Plant Kinesin-12: Localization Heterogeneity and Functional Implications


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Plant Kinesin-12: An Overview
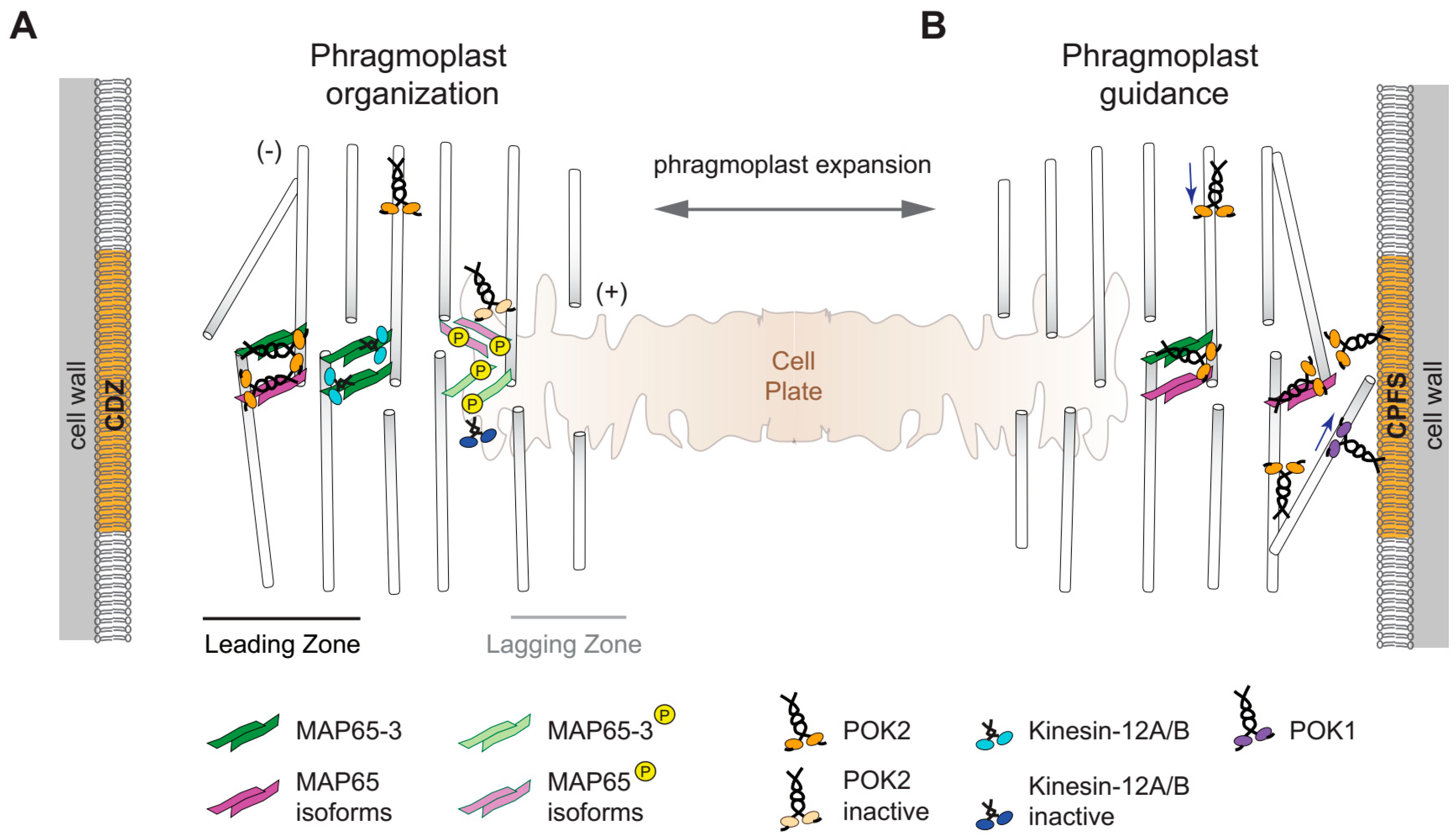
2.1. Redundancy, Conserved and Novel Functions
2.2. Kinesin-12 and Cytokinesis
2.3. Kinesin-12 POKs Are Core Components of the Cortical Division Zone
2.4. An Update on Phragmoplast Guidance: The Primary Role of POKs
3. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Cai, G.; Cresti, M. Are kinesins required for organelle trafficking in plant cells? Front. Plant Sci. 2012, 3, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.N.; Day, I.S. Microtubule Motor Proteins in the Eukaryotic Green Lineage: Functions and Regulation. In The Plant Cytoskeleton, Advances in Plant Biology 2; Liu, B., Ed.; Springer: Berlin, Germany, 2011; pp. 119–141. [Google Scholar]
- Miki, T.; Naito, H.; Nishina, M.; Goshima, G. Endogenous localizome identifies 43 mitotic kinesins in a plant cell. Proc. Natl. Acad. Sci. USA 2014, 111, E1053–E1061. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.J.; Dawe, R.K.; Christie, K.R.; Cleveland, D.W.; Dawson, S.C.; Endow, S.A.; Goldstein, L.S.B.; Goodson, H.V.; Hirokawa, N.; Howard, J.; et al. A standardized kinesin nomenclature. J. Cell Biol. 2004, 167, 19–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, H.; Okada, Y.; Hirokawa, N. Analysis of the kinesin superfamily: Insights into structure and function. Trends in Cell Biol. 2005, 15, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Lipka, E.; Gadeyne, A.; Stöckle, D.; Zimmermann, S.; De Jaeger, G.; Ehrhardt, D.W.; Kirik, V.; Van Damme, D.; Müller, S. The phragmoplast-orienting kinesin-12 class proteins translate the positional information of the preprophase band to establish the cortical division zone in Arabidopsis thaliana. Plant Cell 2014, 26, 2617–2632. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, A.; Livanos, P.; Lipka, E.; Gadeyne, A.; Hauser, M.-T.; Van Damme, D.; Müller, S. Dual localized kinesin-12 POK2 plays multiple roles during cell division and interacts with MAP65-3. EMBO Rep. 2018, 19, e46085. [Google Scholar]
- Müller, S.; Han, S.; Smith, L.G. Two kinesins are involved in the spatial control of cytokinesis in Arabidopsis thaliana. Curr. Biol. 2006, 16, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Seeger, M.A.; Rice, S.E. Intrinsic disorder in the kinesin superfamily. Biophys. Rev. 2013, 5, 233–247. [Google Scholar] [CrossRef]
- Drechsler, H.; McHugh, T.; Singleton, M.R.; Carter, N.J.; McAinsh, A.D. The Kinesin-12 Kif15 is a processive track-switching tetramer. eLife 2014, 3, e01724. [Google Scholar] [CrossRef] [Green Version]
- Chugh, M.; Reißner, M.; Bugiel, M.; Lipka, E.; Herrmann, A.; Roy, B.; Müller, S.; Schäffer, E. Phragmoplast orienting kinesin 2 is a weak motor switching between processive and diffusive modes. Biophys. J. 2018, 115, 375–385. [Google Scholar] [CrossRef]
