murine model. Stem Cell Res. Ther. 2014, 5, 142. [Google Scholar] [CrossRef]
- Dubey, N.K.; Mishra, V.K.; Dubey, R.; Deng, Y.H.; Tsai, F.C.; Deng, W.P. Revisiting the advances in isolation, characterization and secretome of adipose-derived stromal/stem cells. Int. J. Mol. Sci. 2018, 19, 2200. [Google Scholar] [CrossRef]
- Frese, L.; Dijkman, P.E.; Hoerstrup, S.P. Adipose tissue-derived stem cells in regenerative medicine. Transfus. Med. Hemother. 2016, 43, 268–274. [Google Scholar] [CrossRef]
- Gimble, J.M.; Bunnell, B.A.; Guilak, F. Human adipose-derived cells: An update on the transition to clinical translation. Regen. Med. 2012, 7, 225–235. [Google Scholar] [CrossRef]
- Kapur, S.K.; Katz, A.J. Review of the adipose derived stem cell secretome. Biochimie 2013, 95, 2222–2228. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.T.; Dilley, R.J. Novel non-angiogenic role for mesenchymal stem cell-derived vascular endothelial growth factor on keratinocytes during wound healing. Cytokine Growth Factor Rev. 2018, 44, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Doruk, O.; Mizuno, H. Multipotency and secretome: The mechanisms behind the regenerative potential of adipose-derived stem cells. Plast. Aesthetic Res. 2017, 4, 32–40. [Google Scholar]
- Kumar, L.P.; Kandoi, S.; Misra, R.; Vijayalakshmi, S.; Rajagopal, K.; Verma, R.S. The mesenchymal stem cell secretome: A new paradigm towards cell-free therapeutic mode in regenerative medicine. Cytokine Growth Factor Rev. 2019, 46, 1–9. [Google Scholar]
- Maguire, G. Stem cell therapy without the cells. Commun. Integr. Biol. 2013, 6, e26631. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Kim, J.W.; Jun, H.S.; Roh, J.Y.; Lee, H.Y.; Hong, I.S. Stem cell secretome and its effect on cellular mechanisms relevant to wound healing. Mol. Ther. 2018, 26, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.R.; Xu, Y.; Guo, S.L.; Xu, Y.; Wang, Y.; Zhu, F.; Permatasari, F.; Wu, D.; Yin, Z.Q.; Luo, D. The effect of conditioned media of adipose-derived stem cells on wound healing after ablative fractional carbon dioxide laser resurfacing. Biomed Res. Int. 2013, 2013, 519126. [Google Scholar] [CrossRef]
- Campanella, C.; Bavisotto, C.C.; Logozzi, M.; Gammazza, A.M.; Mizzoni, D.; Cappello, F.; Fais, S. On the choice of the extracellular vesicles for therapeutic purposes. Int. J. Mol. Sci. 2019, 20, 236. [Google Scholar] [CrossRef]
- Ferreira, A.D.F.; Gomes, D.A. Stem cell extracellular vesicles in skin repair. Bioengineering 2018, 6, 4. [Google Scholar] [CrossRef]
- Pelizzo, G.; Avanzini, M.A.; Cornaglia, A.I.; De Silvestri, A.; Mantelli, M.; Travaglino, P.; Croce, S.; Romano, P.; Avolio, L.; Iacob, G.; et al. Extracellular vesicles derived from mesenchymal cells: Perspective treatment for cutaneous wound healing in pediatrics. Regen. Med. 2018, 13, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of adipose-derived mesenchymal stem cells promotes skeletal muscle regeneration through synergistic action of extracellular vesicle cargo and soluble proteins. Stem Cell Res. Ther. 2019, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Goncalves, R.M. Mesenchymal stromal cell secretome: Influencing therapeutic potential by cellular pre-conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef] [PubMed]
- Kyurkchiev, D.; Bochev, I.; Ivanova-Todorova, E.; Mourdjeva, M.; Oreshkova, T.; Belemezova, K.; Kyurkchiev, S. Secretion of immunoregulatory cytokines by mesenchymal stem cells. World J. Stem Cells 2014, 6, 552–570. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, M.; Rao, K.S.; Riordan, N.H. A review of therapeutic effects of mesenchymal stem cell secretions and induction of secretory modification by different culture methods. J. Transl. Med. 2014, 12, 260. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Park, B.S.; Sung, J.H.; Yang, J.M.; Park, S.B.; Kwak, S.J.; Park, J.S. Wound healing effect of adipose-derived stem cells: A critical role of secretory factors on human dermal fibroblasts. J. Dermatol. Sci. 2007, 48, 15–24. [Google Scholar] [CrossRef]
