Development of Plastid Genomic Resources for Discrimination and Classification of Epimedium wushanense (Berberidaceae)

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
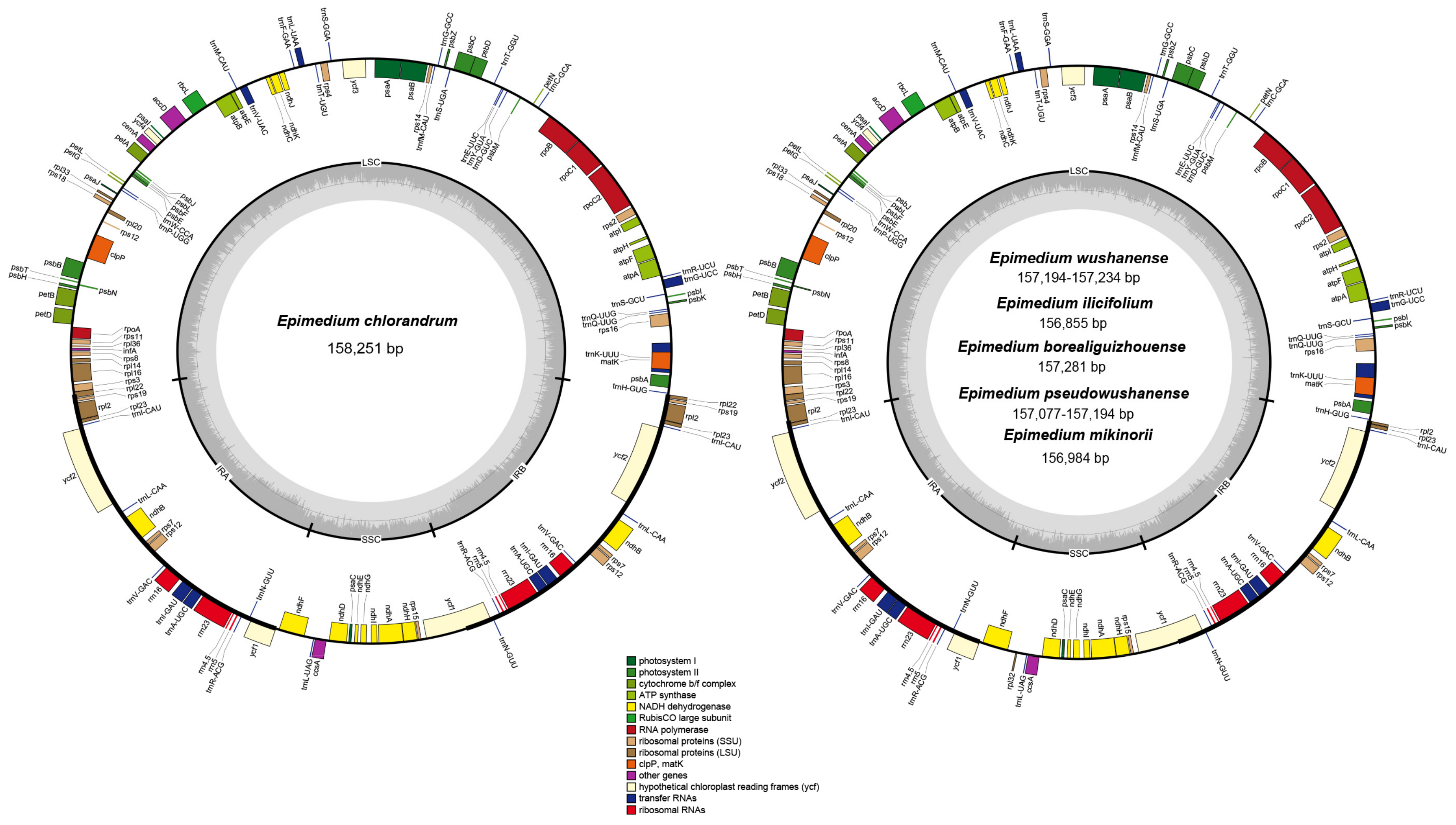
2.1. Plastid Genome Structures of E. wushanense and Its Closely Related Species
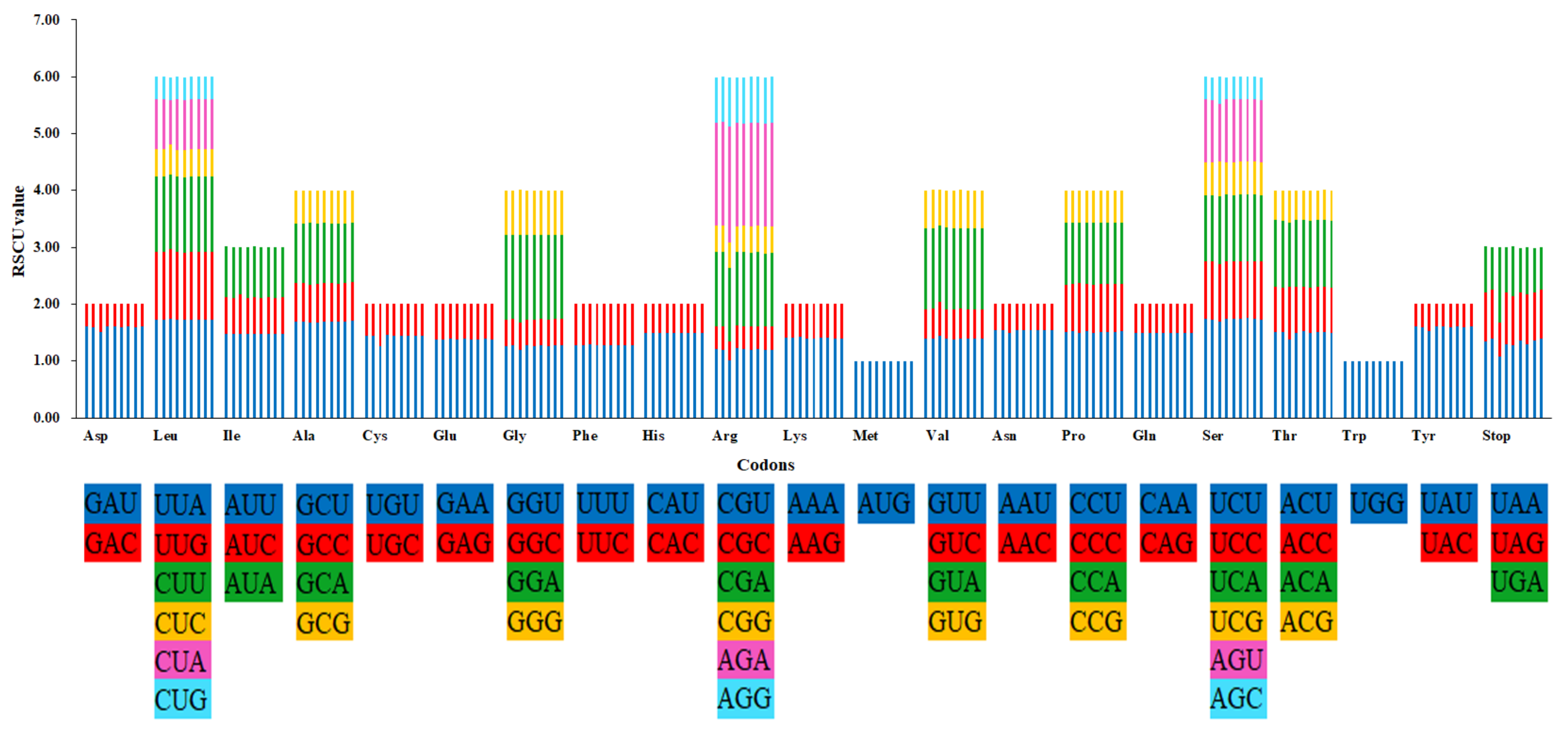
2.2. Codon Usage
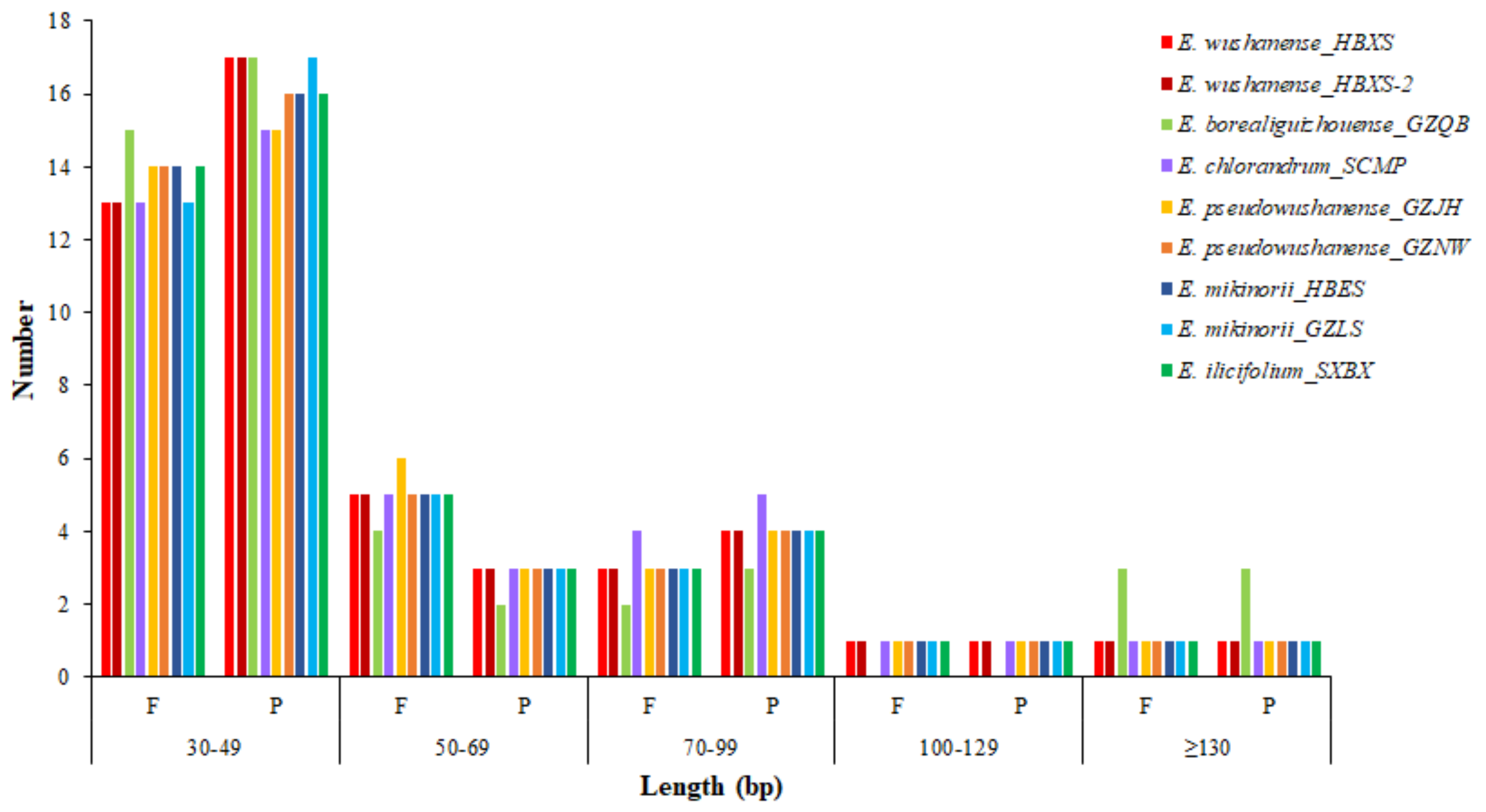
2.3. SSRs and Repeat Structure Analyses
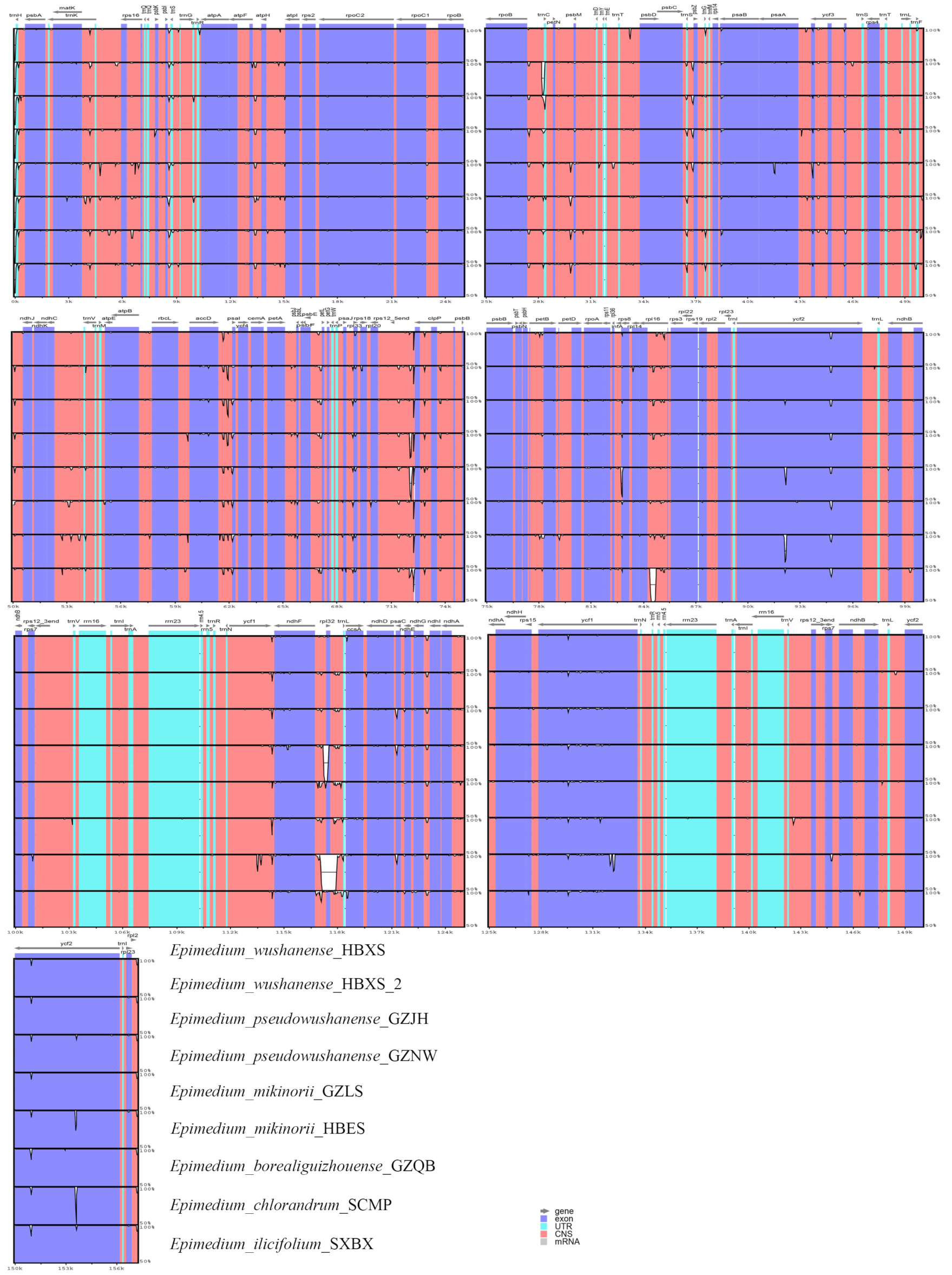
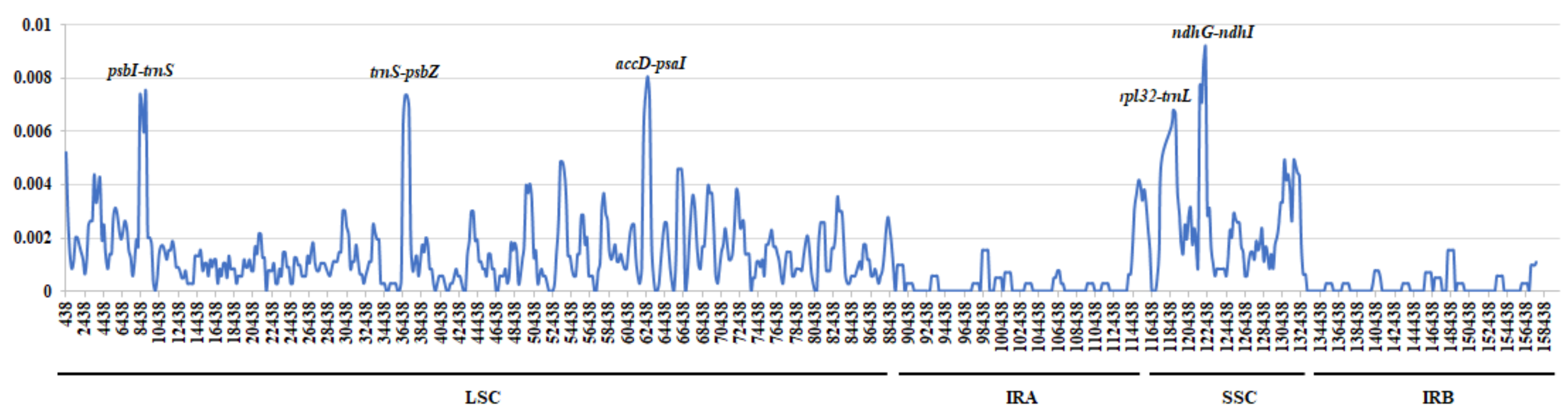
2.4. Genome Sequence Divergence
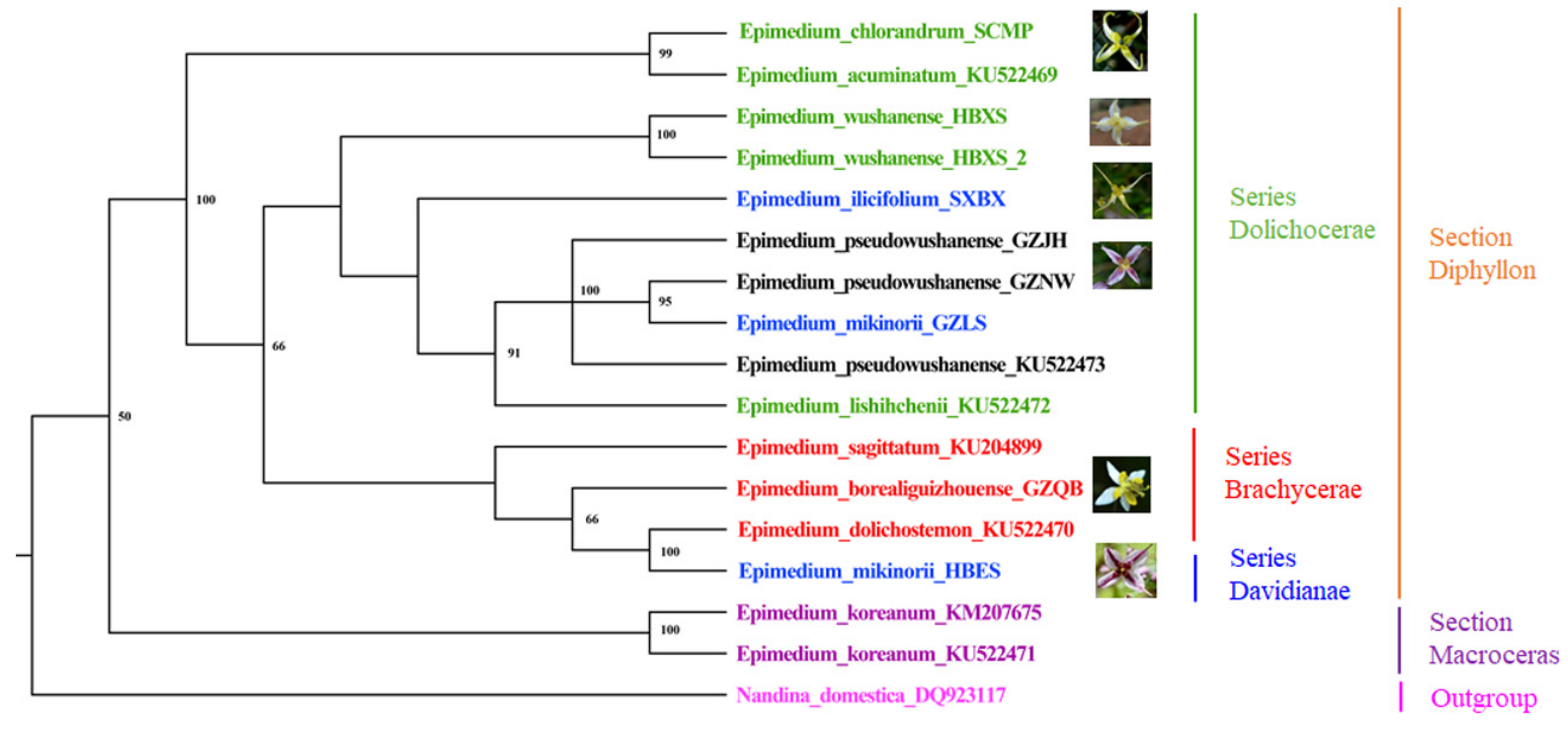
2.5. Phylogenetic and Taxonomic Implications for the Genus Epimedium
3. Materials and Methods
3.1. Taxon Sampling, DNA Extraction, and Sequencing
3.2. Genome Assembly and Genome Annotation
3.3. Genome Structure Analyses and Genome Comparison
3.4. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stearn, W.T. The Genus Epimedium and Other Herbaceous Berberidaceae; Timber Press: Portland, OR, USA, 2002. [Google Scholar]
- Zhang, M.L.; Uhink, C.H.; Kadereit, J.W. Phylogeny and biogeography of Epimedium/Vancouveria (Berberidaceae): Western North American-East Asian disjunctions, the origin of European mountain plant taxa, and East Asian species diversity. Syst. Bot. 2007, 32, 81–92. [Google Scholar] [CrossRef]
- Guo, B.L.; Pei, L.K.; Xiao, P.G. Further research on taxonomic significance of flavonoids in Epimedium (Berberidaceae). J. Syst. Evol. 2008, 46, 874–885. [Google Scholar]