- Shen, Z.; Collatos, A.R.; Bibeau, J.P.; Furt, F.; Vidali, L. Phylogenetic analysis of the kinesin superfamily from Physcomitrella. Front. Plant Sci. 2012, 3, 230. [Google Scholar] [CrossRef]
- Sturgill, E.G.; Das, D.K.; Takizawa, Y.; Shin, Y.; Collier, S.E.; Ohi, M.D.; Hwang, W.; Lang, M.J.; Ohi, R. Kinesin-12 Kif15 targets kinetochore fibers through an intrinsic two-step mechanism. Curr. Biol. 2014, 24, 2307–2313. [Google Scholar] [CrossRef]
- Drechsler, H.; McAinsh, A.D. Kinesin-12 motors cooperate to suppress microtubule catastrophes and drive the formation of parallel microtubule bundles. Proc. Natl. Acad. Sci. USA 2016, 113, E1635–E1644. [Google Scholar] [CrossRef] [Green Version]
- Lakshmikanth, G.S.; Warrick, H.M.; Spudich, J.A. A mitotic kinesin-like protein required for normal karyokinesis, myosin localization to the furrow, and cytokinesis in Dictyostelium. Proc. Natl. Acad. Sci. USA 2004, 101, 16519–16524. [Google Scholar] [CrossRef]
- Liu, M.; Nadar, V.C.; Kozielski, F.; Kozlowska, M.; Yu, W.; Baas, P.W. Kinesin-12, a mitotic microtubule-associated motor protein, impacts axonal growth, navigation, and branching. J. Neurosci. 2010, 30, 14896–14906. [Google Scholar] [CrossRef]
- Buster, D.W.; Baird, D.H.; Yu, W.; Solowska, J.M.; Chauvière, M.; Mazurek, A.; Kress, M.; Baas, P.W. Expression of the mitotic kinesin Kif15 in postmitotic neurons: Implications for neuronal migration and development. J. Neurosci. 2003, 32, 79–96. [Google Scholar] [CrossRef]
- Zmasek, C.M.; Eddy, S.R. ATV: Display and manipulation of annotated phylogenetic trees. Bioinformatics 2001, 17, 383–384. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 1–7. [Google Scholar] [CrossRef]
- Livanos, P.; Müller, S. Division plane establishment and cytokinesis. Annu. Rev. Plant Biol. 2019, 70, 239–267. [Google Scholar] [CrossRef]
- Müller, S. Plant cell division-defining and finding the sweet spot for cell plate insertion. Curr. Opin. Cell Biol. 2019, 60, 9–18. [Google Scholar] [CrossRef]
- Guo, L.; Ho, C.M.; Kong, Z.; Lee, Y.R.; Qian, Q.; Liu, B. Evaluating the microtubule cytoskeleton and its interacting proteins in monocots by mining the rice genome. Ann. Bot. 2008, 103, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.; Simmons, M.; Reddy, A. Comprehensive comparative analysis of kinesins in photosynthetic Eukaryotes. BMC Genom. 2006, 7, 18. [Google Scholar] [CrossRef]
- Tomei, E.J.; Wolniak, S.M. Transcriptome analysis reveals a diverse family of kinesins essential for spermatogenesis in the fern Marsilea. Cytoskeleton 2016, 73, 145–159. [Google Scholar] [CrossRef]
- Yamada, M.; Goshima, G. Mitotic spindle assembly in land plants: Molecules and mechanisms. Biology 2017, 6, 6. [Google Scholar] [CrossRef]
- Lee, Y.R.; Liu, B. Identification of a phragmoplast-associated kinesin-related protein in higher plants. Curr. Biol. 2000, 10, 797–800. [Google Scholar] [CrossRef] [Green Version]
- Pan, R.; Lee, Y.R.; Liu, B. Localization of two homologous Arabidopsis kinesin-related proteins in the phragmoplast. Planta 2004, 220, 156–164. [Google Scholar] [CrossRef]