- Zhao, J.; Hu, L.; Liu, J.; Gong, N.; Chen, L. The effects of cytokines in adipose stem cell-conditioned medium on the migration and proliferation of skin fibroblasts in vitro. BioMed Res. Int. 2013, 2013, 578479. [Google Scholar] [CrossRef]
- Kober, J.; Gugerell, A.; Schmid, M.; Zeyda, M.; Buchberger, E.; Nickl, S.; Hacker, S.; Ankersmit, H.J.; Keck, M. Wound healing effect of conditioned media obtained from adipose tissue on human skin cells a comparative in vitro study. Ann. Plast. Surg. 2016, 77, 156–163. [Google Scholar] [CrossRef]
- Collawn, S.S.; Mobley, J.A.; Banerjee, N.S.; Chow, L.T. Conditioned media from adipose-derived stromal cells accelerates healing in 3-dimensional skin cultures. Ann. Plast. Surg. 2016, 76, 446–452. [Google Scholar] [CrossRef]
- Seo, E.; Lim, J.S.; Jun, J.B.; Choi, W.; Hong, I.S.; Jun, H.S. Exendin-4 in combination with adipose-derived stem cells promotes angiogenesis and improves diabetic wound healing. J. Transl. Med. 2017, 15, 35. [Google Scholar] [CrossRef]
- Cooper, D.R.; Wang, C.Y.; Patel, R.; Trujillo, A.; Patel, N.A.; Prather, J.; Gould, L.J.; Wu, M.H. Human adipose-derived stem cell conditioned media and exosomes containing malat1 promote human dermal fibroblast migration and ischemic wound healing. Adv. Wound Care 2018, 7, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Wu, W.H.; Choi, J.H.; Kim, J.; Jun, J.H.; Ko, Y.; Lee, J.H. Galectin-1 from conditioned medium of three-dimensional culture of adipose-derived stem cells accelerates migration and proliferation of human keratinocytes and fibroblasts. Wound Repair Regen. 2018, 26, S9–S18. [Google Scholar] [CrossRef] [PubMed]
- Noverina, R.; Widowati, W.; Ayuningtyas, W.; Kurniawan, D.; Afifah, E.; Laksmitawati, D.R.; Rinendyaputri, R.; Rilianawati, R.; Faried, A.; Bachtiar, I.; et al. Growth factors profile in conditioned medium human adipose tissue-derived mesenchymal stem cells (cm-hatmscs). Clin. Nutr. Exp. 2019, 24, 34–44. [Google Scholar] [CrossRef]
- Stojanovic, S.; Najman, S. The effect of conditioned media of stem cells derived from lipoma and adipose tissue on macrophages’ response and wound healing in indirect co-culture system in vitro. Int. J. Mol. Sci. 2019, 20, 1671. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.C.; Jeon, E.S.; Lee, I.H.; Kim, H.S.; Kim, M.B.; Kim, J.H. Tumor necrosis factor-alpha-activated human adipose tissue-derived mesenchymal stem cells accelerate cutaneous wound healing through paracrine mechanisms. J. Investig. Dermatol. 2011, 131, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Kim, J.; Kim, M.Y.; Bae, Y.S.; Ryu, S.H.; Lee, T.G.; Kim, J.H. Proteomic analysis of tumor necrosis factor-alpha-induced secretome of human adipose tissue-derived mesenchymal stem cells. J. Proteome Res. 2010, 9, 1754–1762. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Gao, P.L.; Wang, K.; Wang, J.Y.; Zhong, Y.; Luo, Y. Fibrous scaffolds potentiate the paracrine function of mesenchymal stem cells: A new dimension in cell-material interaction. Biomaterials 2017, 141, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Irons, R.F.; Cahill, K.W.; Rattigan, D.A.; Marcotte, J.H.; Fromer, M.W.; Chang, S.; Zhang, P.; Behling, E.M.; Behling, K.C.; Caputo, F.J. Acceleration of diabetic wound healing with adipose-derived stem cells, endothelial-differentiated stem cells, and topical conditioned medium therapy in a swine model. J. Vasc. Surg. 2018, 68, 115S–125S. [Google Scholar] [CrossRef]
- Hartjes, T.A.; Mytnyk, S.; Jenster, G.W.; van Steijn, V.; van Royen, M.E. Extracellular vesicle quantification and characterization: Common methods and emerging approaches. Bioengineering 2019, 6, 7. [Google Scholar] [CrossRef]
- Raik, S.; Kumar, A.; Bhattacharyya, S. Insights into cell-free therapeutic approach: Role of stem cell “soup-ernatant”. Biotechnol. Appl. Biochem. 2018, 65, 104–118. [Google Scholar] [CrossRef]