- De Smet, Y.; Goetghebeur, P.; Wanke, S.; Asselman, P.; Samain, M.S. Additional evidence for recent divergence of Chinese Epimedium (Berberidaceae) derived from AFLP, chloroplast and nuclear data supplemented with characterisation of leaflet pubescence. Plant Ecol. Evol. 2012, 145, 73–87. [Google Scholar] [CrossRef]
- Ying, T.S.; Boufford, D.E.; Brach, A.R. Epimedium Linnaeus. In Flora of China; Science Press: St. Louis, MO, USA; Botanical Garden Press: Beijing, China, 2011; Volume 19, pp. 787–799. [Google Scholar]
- Guo, B.L.; He, S.Z.; Zhong, G.Y.; Xiao, P.G. Two new species of Epimedium (Berberidaceae) from China. Acta Phytotaxon. Sin. 2007, 45, 813–821. [Google Scholar] [CrossRef]
- He, S.Z. The Genus Epimedium of China in Colour; Guizhou Publishing Group: Guizhou, China, 2014. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; Chemical Industry Press: Beijing, China, 2015; Volume I. [Google Scholar]
- Ma, H.; He, X.; Yang, Y.; Li, M.; Hao, D.; Jia, Z. The genus Epimedium: An ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2011, 134, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dang, H.; Li, J.; Wang, Y. The Epimedium wushanense (Berberidaceae) species complex, with one new species from Sichuan, China. Phytotaxa 2014, 172, 39. [Google Scholar] [CrossRef]
- Guo, M.Y.; Xu, Y.Q.; Ren, L.; He, S.Z.; Pang, X.H. A systematic study on DNA barcoding of medicinally important genus Epimedium L. (Berberidaceae). Genes Basel 2018, 9, 637. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Muller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Palmer, J.D.; Jansen, R.K.; Michaels, H.J.; Chase, M.W.; Manhart, J.R. Chloroplast DNA Variation and Plant Phylogeny. Ann. Mo. Bot. Gard. 1988, 75, 1180–1206. [Google Scholar] [CrossRef]
