Formation of α-Farnesene in Tea (Camellia sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants



Abstract
:1. Introduction
2. Results
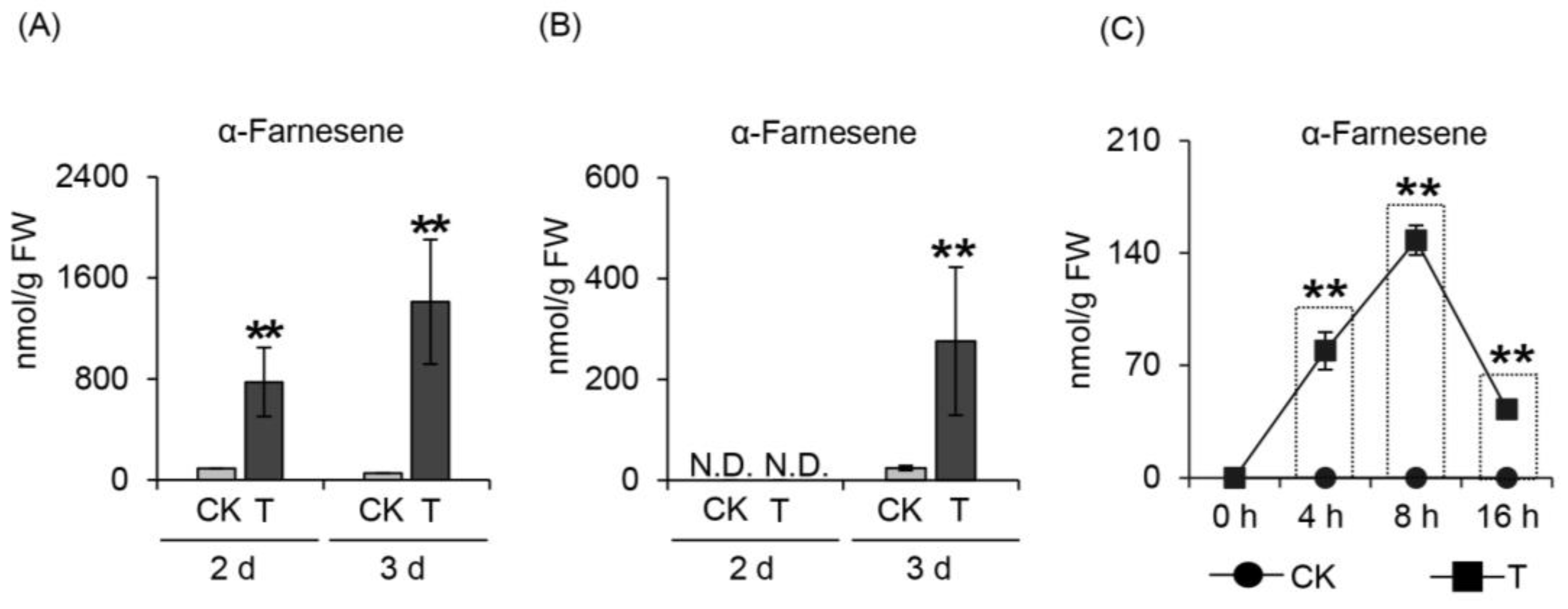
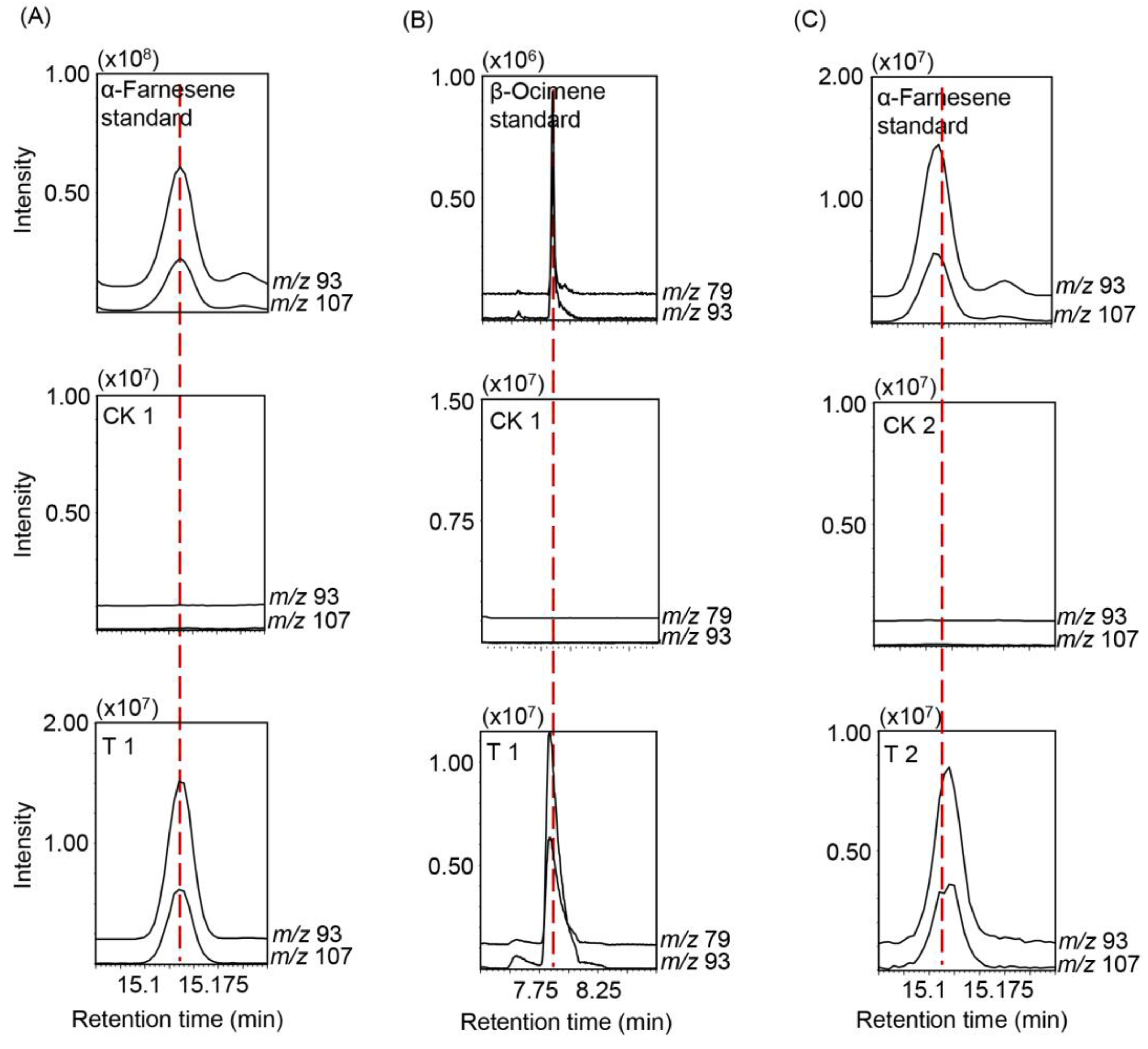
2.1. Effect of Herbivore Attack on α-Farnesene Formation
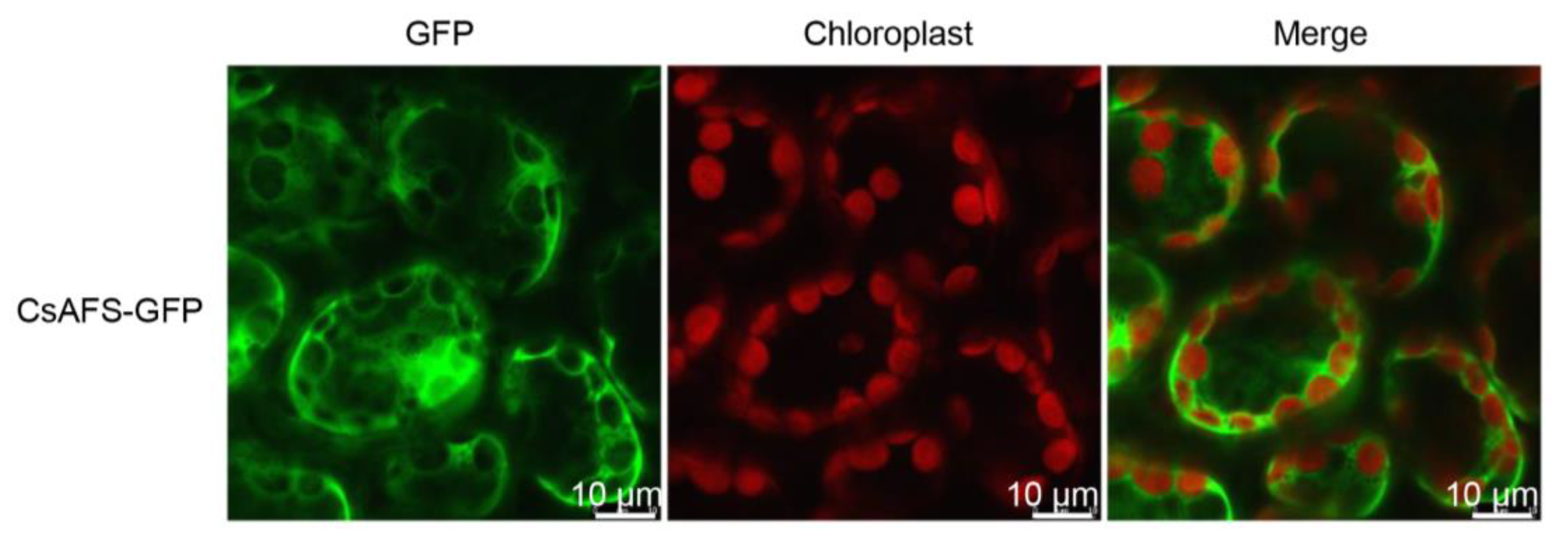
2.2. Identification of α-Farnesene Synthases in Tea Plants
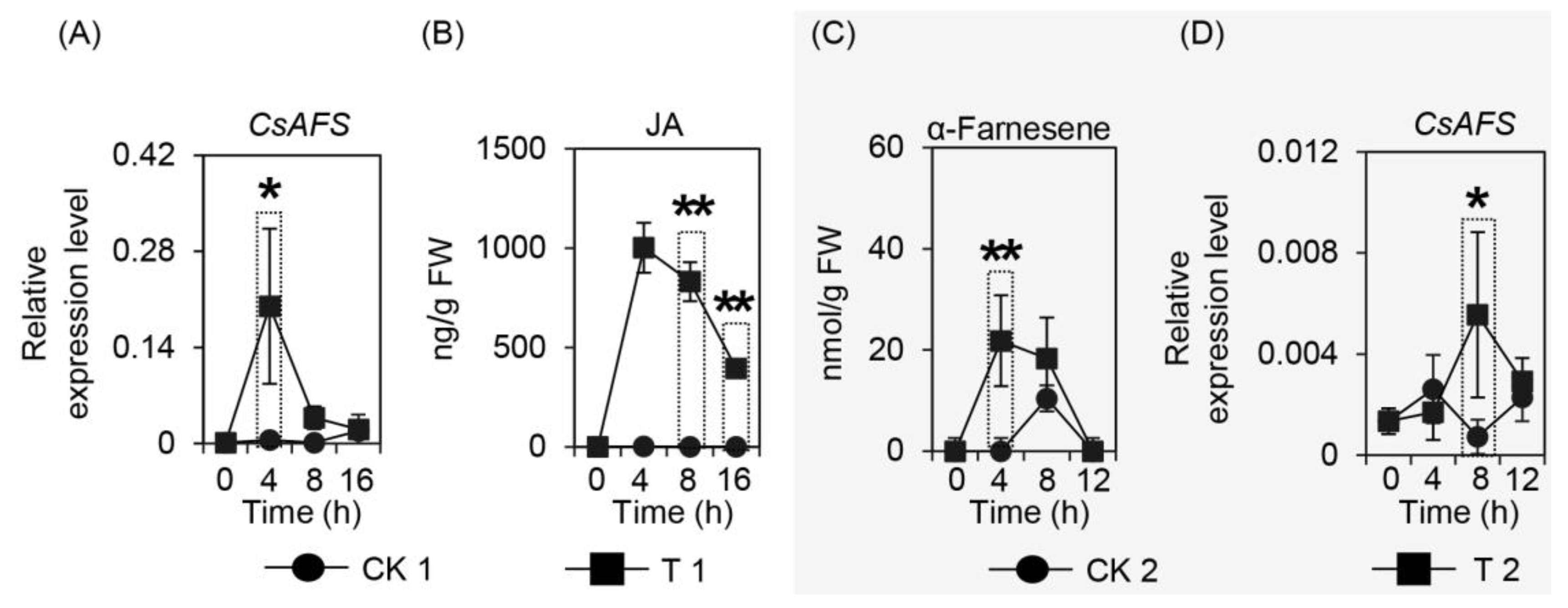
2.3. Effect of Wounding on CsAFS Expression, Jasmonic Acid Formation, and α-Farnesene Content in Tea Leaves
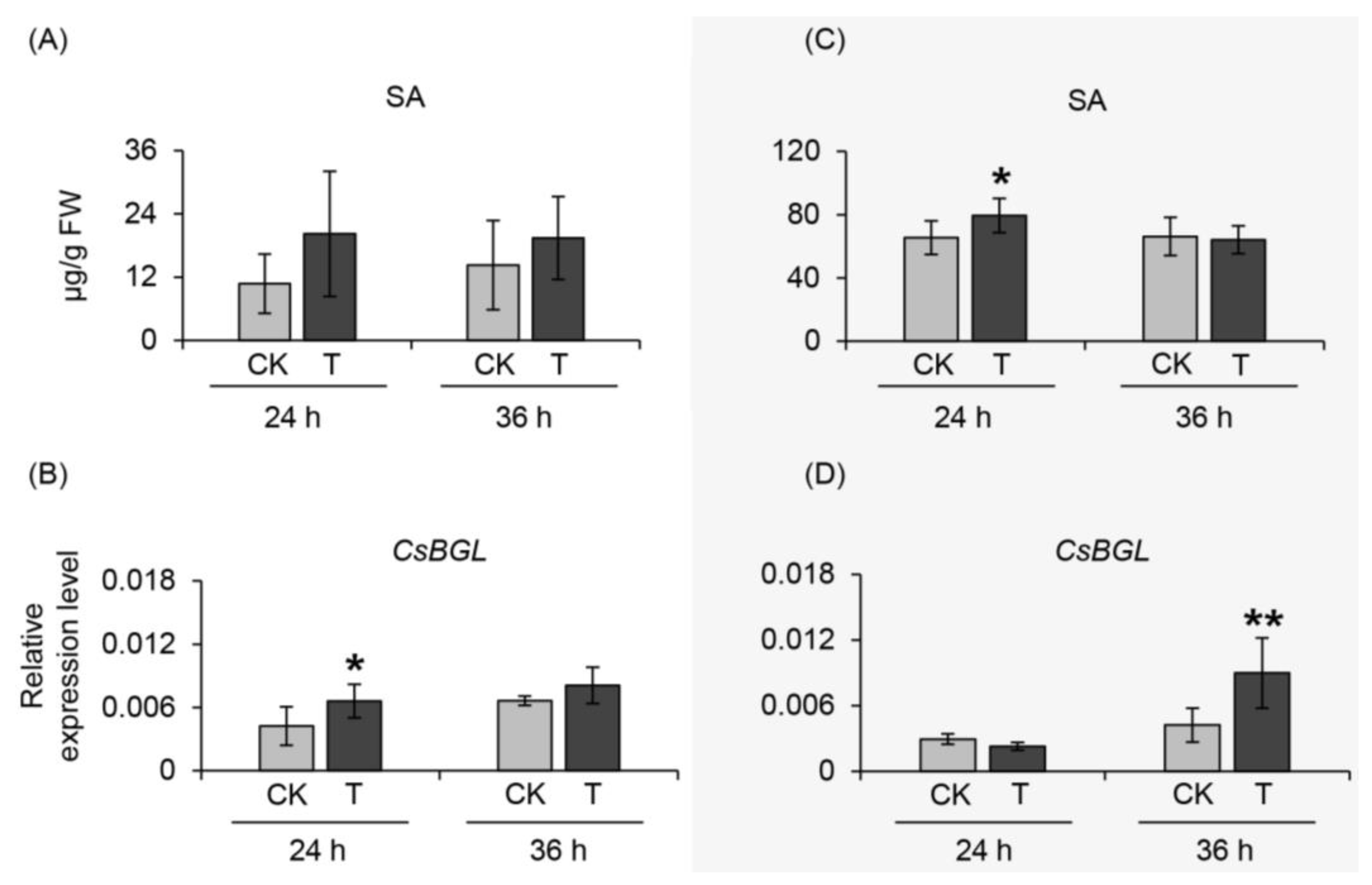
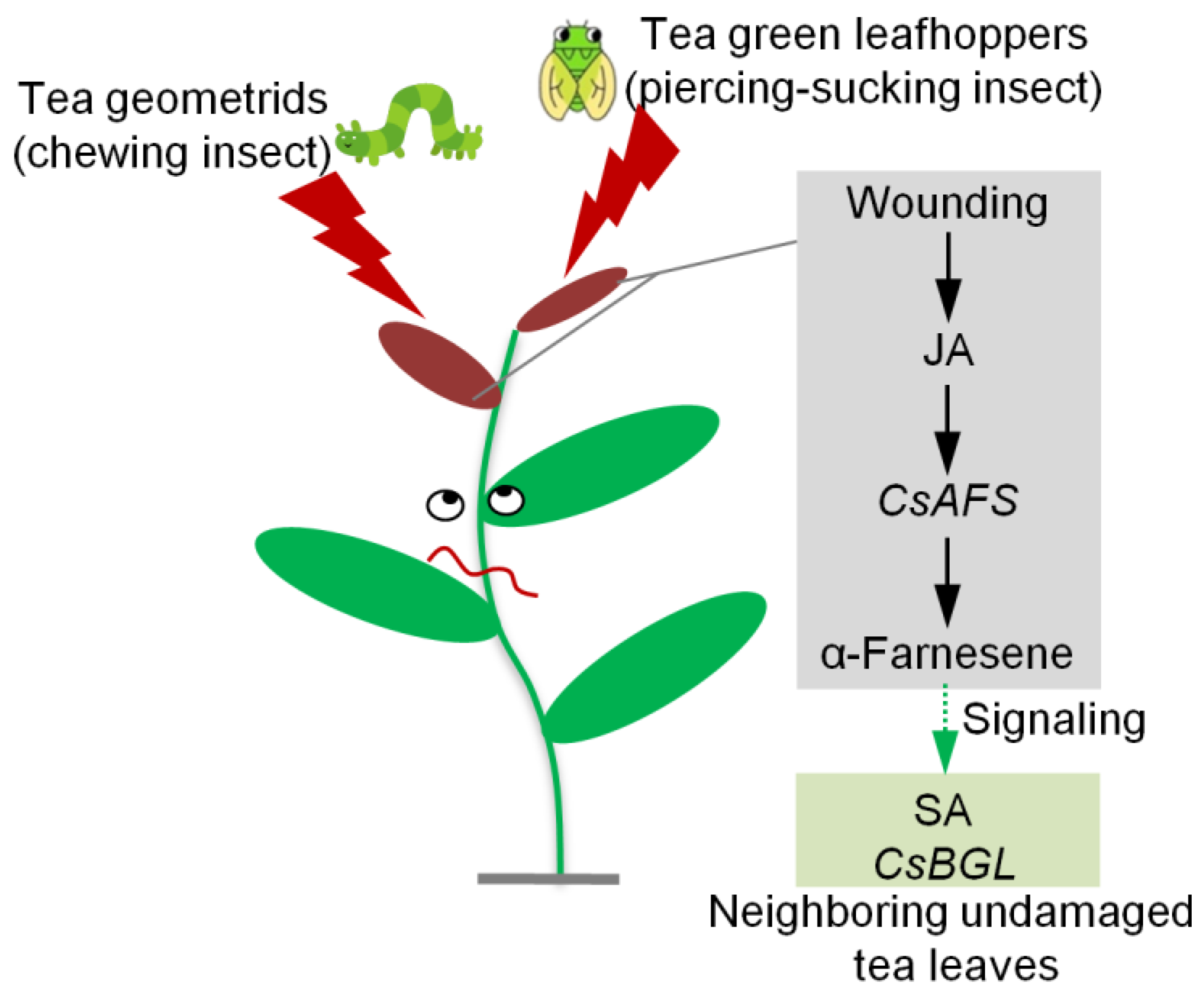
2.4. Effect of α-Farnesene on Neighboring Undamaged Tea Leaves
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.1.1. Two Species of Herbivores Attack Treatment
4.1.2. Continuous Wounding Treatment
4.1.3. JA Treatment
4.1.4. α-Farnesene Treatment
4.2. Analysis of α-Farnesene Content in Tea Leaves
4.3. Analysis of Phytohormones Content in Tea Leaves
4.4. CsAFSs Recombinant Expression and Enzyme Assay
4.4.1. RNA Isolation and Reverse Transcription
4.4.2. Gene Cloning
4.4.3. E. coli Expression of CsAFS
4.4.4. SDS-PAGE Analysis of CsAFS
4.4.5. Enzyme Assay and Functional Characterization of CsAFS
4.5. Subcellular Location Analysis of CsAFS
4.6. Analysis of CsAFS Activity in N. Benthamiana Overexpression Lines
4.7. Transcript Expression Analysis of Genes
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HIPVs | Herbivore-induced plant volatiles |
| JA | Jasmonic acid |
| SA | Salicylic acid |
| ABA | Abscisic acid |
| ET | Ethylene |
| TPS | Terpene synthase genes |
| AFS | α-Farnesene synthase |
| BGL | β-1, 3-Glucanase |
| EF1 | Encoding elongation factor 1 |
| ACS | 1-Aminocyclopropane-1-carboxylate synthase |
| EIN3 | Ethylene insensitive protein 3 |
| FPP | Farnesyl diphosphate |
| GPP | Geranyl diphosphate |
| GC-MS | Gas chromatography–mass spectrometry |
| SPME | Solid phase microextraction |
| UPLC-QTOF-MS | Ultra–performance liquid chromatography/quadrupole time–of–flight mass spectrometry |
References