- Lee, Y.R.; Li, Y.; Liu, B. Two Arabidopsis phragmoplast-associated kinesins play a critical role in cytokinesis during male gametogenesis. Plant Cell 2007, 19, 2595–2605. [Google Scholar] [CrossRef]
- Vanstraelen, M.; Inzé, D.; Geelen, D. Mitosis-specific kinesins in Arabidopsis. Trends Plant Sci. 2006, 11, 167–175. [Google Scholar] [CrossRef]
- Tian, S.; Wu, J.; Li, F.; Zou, J.; Liu, Y.; Zhou, B.; Bai, Y.; Sun, M.X. NtKRP, a kinesin-12 protein, regulates embryo/seed size and seed germination via involving in cell cycle progression at the G2/M transition. Sci. Rep. 2016, 6, 35641. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Wu, J.; Liu, Y.; Huang, X.; Li, F.; Wang, Z.; Sun, M.X. Ribosomal protein NtRPL17 interacts with kinesin-12 family protein NtKRP and functions in the regulation of embryo/seed size and radicle growth. J. Exp. Bot. 2017, 68, 5553–5564. [Google Scholar] [CrossRef] [Green Version]
- Smertenko, A.; Assaad, F.; Baluška, F.; Bezanilla, M.; Buschmann, H.; Drakakaki, G.; Hauser, M.T.; Janson, M.; Mineyuki, Y.; Moore, I.; et al. Plant Cytokinesis: Terminology for structures and processes. Trends Cell Biol. 2017, 27, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, J.; Neumann, E.; Van Damme, D.; Stoppin-Mellet, V.; Ebel, C.; Barbier, E.; Geelen, D.; Vantard, M. Two microtubule-associated proteins of Arabidopsis MAP65s promote antiparallel microtubule bundling. Mol. Biol. Cell 2008, 19, 4534–4544. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.M.; Lee, Y.R.; Kiyama, L.D.; Dinesh-Kumar, S.P.; Liu, B. Arabidopsis microtubule-associated protein MAP65-3 cross-links antiparallel microtubules toward their plus ends in the phragmoplast via its distinct C-terminal microtubule binding domain. Plant Cell 2012, 24, 2071–2085. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Smertenko, A.; Wagner, V.; Heinrich, M.; Hussey, P.J.; Hauser, M.T. The plant microtubule-associated protein AtMAP65-3/PLE is essential for cytokinetic phragmoplast function. Curr. Biol. 2004, 14, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.M.; Hotta, T.; Guo, F.; Roberson, R.W.; Lee, Y.R.; Liu, B. Interaction of antiparallel microtubules in the phragmoplast is mediated by the microtubule-associated protein MAP65-3 in Arabidopsis. Plant Cell 2011, 23, 2909–2923. [Google Scholar] [CrossRef] [PubMed]
- Kosetsu, K.; de Keijzer, J.; Janson, M.E.; Goshima, G. MICROTUBULE-ASSOCIATED PROTEIN65 is essential for maintenance of phragmoplast bipolarity and formation of the cell plate in Physcomitrella patens. Plant Cell 2013, 25, 4479–4492. [Google Scholar] [CrossRef]
- Hancock, W.O. Mitotic kinesins: A reason to delve into Kinesin-12. Curr. Biol. 2014, 24, R968–R970. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Jürgens, G. Plant cytokinesis—No ring, no constriction but centrifugal construction of the partitioning membrane. Semin. Cell Dev. Biol. 2016, 53, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Jurgens, G. Cytokinesis in higher plants. Annu. Rev. Plant Biol. 2005, 56, 281–299. [Google Scholar] [CrossRef]