- Wu, F.H.; Chan, M.T.; Liao, D.C.; Hsu, C.T.; Lee, Y.W.; Daniell, H.; Duvall, M.R.; Lin, C.S. Complete chloroplast genome of Oncidium Gower Ramsey and evaluation of molecular markers for identification and breeding in Oncidiinae. BMC Plant Bio. 2010, 10, 68. [Google Scholar] [CrossRef]
- Carbonell-Caballero, J.; Alonso, R.; Ibanez, V.; Terol, J.; Talon, M.; Dopazo, J. A Phylogenetic Analysis of 34 Chloroplast Genomes Elucidates the Relationships between Wild and Domestic Species within the Genus Citrus. Mol. Biol. Evol. 2015, 32, 2015–2035. [Google Scholar] [CrossRef] [PubMed]
- Curci, P.L.; De Paola, D.; Sonnante, G. Development of chloroplast genomic resources for Cynara. Mol. Ecol. Resour. 2016, 16, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Pessoa-Filho, M.; Martins, A.M.; Ferreira, M.E. Molecular dating of phylogenetic divergence between Urochloa species based on complete chloroplast genomes. BMC Genom. 2017, 18, 516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Du, L.; Liu, A.; Chen, J.; Wu, L.; Hu, W.; Zhang, W.; Kim, K.; Lee, S.; Yang, T.; et al. The Complete Chloroplast Genome Sequences of Five Epimedium Species: Lights into Phylogenetic and Taxonomic Analyses. Front. Plant Sci. 2016, 7, 306. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, X.; Cui, Y.; Sun, W.; Li, Y.; Wang, Y.; Song, J.; Yao, H. Molecular Structure and Phylogenetic Analyses of Complete Chloroplast Genomes of Two Aristolochia Medicinal Species. Int. J. Mol. Sci. 2017, 18, 1839. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, J.; Cui, Y.; Wang, Y.; Duan, B.; Yao, H. Identification of Ligularia Herbs Using the Complete Chloroplast Genome as a Super-Barcode. Front. Pharmacol. 2018, 9, 695. [Google Scholar] [CrossRef]