- Rietveld, A.; Wiseman, S. Antioxidant effects of tea: evidence from human clinical trials. J. Nutr. 2003, 133, 3285S–3292S. [Google Scholar] [CrossRef] [PubMed]
- Alborn, H.T.; Turlings, T.C.J.; Jones, T.H.; Stenhagen, G.; Loughrin, J.H.; Tumlinson, J.H. An elicitor of plant volatiles from beet armyworm oral secretion. Science 1997, 276, 945–949. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Gershenzon, J.; Baldwin, I.T.; Kessler, A. Attracting friends to feast on foes: engineering terpene emission to make crop plants more attractive to herbivore enemies. Curr. Opin. Biotech. 2003, 14, 169–176. [Google Scholar] [CrossRef]
- Dong, F.; Yang, Z.Y.; Baldermann, S.; Sato, Y.; Asai, T.; Watanabe, N. Herbivore-induced volatiles from tea (Camellia sinensis) plants and their involvement in intraplant communication and changes in endogenous nonvolatile metabolites. J. Agr. Food Chem. 2011, 59, 13131–13135. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Liao, Y.; Li, J.; Zhou, Y.; Tang, J.; Dong, F.; Yang, Z. α-Farnesene and ocimene induce metabolite changes by volatile signaling in neighboring tea (Camellia sinensis) plants. Plant Sci. 2017, 264, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Tasin, M.; Bäckman, A.C.; Bengtsson, M.; Ioriatti, C.; Witzqall, P. Essential host plant cues in the grapevine moth. Naturwissenschaften 2006, 93, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, L.; Kännaste, A.; Niinemets, Ü. Gas chromatography-mass spectrometry method for determination of monoterpene and sesquiterpene emissions from stressed plants. Stud. U. Babes-Bol. Che. 2009, 54, 329–339. [Google Scholar]
- Kalberer, N.M.; Turlings, T.C.J.; Rahier, M. Attraction of a leaf beetle (Oreina cacaliae) to damaged host plants. J. Chem. Ecol. 2001, 27, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Hern, A.; Dorn, S. Sexual dimorphism in the olfactory orientation of adult Cydia pomonella in response to α-farnesene. Entomol. Exp. Appl. 1999, 92, 63–72. [Google Scholar] [CrossRef]