- Sasabe, M.; Machida, Y. Regulation of organization and function of microtubules by the mitogen-activated protein kinase cascade during plant cytokinesis. Cytoskeleton 2012, 69, 913–918. [Google Scholar] [CrossRef]
- Ganguly, A.; DeMott, L.; Zhu, C.; McClosky, D.D.; Anderson, C.T.; Dixit, R. Importin-β directly regulates the motor activity and turnover of a kinesin-4. Dev. Cell 2018, 44, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Vavrdová, T.; Šamaj, J.; Komis, G. Phosphorylation of plant microtubule-associated proteins during cell division. Front. Plant Sci. 2019, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Petrovská, B.; Cenklová, V.; Pochylová, Z.; Kourová, H.; Doskočilová, A.; Plíhal, O.; Binarová, L.; Binarová, P. Plant Aurora kinases play a role in maintenance of primary meristems and control of endoreduplication. New Phytol. 2012, 193, 590–604. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, D.; Bouget, F.Y.; Van Poucke, K.; Inzé, D.; Geelen, D. Molecular dissection of plant cytokinesis and phragmoplast structure: A survey of GFP-tagged proteins. Plant J. 2004, 40, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Allen, T.; Kim, G.J.; Sidorova, A.; Borg, M.; Park, S.K.; Twell, D. Arabidopsis Fused kinase and the Kinesin-12 subfamily constitute a signalling module required for phragmoplast expansion. Plant J. 2012, 72, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Johnson, A.; Smertenko, A.; Rahman, D.; Park, S.K.; Hussey, P.J.; Twell, D. A divergent cellular role for the FUSED kinase family in the plant-specific cytokinetic phragmoplast. Curr. Biol. 2005, 15, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Bourdon, V.; Dickinson, H.G.; Twell, D.; Park, S.K. Arabidopsis Fused kinase TWO-IN-ONE dominantly inhibits male meiotic cytokinesis. Plant Reprod. 2014, 27, 7–17. [Google Scholar] [CrossRef]
- Rasmussen, C.G.; Sun, B.; Smith, L.G. Tangled localization at the cortical division site of plant cells occurs by several mechanisms. J. Cell Sci. 2011, 124, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Zhao, Q.; Rodrigo-Peiris, T.; Brkljacic, J.; He, C.S.; Müller, S.; Meier, I. RanGAP1 is a continuous marker of the Arabidopsis cell division plane. Proc. Natl. Acad. Sci. USA 2008, 105, 18637–18642. [Google Scholar] [CrossRef]
- Walker, K.L.; Müller, S.; Moss, D.; Ehrhardt, D.W.; Smith, L.G. Arabidopsis TANGLED identifies the division plane throughout mitosis and cytokinesis. Curr. Biol. 2007, 17, 1827–1836. [Google Scholar] [CrossRef]