- Cusack, B.P.; Wolfe, K.H. When gene marriages don’t work out: Divorce by subfunctionalization. Trends Genet. 2007, 23, 270–272. [Google Scholar] [CrossRef]
- Ueda, M.; Fujimoto, M.; Arimura, S.; Murata, J.; Tsutsumi, N.; Kadowaki, K. Loss of the rpl32 gene from the chloroplast genome and subsequent acquisition of a preexisting transit peptide within the nuclear gene in Populus. Gene 2007, 402, 51–56. [Google Scholar] [CrossRef]
- Park, S.; Jansen, R.K.; Park, S. Complete plastome sequence of Thalictrum coreanum (Ranunculaceae) and transfer of the rpl32 gene to the nucleus in the ancestor of the subfamily Thalictroideae. BMC Plant Biol. 2015, 15, 40. [Google Scholar] [CrossRef]
- Zhang, Y.; Dang, H.; Li, S.; Li, J.; Wang, Y. Five new synonyms in Epimedium (Berberidaceae) from China. PhytoKeys 2015, 49, 1–12. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, H.L. Complete Chloroplast Genome Sequences from Korean Ginseng (Panax schinseng Nees) and Comparative Analysis of Sequence Evolution among 17 Vascular Plants. DNA Res. 2004, 11, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; Mcdevitt, R.; Vendramin, G.G.; Rafalski, J.A. Polymorphic simple sequence repeat regions in chloroplast genomes: Applications to the population genetics of pines. Proc. Natl. Acad. Sci. USA 1995, 92, 7759–7763. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Jia, H.; Li, X.; Chai, M.; Jia, H.; Chen, Z.; Wang, G.; Chai, C.; van de Weg, E.; Gao, Z. Development of simple sequence repeat (SSR) markers from a genome survey of Chinese bayberry (Myrica rubra). BMC Genom. 