- Chen, L.; Fadamiro, H.Y. Differential electroantennogram response of females and males of two parasitoid species to host-related green leaf volatiles and inducible compounds. B. Entomol. Res. 2007, 97, 515–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.G.; Wang, C.Z. Wound-induced green leaf volatiles cause the release of acetylated derivatives and a terpenoid in maize. Phytochemistry 2006, 67, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wang, D.; Chen, X.; Köllner, T.G.; Mazarei, M.; Guo, H.; Pantalone, V.R.; Arelli, P.; Stewart, C.N., Jr.; Wang, N. An (E,E)-α-farnesene synthase gene of soybean has a role in defence against nematodes and is involved in synthesizing insect-induced volatiles. Plant Biotechnol. J. 2017, 15, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: a mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Schnee, C.; Köllner, T.G.; Gershenzon, J.; Deqenhardt, J. The maize gene terpene synthase 1 encodes a sesquiterpene synthase catalyzing the formation of (E)-β-farnesene, (E)-nerolidol, and (E,E)-farnesol after herbivore damage. Plant Physiol. 2002, 130, 2049–2060. [Google Scholar] [CrossRef] [PubMed]
- Pechous, S.W.; Whitaker, B.D. Cloning and functional expression of an (E, E)-α-farnesene synthase cDNA from peel tissue of apple fruit. Planta 2004, 219, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D.; Lee, S. Terpene specialized metabolism in Arabidopsis thaliana. Arabidopsis Book 2011. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; D’Auria, J.C.; Farooq, A.; Pichersky, E.; Gershenzon, J. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. Plant Cell 2003, 15, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Davidovich-Rikanati, R.; Lewinsohn, E.; Bar, E.; Iijima, Y.; Pichersky, E.; Sitrit, Y. Overexpression of the lemon basil α-zingiberene synthase gene increases both mono-and sesquiterpene contents in tomato fruit. Plant J. 2008, 56, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Pare, P.W.; Tumlinson, J.H. Induced synthesis of plant volatiles. Nature 1997, 385, 30–31. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Degenhardt, D.C.; Lincoln, D.E. Volatile emissions from an odorous plant in response to herbivory and methyl jasmonate exposure. J. Chem. Ecol. 2006, 32, 725–743. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, C.M.; Lewis, W.J.; Pare, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Mithöfer, A.; Wanner, G.; Boland, W. Effects of feeding Spodoptera littoralis on lima bean leaves. II. Continuous mechanical wounding resembling insect feeding is sufficient to elicit herbivory-related volatile emission. Plant Physiol. 2005, 137, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Mithöfer, A.; Boland, W. Before gene expression: early events in plant–insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.S.; Koo, A.J.K.; Gao, X.; Jayanty, S.; Thines, B.; Jones, A.D.; Howe, G.A. Regulation and function of Arabidopsis JASMONATE ZIM-domain genes in response to wounding and herbivory. Plant Physiol. 2008, 146, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Koo, A.J.K.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Hettenhausen, C.; Meldau, S.; Baldwin, I.T. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of Nicotiana attenuata. Plant Cell 2007, 19, 1096–1122. [Google Scholar] [CrossRef]
- Maeda, T.; Liu, Y.; Ishiwari, H.; Shimoda, T. Conditioned olfactory responses of a predatory mite, Neoseiulus womersleyi, to volatiles from prey-infested plants. Entomol. Exp. Appl. 2006, 121, 167–175. [Google Scholar] [CrossRef]
- Cai, X.M.; Sun, X.L.; Dong, W.X.; Wang, G.C.; Chen, Z.M. Herbivore species, infestation time, and herbivore density affect induced volatiles in tea plants. Chemoecology 2014, 24, 1–14. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [PubMed]
- Fu, X.; Chen, Y.; Mei, X.; Katsuno, T.; Kobayashi, E.; Dong, F.; Watanabe, N.; Yang, Z. Regulation of formation of volatile compounds of tea (Camellia sinensis) leaves by single light wavelength. Sci. Rep-UK 2015, 5, 16858. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; He, Y.; Yan, X.; Zhao, S.Q.; Zhu, J.Y.; Wei, C.L. Unraveling a crosstalk regulatory network of temporal aroma accumulation in tea plant (Camellia sinensis) leaves by integration of metabolomics and transcriptomics. Environ. Exp. Bot. 2018, 149, 81–94. [Google Scholar] [CrossRef]