- Toh, W.S.; Lai, R.C.; Zhang, B.; Lim, S.K. Msc exosome works through a protein-based mechanism of action. Biochem. Soc. Trans. 2018, 46, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (misev2018): A position statement of the international society for extracellular vesicles and update of the misev2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, T.; Bruno, S.; Tetta, C.; Kalinina, N.; Porta, M.; Camussi, G. Platelet-derived growth factor regulates the secretion of extracellular vesicles by adipose mesenchymal stem cells and enhances their angiogenic potential. Cell Commun. Signal. 2014, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, J.; Zhou, X.; Xiong, Z.; Zhao, J.; Yu, R.; Huang, F.; Zhang, H.; Chen, L. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci. Rep. 2016, 6, 32993. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.L.; Shen, Y.M.; Li, X.C.; Li, B.; Zhao, S.; Gu, J.J.; Chen, Y.F.; Ma, B.T.; Wei, J.J.; Han, Q.; et al. Exosomes derived from hypoxia-treated human adipose mesenchymal stem cells enhance angiogenesis through the pka signaling pathway. Stem Cells Dev. 2018, 27, 456–465. [Google Scholar] [CrossRef]
- Ferreira, A.D.F.; Cunha, P.D.S.; Carregal, V.M.; da Silva, P.C.; de Miranda, M.C.; Kunrath-Lima, M.; de Melo, M.I.A.; Faraco, C.C.F.; Barbosa, J.L.; Frezard, F.; et al. Extracellular vesicles from adipose-derived mesenchymal stem/stromal cells accelerate migration and activate akt pathway in human keratinocytes and fibroblasts independently of mir-205 activity. Stem Cells Int. 2017, 2017, 9841035. [Google Scholar] [CrossRef]
- Xue, G.; Hemmings, B.A. Pkb/akt-dependent regulation of cell motility. J. Natl. Cancer Inst. 2013, 105, 393–404. [Google Scholar] [CrossRef]
- Zhang, W.; Bai, X.; Zhao, B.; Li, Y.; Zhang, Y.; Li, Z.; Wang, X.; Luo, L.; Han, F.; Zhang, J.; et al. Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the pi3k/akt signaling pathway. Exp. Cell Res. 2018, 370, 333–342. [Google Scholar] [CrossRef]
- Ren, S.; Chen, J.; Duscher, D.; Liu, Y.T.; Guo, G.J.; Kang, Y.; Xiong, H.W.; Zhan, P.; Wang, Y.; Wang, C.; et al. Microvesicles from human adipose stem cells promote wound healing by optimizing cellular functions via akt and erk signaling pathways. Stem Cell Res. Ther. 2019, 10, 47. [Google Scholar] [CrossRef]
- Choi, E.W.; Seo, M.K.; Woo, E.Y.; Kim, S.H.; Park, E.J.; Kim, S. Exosomes from human adipose-derived stem cells promote proliferation and migration of skin fibroblasts. Exp. Derm. 2018, 27, 1170–1172. [Google Scholar] [CrossRef]
- Li, X.; Xie, X.Y.; Lian, W.S.; Shi, R.F.; Han, S.L.; Zhang, H.J.; Lu, L.G.; Li, M.Q. Exosomes from adipose-derived stem cells overexpressing nrf2 accelerate cutaneous wound healing by promoting vascularization in a diabetic foot ulcer rat model. Exp. Mol. Med. 2018, 50, 29. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Rojo de la Vega, M.; Wen, Q.; Bharara, M.; Jiang, T.; Zhang, R.; Zhou, S.; Wong, P.K.; Wondrak, G.T.; Zheng, H.; et al. An essential role of nrf2 in diabetic wound healing. Diabetes 2016, 65, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Fu, B.; Yang, X.; Xiao, Y.; Pan, M. Adipose mesenchymal stem cell-derived exosomes promote cell proliferation, migration, and inhibit cell apoptosis via wnt/beta-catenin signaling in cutaneous wound healing. J. Cell. Biochem. 2019, 120, 10847–10854. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.S.; Wei, Q.; Gurung, A.; Youn, A.; Bright, T.; Poon, R.; Whetstone, H.; Guha, A.; Alman, B.A. Beta-catenin regulates wound size and mediates the effect of tgf-beta in cutaneous healing. FASEB J. 2006, 20, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, M.; Gong, A.; Zhang, X.; Wu, X.; Zhu, Y.; Shi, H.; Wu, L.; Zhu, W.; Qian, H.; et al. Hucmsc-exosome mediated-wnt4 signaling is required for cutaneous wound healing. Stem Cells 2015, 33, 2158–2168. [Google Scholar] [CrossRef]