2012, 13, 201. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. Chloroplast Evolution: Secondary Symbiogenesis and Multiple Losses. Curr. Biol. 2002, 12, R62–R64. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.B.; Yang, S.X.; Li, H.T.; Yang, J.; Li, D.Z. Comparative chloroplast genomes of Camellia species. PLoS ONE 2013, 8, e73053. [Google Scholar] [CrossRef] [PubMed]
- Kane, N.; Sveinsson, S.; Dempewolf, H.; Yang, J.Y.; Zhang, D.; Engels, J.M.; Cronk, Q. Ultra-barcoding in cacao (Theobroma spp.; Malvaceae) using whole chloroplast genomes and nuclear ribosomal DNA. Am. J. Bot. 2012, 99, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Sheng, M.Y.; Wang, L.J.; Tian, X.J. Karyomorphology of eighteen species of genus Epimedium (Berberidaceae) and its phylogenetic implications. Genet. Resour. Crop Evol. 2010, 57, 1165–1176. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, L.; Chen, J.; Sun, W.; Wang, Y. Taxonomic and phylogenetic analysis of Epimedium L. based on amplified fragment length polymorphisms. Sci. Hortic. Amst. 2014, 170, 284–292. [Google Scholar] [CrossRef]
- Olmstead, R.G.; Bedoya, A.M. Whole genomes: The holy grail. A commentary on: ‘Molecular phylogenomics of the tribe Shoreeae (Dipterocarpaceae) using whole plastidgenomes’. Ann. Bot. 2019, 123, iv–v. [Google Scholar] [CrossRef]
- Lee-Yaw, J.A.; Grassa, C.J.; Joly, S.; Andrew, R.L.; Rieseberg, L.H. An evaluation of alternative explanations for widespread cytonuclear discordance in annual sunflowers (Helianthus). New Phytol. 2019, 221, 515–526. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, K.; Kim, N.R.; Lee, S.C.; Yang, T.J.; Kim, Y.D. The complete chloroplast genome of a medicinal plant Epimedium koreanum Nakai (Berberidaceae). Mitochondrial DNA A 2016, 27, 4342–4343. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Moore, M.J.; Zhang, S.; Soltis, P.S.; Soltis, D.E.; Zhao, T.; Meng, A.; Li, X.; Li, J.; Wang, H. Phylogenomic and structural analyses of 18 complete plastomes across nearly all families of early-diverging eudicots, including an angiosperm-wide analysis of IR gene content evolution. Mol. Phylogenet. Evol. 2016, 96, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Shi, L.; Zhu, Y.; Chen, H.; Zhang, J.; Lin, X.; Guan, X. CpGAVAS, an integrated web server for the annotation, visualization, analysis, and GenBank submission of completely sequenced chloroplast genome sequences. BMC Genom. 2012, 13, 715. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Helt, G.A.; Reese, J.T.; Munoz-Torres, M.C.; Childers, C.P.; Buels, R.M.; Stein, L.; Holmes, I.H.; Elsik, C.G.; Lewis, S.E. Web Apollo: A web-based genomic annotation editing platform. Genome Biol. 2013, 14, R93. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Sharp, P.M.; Li, W.H. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 1987, 15, 1281–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP Ver.# 4b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Voucher No. | Accession No. | LSC Length/bp | SSC Length/bp | IR Length/bp | Genome Size/bp | GC Content/% |
|---|---|---|---|---|---|---|---|
| E. wushanense | HBXS | MK408753 | 88,527 | 17,091 | 25,788 | 157,194 | 38.80 |