- Deng, W.W.; Wu, Y.L.; Li, Y.Y.; Tan, Z.; Wei, C.L. Molecular cloning and characterization of hydroperoxide lyase gene in the leaves of tea plant (Camellia sinensis). J. Agr. Food Chem. 2016, 64, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.F.; Liu, J.J.; He, Z.R.; Wang, F.M.; Yang, H.; Yan, Y.F.; Gao, M.J.; Gruber, M.Y.; Wan, X.C.; Wei, S. Implementation of CsLIS/NES in linalool biosynthesis involves transcript splicing regulation in Camellia sinensis. Plant Cell Environ. 2018, 41, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Watanabe, N.; Yang, Z. Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea (Camellia sinensis) to safely and effectively improve tea aroma. Crit. Rev. Food Sci. 2019, 59, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant terpenoid synthases: molecular biology and phylogenetic analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef]
- Martin, D.M.; Fäldt, J.; Bohlmann, J. Functional characterization of nine norway spruce TPS genes and evolution of gymnosperm terpene synthases of the TPS-d subfamily. Plant Physiol. 2004, 135, 1908–1927. [Google Scholar] [CrossRef]
- Dudareva, N.; Martin, D.; Kish, C.M.; Kolosova, N.; Gorenstein, N.; Fäldt, J.; Miller, B.; Bohlmann, J. (E)-β-Ocimene and myrcene synthase genes of floral scent biosynthesis in snapdragon: function and expression of three terpene synthase genes of a new terpene synthase subfamily. Plant Cell 2003, 15, 1227–1241. [Google Scholar] [CrossRef]
- Crock, J.; Wildung, M.; Croteau, R. Isolation and bacterial expression of a sesquiterpene synthase cDNA clone from peppermint (Mentha x piperita, L.) that produces the aphid alarm pheromone (E)-β-farnesene. P. Natl. Acad. Sci. USA 1997, 94, 12833–12838. [Google Scholar] [CrossRef]
- Phillips, M.A.; Wildung, M.R.; Williams, D.C.; Hyatt, D.C.; Croteau, R. cDNA isolation, functional expression, and characterization of (+)-α-pinene synthase and (−)-α-pinene synthase from loblolly pine (Pinus taeda): stereocontrol in pinene biosynthesis. Arch. Biochem. Biophys. 2003, 411, 267–276. [Google Scholar] [CrossRef]