- Sagaradze, G.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Kalinina, N.; Akopyan, Z.; Efimenko, A. Conditioned medium from human mesenchymal stromal cells: Towards the clinical translation. Int. J. Mol. Sci. 2019, 20, 1656. [Google Scholar] [CrossRef]
- Park, S.; Lee, D.R.; Nam, J.S.; Ahn, C.W.; Kim, H. Fetal bovine serum-free cryopreservation methods for clinical banking of human adipose-derived stem cells. Cryobiology 2018, 81, 65–73. [Google Scholar] [CrossRef]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of extracellular vesicles: General methodologies and latest trends. Biomed Res. Int. 2018, 2018, 8545347. [Google Scholar] [CrossRef]
- Grada, A.; Mervis, J.; Falanga, V. Research techniques made simple: Animal models of wound healing. J. Investig. Dermatol. 2018, 138, 2095–2105. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| ASC Source | Model | Effect | Reference |
|---|---|---|---|
| Human lipoaspirates from healthy females. | HDF | Increased cell proliferation and migration. Higher type I collagen secretion. | [56] |
| Human lipoaspirates from female patients (18–35 years old). | HDF | Increased cell proliferation and migration. | [57] |
| Human adipose tissue from donors undergoing abdominoplasty. | HDF | No effect. | [58] |
| HUVEC | No effect. | ||
| Human keratinocytes | Reduction of keratinocyte proliferation. | ||
| Human abdominal adipose tissue and lipoaspirates from patients undergoing panniculectomy or abdominoplasty. | 3-D skin cultures of primary human keratinocytes | Acceleration of wound closure. | [59] |
| Commercial human ASCs. | HUVEC | Increased cell proliferation, migration, and invasion ability. | [60] |
| HaCaT | Increased cell proliferation, migration, and VEGF secretion. | ||
| Commercial human ASCs of normal donors (25–45 years old) undergoing elective surgery. | HDF | Increased cell migration. | [61] |
| Adipose tissue of breast cancer patients. | HDF | Increased cell proliferation, migration, and invasion ability. | [46] |
| HEKa | |||
| HUVEC | |||
| Human sub-cutaneous adipose tissue from male patient (51-year-old) undergoing skin-graft surgery. | HDF | Increased cell proliferation and migration. | [62] |
| HaCaT | |||
| Sub-cutaneous adipose tissue (average age of patients 47.2 ± 10.8) or sub-cutaneous lipoma (average age of patients 41.8 ± 7.1). | RAW 264.7 | Functional cell activation and M2 phenotype polarization. | [64] |
| L929 Fibroblasts | Acceleration of scratched monostrate closure, increased cell migration. |
| ASC Source | Model | Effect | Reference |
|---|---|---|---|
| Human sub-cutaneous adipose tissue. | Rat skin excisional wound model | Stimulation of wound closure, angiogenesis, proliferation, and macrophage infiltration. | [65] |
| Commercial rat ASC. | Rat skin excisional wound model | Accelerated wound closure, increased macrophage recruitment and M2 phenotype polarization. | [67] |
| Adipose tissue of breast cancer patients. | Nude mice full-thickness excisional skin wound model | Stimulation of wound closure, dermal thickening, angiogenesis, and immune cell recruitment. | [46] |
| Commercial human ASCs of normal donors (25–45 years old) undergoing elective surgery. | Rat ischemic skin wound model | Acceleration of wound closure. | [61] |
| Gluteal region of Yorkshire pigs. | Yorkshire pig diabetic skin wound model | Acceleration of wound closure, increased angiogenesis, reduced inflammatory profile. | [68] |
| ASC Source | Model | Effect | Reference |
|---|---|---|---|
| Commercial human ASC | HMEC | Stimulation of vessel-like structure formation | [73] |
| Human sub-cutaneous adipose tissue obtained from healthy females (18–35 years old). | HDF | Stimulation of cell migration and proliferation. Higher collagen synthesis. | [74] |
| Human sub-cutaneous adipose tissue of normal donors (25–45 years old) undergoing elective surgery. | HDF | Increase of cell migration rate. | [61] |