| E. wushanense | HBXS-2 | MK992920 | 88578 | 17,090 | 25,783 | 157,234 | 38.80 |
| E. pseudowushanense | GZJH | MK408750 | 88,449 | 17,070 | 25,779 | 157,077 | 38.78 |
| E. pseudowushanense | GZNW | MK992919 | 88,538 | 17,079 | 25,788 | 157,193 | 38.78 |
| E. mikinorii | HBES | MK408752 | 88,360 | 17,074 | 25,775 | 156,984 | 38.81 |
| E. mikinorii | GZLS | MK992918 | 88,522 | 16,833 | 25,783 | 156,921 | 38.80 |
| E. chlorandrum | SCMP | MK408754 | 86,627 | 16,238 | 27,693 | 158,251 | 38.89 |
| E. ilicifolium | SXBX | MK992921 | 88,217 | 17,038 | 25,800 | 156,855 | 38.77 |
| E. borealiguizhouense | GZQB | MK408751 | 88,603 | 17,040 | 25,819 | 157,281 | 38.78 |
| SSR Type | Repeat Unit | Amount/Ratio (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| E. borealiguizhouense | E. mikinorii | E. chlorandrum | E. pseudowushanense | E. wushanense | E. ilicifolium | |||||
| GZQB | HBES | GZLS | SCMP | GZJH | GZNW | HBXS | HBXS-2 | SXBX | ||
| Mono | A/T | 64/88.9 | 61/85.9 | 67/88.2 | 66/84.6 | 68/88.3 | 67/87.0 | 69/88.5 | 71/89.9 | 65/85.5 |
| C/G | 2/2.8 | 2/2.8 | 2/2.6 | 3/3.8 | 2/2.6 | 2/2.6 | 2/2.6 | 2/2.5 | 2/2.6 | |
| Di | AT/AT | 1/1.4 | 2/2.8 | 2/2.6 | 1/1.3 | 2/2.6 | 2/2.6 | 2/2.6 | 3/3.8 | 2/2.6 |
| Tri | AAT/ATT | 1/1.4 | 1/1.4 | 1/1.3 | 2/2.6 | 1/1.3 | 1/1.3 | 1/1.3 | 0/0 | 1/1.3 |
| AGG/CCT | 3/4.2 | 3/4.2 | 3/3.9 | 3/3.8 | 3/3.9 | 3/3.9 | 3/3.8 | 3/3.8 | 3/3.9 | |
| AAG/CTT | 1/1.4 | 1/1.4 | 0/0 | 1/1.3 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1.3 | |
| Tetra | AAAT/ATTT | 0/0 | 0/0 | 0/0 | 1/1.3 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1.3 |