- Klessig, D.F.; Durner, J.; Noad, R.; Navarre, D.A.; Wendehenne, D.; Kumar, D.; Zhou, J.M.; Shah, J.; Zhang, S.; Kachroo, P.; et al. Nitric oxide and salicylic acid signaling in plant defense. P. Natl. Acad. Sci. USA 2000, 97, 8849–8855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erb, M.; Meldau, S.; Howe, G.A. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stotz, H.U.; Koch, T.; Biedermann, A.; Weniger, K.; Boland, W.; Mitchell-Olds, T. Evidence for regulation of resistance in Arabidopsis to Egyptian cotton worm by salicylic and jasmonic acid signaling pathways. Planta 2002, 214, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Senaratna, T.; Touchell, D.; Bunn, E.; Dixon, K. Acetyl salicylic acid (Aspirin) and salicylic acid induce multiple stress tolerance in bean and tomato plants. Plant Growth Regul. 2000, 30, 157–161. [Google Scholar] [CrossRef]
- Borsani, O.; Valpuesta, V.; Botella, M.A. Evidence for a role of salicylic acid in the oxidative damage generated by NaCl and osmotic stress in Arabidopsis seedlings. Plant Physiol. 2001, 126, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Malamy, J.; Carr, J.P.; Klessig, D.F.; Raskin, I. Salicylic acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990, 250, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Vernooij, B.; Friedrich, L.; Morse, A.; Reist, R.; Kolditz-Jawhar, R.; Ward, E.; Uknes, S.; Kessmann, H.; Ryals, J. Salicylic acid is not the translocated signal responsible for inducing systemic acquired resistance but is required in signal transduction. Plant Cell 1994, 6, 959–965. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile C6-aldehydes and allo-ocimene activate defense genes and induce resistance against Botrytis cinerea in Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.W.; Drew, M.C. Ethylene and plant responses to stress. Physiol. Plantarum. 1997, 100, 620–630. [Google Scholar] [CrossRef]