- Schaefer, E.; Belcram, K.; Uyttewaal, M.; Duroc, Y.; Goussot, M.; Legland, D.; Laruelle, E.; de Tauzia-Moreau, M.L.; Pastuglia, M.; Bouchez, D. The preprophase band of microtubules controls the robustness of division orientation in plants. Science 2017, 356, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Stöckle, D.; Herrmann, A.; Lipka, E.; Lauster, T.; Gavidia, R.; Zimmermann, S.; Müller, S. Putative RopGAPs impact division plane selection and interact with kinesin-12 POK1. Nat. Plants 2016, 2, 16120. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, H.; Dols, J.; Kopischke, S.; Peña, E.J.; Andrade-Navarro, M.A.; Heinlein, M.; Szymanski, D.B.; Zachgo, S.; Doonan, J.H.; Lloyd, C.W. Arabidopsis KCBP interacts with AIR9 but stays in the cortical division zone throughout mitosis via its MyTH4-FERM domain. J. Cell Sci. 2015, 128, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Abu-Abied, M.; Belausov, E.; Hagay, S.; Peremyslov, V.; Dolja, V.; Sadot, E. Myosin XI-K is involved in root organogenesis, polar auxin transport, and cell division. J. Exp. Bot. 2018, 69, 2869–2881. [Google Scholar] [CrossRef] [PubMed]
- Arima, K.; Tamaoki, D.; Mineyuki, Y.; Yasuhara, H.; Nakai, T.; Shimmen, T.; Yoshihisa, T.; Sonobe, S. Displacement of the mitotic apparatuses by centrifugation reveals cortical actin organization during cytokinesis in cultured tobacco BY-2 cells. J. Plant Res. 2018, 131, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, S.; Knight, A.E.; Hodge, T.P.; Baluška, F.; Šamaj, J.; Volkmann, D.; Kendrick-Jones, J. Characterization of the unconventional myosin VIII in plant cells and its localization at the post-cytokinetic cell wall. Plant J. 1999, 19, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Molchan, T.M.; Valster, A.H.; Hepler, P.K. Actomyosin promotes cell plate alignment and late lateral expansion in Tradescantia stamen hair cells. Planta 2002, 214, 683–693. [Google Scholar] [CrossRef]
- Feng, J.; Hu, Z.; Chen, H.; Hua, J.; Wu, R.; Dong, Z.; Qiang, L.; Liu, Y.; Baas, P.W.; Liu, M. Depletion of kinesin-12, a myosin-IIB interacting protein, promotes migration of cortical astrocytes. J. Cell Sci. 2016, 129, 2438–2447. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-Z.; Bezanilla, M. Myosin VIII associates with microtubule ends and together with actin plays a role in guiding plant cell division. eLife 2014, 3, e03498. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, S.; Livanos, P. Plant Kinesin-12: Localization Heterogeneity and Functional Implications. Int. J. Mol. Sci. 2019, 20, 4213. https://doi.org/10.3390/ijms20174213
Müller S, Livanos P. Plant Kinesin-12: Localization Heterogeneity and Functional Implications. International Journal of Molecular Sciences. 2019; 20(17):4213. https://doi.org/10.3390/ijms20174213
Chicago/Turabian StyleMüller, Sabine, and Pantelis Livanos. 2019. "Plant Kinesin-12: Localization Heterogeneity and Functional Implications" International Journal of Molecular Sciences 20, no. 17: 4213. https://doi.org/10.3390/ijms20174213
APA StyleMüller, S., & Livanos, P. (2019). Plant Kinesin-12: Localization Heterogeneity and Functional Implications. International Journal of Molecular Sciences, 20(17), 4213. https://doi.org/10.3390/ijms20174213