| Penta | AAGAT/ATCTT | 0/0 | 1/1.4 | 1/1.3 | 1/1.3 | 1/1.3 | 1/1.3 | 1/1.3 | 0/0 | 0/0 |
| AATAT/ATATT | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1.3 | |
| Hexa | AACGAC/CGTTGT | 0/0 | 0/0 | 0/0 | 0/0 | 0/0 | 1/1.3 | 0/0 | 0/0 | 0/0 |
| Total | 72/100 | 71/100 | 76/100 | 78/100 | 77/100 | 77/100 | 78/100 | 79/100 | 76/100 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, M.; Ren, L.; Xu, Y.; Liao, B.; Song, J.; Li, Y.; Mantri, N.; Guo, B.; Chen, S.; Pang, X. Development of Plastid Genomic Resources for Discrimination and Classification of Epimedium wushanense (Berberidaceae). Int. J. Mol. Sci. 2019, 20, 4003. https://doi.org/10.3390/ijms20164003
Guo M, Ren L, Xu Y, Liao B, Song J, Li Y, Mantri N, Guo B, Chen S, Pang X. Development of Plastid Genomic Resources for Discrimination and Classification of Epimedium wushanense (Berberidaceae). International Journal of Molecular Sciences. 2019; 20(16):4003. https://doi.org/10.3390/ijms20164003
Chicago/Turabian StyleGuo, Mengyue, Li Ren, Yanqin Xu, Baosheng Liao, Jingyuan Song, Ying Li, Nitin Mantri, Baolin Guo, Shilin Chen, and Xiaohui Pang. 2019. "Development of Plastid Genomic Resources for Discrimination and Classification of Epimedium wushanense (Berberidaceae)" International Journal of Molecular Sciences 20, no. 16: 4003. https://doi.org/10.3390/ijms20164003
APA StyleGuo, M., Ren, L., Xu, Y., Liao, B., Song, J., Li, Y., Mantri, N., Guo, B., Chen, S., & Pang, X. (2019). Development of Plastid Genomic Resources for Discrimination and Classification of Epimedium wushanense (Berberidaceae). International Journal of Molecular Sciences, 20(16), 4003. https://doi.org/10.3390/ijms20164003