- Arimura, G.; Ozawa, R.; Nishioka, T.; Boland, W.; Koch, T.; Kühnemann, F.; Takabayashi, J. Herbivore-induced volatiles induce the emission of ethylene in neighboring lima bean plants. Plant J. 2002, 29, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, S1311–S1351. [Google Scholar] [CrossRef] [PubMed]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.M.; Hadwiger, L.A. Horovitz D. Molecular characterization of a pea β-1, 3-glucanase induced by Fusarium solani and chitosan challenge. Plant Mol. Bio 1992, 20, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Bucher, G.L.; Tarina, C.; Heinlein, M.; Di Serio, F.; Meins, F., Jr.; Iglesias, V.A. Local expression of enzymatically active class I β-1, 3-glucanase enhances symptoms of TMV infection in tobacco. Plant J. 2001, 28(3), 361–369. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, Y.; Gui, J.D.; Fu, X.; Mei, X.; Zhen, Y.; Ye, T.; Du, B.; Dong, F.; Watanabe, N.; et al. Formation of volatile tea constituent indole during the oolong tea manufacturing process. J. Agr. Food Chem. 2016, 64, 5011–5019. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.T.; Zhou, Y.; Fu, X.M.; Mei, X.; Cheng, S.; Gui, J.; Dong, F.; Tang, J.; Ma, S.; Yang, Z. Does oolong tea (Camellia sinensis) made from a combination of leaf and stem smell more aromatic than leaf-only tea? Contribution of the stem to oolong tea aroma. Food Chem. 2017, 237, 488–498. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, Y.; Fu, X.M.; Liao, Y.; Yuan, Y.; Jia, Y.; Dong, F.; Yang, Z. Biosynthesis of jasmine lactone in tea (Camellia sinensis) leaves and its formation in response to multiple stresses. J. Agr. Food Chem. 2018, 66, 3899–3909. [Google Scholar] [CrossRef]
- Li, J.L.; Zeng, L.T.; Liao, Y.Y.; Gu, D.; Tang, J.; Yang, Z. Influence of chloroplast defects on formation of jasmonic acid and characteristic aroma compounds in tea (Camellia sinensis) leaves exposed to postharvest stresses. Int. J. Mol. Sci. 2019, 20, 1044. [Google Scholar] [CrossRef]
- Kang, Z.W.; Liu, F.H.; Zhang, Z.F.; Tian, H.G.; Liu, T.X. Volatile β-ocimene can regulate developmental performance of peach aphid Myzus persicae through activation of defense responses in Chinese cabbage Brassica pekinensis. Front. Plant Sci. 2018, 9, 708. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Used for Cloning CsAFS-pET32a | |