| Adult fat samples. | HUVEC | Stimulation of angiogenesis, upregulation and downregulation of angiogenesis-stimulating or -inhibitory genes, respectively. | [75] |
| Human lipoaspirates. | Primary human keratinocytes HDF | Increased cell proliferation and migration rate. Upregulated Akt pathway | [76] |
| Human Lipoaspirates (mean age of the patients: 20 ± 1.5 years). | HDF | Stimulation of cell proliferation and migration. Increased type-1 and type-3 collagen levels. Upregulated PI3K/Akt signaling pathway. | [78] |
| Human subcutaneous adipose tissue. | HUVEC | Increase of cell proliferation, migration and angiogenesis. Upregulation of proliferative markers and growth factors. Upregulation of Akt and ERK signaling pathways. | [79] |
| HaCaT | Increase of cells migration and proliferation. Upregulation of proliferative markers and growth factors. Upregulation of Akt and ERK signaling pathways. | ||
| Primary human foreskin fibroblasts | Increase of cells migration and proliferation | ||
| Upregulation of proliferative markers and growth factors. | |||
| Upregulation of Akt and ERK signaling pathways. | |||
| Human lipoaspirates. | HDF | Stimulated cell proliferation and migration. Increased expression of genes related with skin regeneration. | [80] |
| Adipose tissue harvested from healthy people or normal rats. | Endothelial progenitor cells isolated from peripheral blood of healthy subjects or patients with diabetes mellitus. | Increased cell viability and tube formation ability. | [81] |
| Human facial adipose tissue. | HaCaT | Increased cell proliferation and migration. Reduced H2O2-induced apoptosis levels. | [83] |
| ASC Source | Model | Effect | Reference |
|---|---|---|---|
| Human subcutaneous adipose tissue obtained from healthy females (18–35 years old). | Mouse full-thickness skin wound model | Acceleration of wound repair. | [74] |
| Human subcutaneous adipose tissue obtained after liposuction surgery (mean age of the patients: 20 ± 1.5 years). | Mouse full-thickness skin wound model | Increased number of blood vessels. Acceleration of wound repair. | [78] |
| Human subcutaneous adipose tissue obtained by liposuction. | Rat excisional wound-splinting model. | Acceleration of wound repair. | [76] |
| Human subcutaneous adipose tissue. | Mouse full-thickness skin wound model | Acceleration of wound repair. Increased collagen deposition. Increased neovascularization and cell proliferation. | [79] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombardi, F.; Palumbo, P.; Augello, F.R.; Cifone, M.G.; Cinque, B.; Giuliani, M. Secretome of Adipose Tissue-Derived Stem Cells (ASCs) as a Novel Trend in Chronic Non-Healing Wounds: An Overview of Experimental In Vitro and In Vivo Studies and Methodological Variables. Int. J. Mol. Sci. 2019, 20, 3721. https://doi.org/10.3390/ijms20153721
Lombardi F, Palumbo P, Augello FR, Cifone MG, Cinque B, Giuliani M. Secretome of Adipose Tissue-Derived Stem Cells (ASCs) as a Novel Trend in Chronic Non-Healing Wounds: An Overview of Experimental In Vitro and In Vivo Studies and Methodological Variables. International Journal of Molecular Sciences. 2019; 20(15):3721. https://doi.org/10.3390/ijms20153721
Chicago/Turabian StyleLombardi, Francesca, Paola Palumbo, Francesca Rosaria Augello, Maria Grazia Cifone, Benedetta Cinque, and Maurizio Giuliani. 2019. "Secretome of Adipose Tissue-Derived Stem Cells (ASCs) as a Novel Trend in Chronic Non-Healing Wounds: An Overview of Experimental In Vitro and In Vivo Studies and Methodological Variables" International Journal of Molecular Sciences 20, no. 15: 3721. https://doi.org/10.3390/ijms20153721
APA StyleLombardi, F., Palumbo, P., Augello, F. R., Cifone, M. G., Cinque, B., & Giuliani, M. (2019). Secretome of Adipose Tissue-Derived Stem Cells (ASCs) as a Novel Trend in Chronic Non-Healing Wounds: An Overview of Experimental In Vitro and In Vivo Studies and Methodological Variables. International Journal of Molecular Sciences, 20(15), 3721. https://doi.org/10.3390/ijms20153721