| CsAFS-Forward | CCACAACTTTCCCACCTCTTCTT |
| CsAFS-Reverse | CAAATTATTTGGTGAGACCTGTGGAGG |
| CsAFS-EcoRV-Forward | AAGGCCATGGCTGATATCATGGATTGTAGTAAAGGAATGCTAG |
| CsAFS-SalI-Reverse | CCGCAAGCTTGTCGACTCAATTGATCTTGAGAGGTTCG |
| Primers Used for Cloning CsAFS-GFP | |
| CsAFS-Forward | CCACAACTTTCCCACCTCTTCTT |
| CsAFS-Reverse | CAAATTATTTGGTGAGACCTGTGGAGG |
| CsAFS-SacI-Forward | AGAACACGGGGGACGAGCTCATGGATTGTAGTAAAGGAATGCTAG |
| CsAFS-SpeI-Reverse | CCATGGTGGCACTAGTATTGATCTTGAGAGGTTCGATAAGC |
| Gene Name | Accession Number | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|---|
| CsEF1 | KA280301 | TTGGACAAGCTCAAGGCTGAACG | ATGGCCAGGAGCATCAAT GACAGT |
| CsAFS | GFMV01032657 | TGTCAACACAGCTAGAGTGGC | AGCATAGAGAGGACTTGGGC |
| CsACS | - | TGAGAGGCGATAGAGTGACATT | GCTGCTTCCAAGGCTGATT |
| CsEIN3 | - | GGTAAGGAAGGAGTTGATGCT | CCGCTTGGTATTTCGCTATTG |
| CsBGL | GGEB01035404 | GTGCCCCATTGCTCCTTAAC | TTCCACCAACTGTAGGCCAT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zeng, L.; Liao, Y.; Li, J.; Tang, J.; Yang, Z. Formation of α-Farnesene in Tea (Camellia sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants. Int. J. Mol. Sci. 2019, 20, 4151. https://doi.org/10.3390/ijms20174151
Wang X, Zeng L, Liao Y, Li J, Tang J, Yang Z. Formation of α-Farnesene in Tea (Camellia sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants. International Journal of Molecular Sciences. 2019; 20(17):4151. https://doi.org/10.3390/ijms20174151
Chicago/Turabian StyleWang, Xuewen, Lanting Zeng, Yinyin Liao, Jianlong Li, Jinchi Tang, and Ziyin Yang. 2019. "Formation of α-Farnesene in Tea (Camellia sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants" International Journal of Molecular Sciences 20, no. 17: 4151. https://doi.org/10.3390/ijms20174151
APA StyleWang, X., Zeng, L., Liao, Y., Li, J., Tang, J., & Yang, Z. (2019). Formation of α-Farnesene in Tea (Camellia sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants. International Journal of Molecular Sciences, 20(17), 4151. https://doi.org/10.3390/ijms20174151