Chinese Herbal Medicine Ganoderma tsugae Displays Potential Anti-Cancer Efficacy on Metastatic Prostate Cancer Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. GTEE Inhibits Cell Growth and Metastatic Capability of mPCa Cells
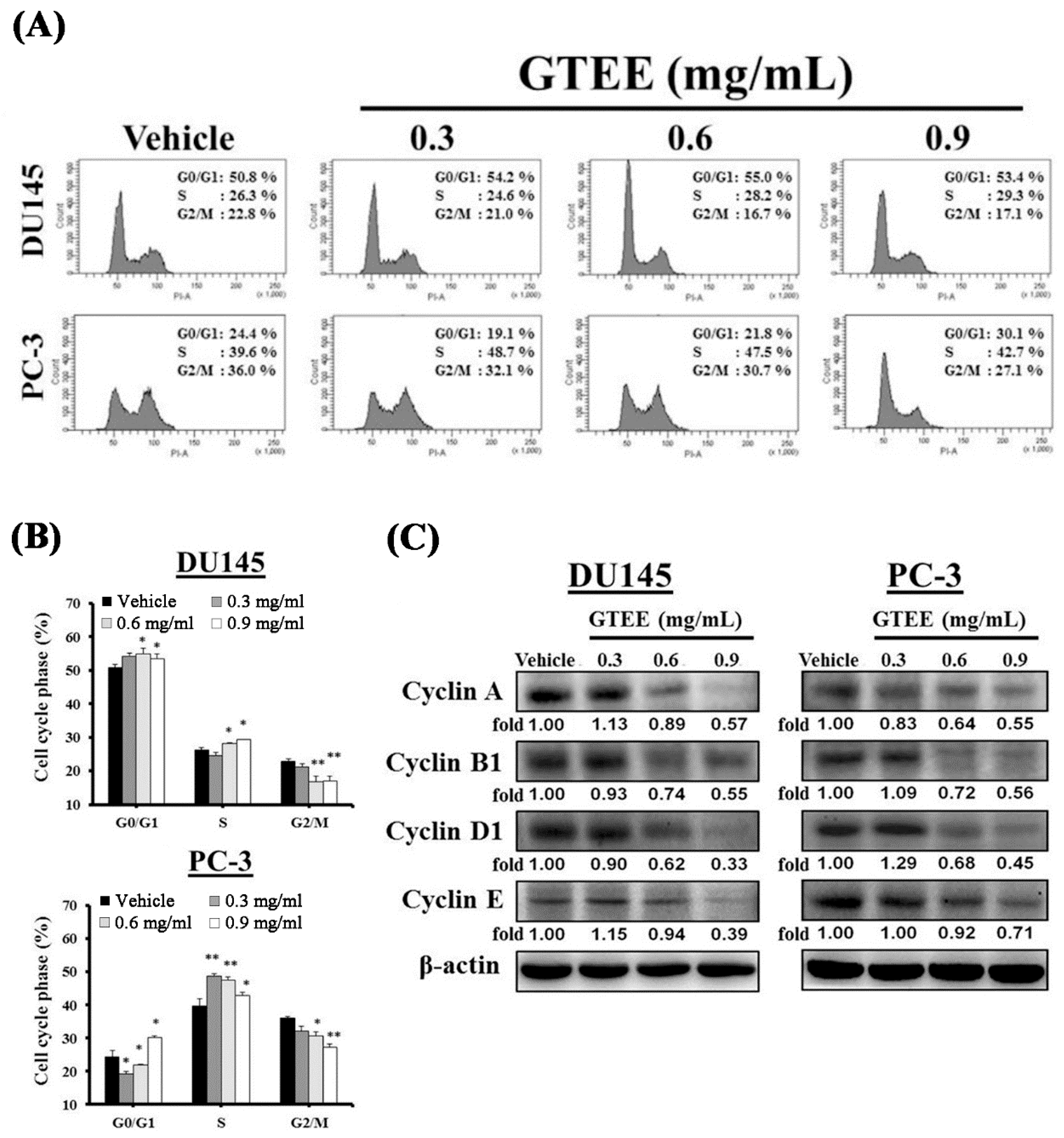
2.2. GTEE Induces Cell Cycle Arrest and Inhibits Expression of Cell Cycle-Related Proteins in mPCa Cells
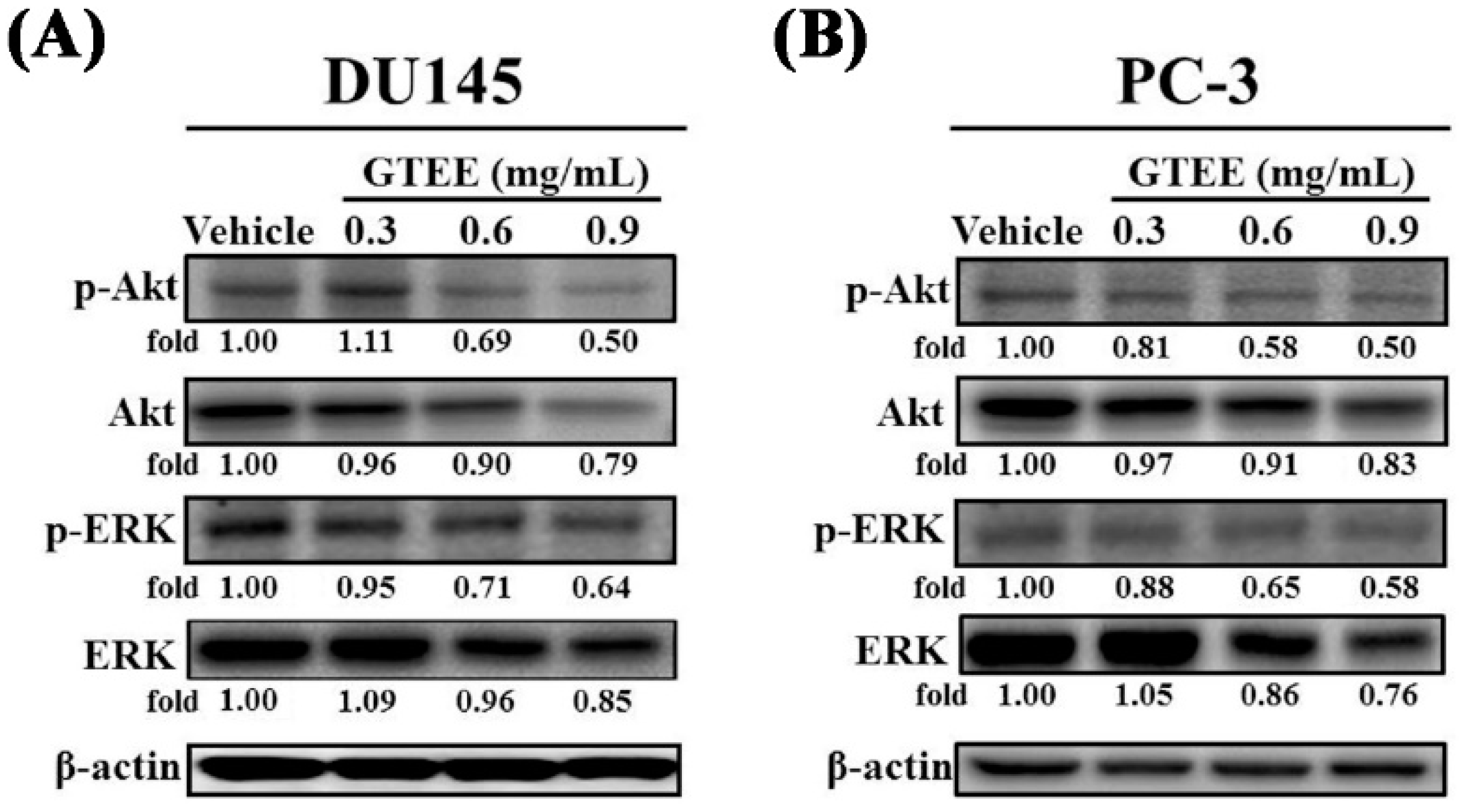
2.3. GTEE Inhibits the PI3K/Akt and MAPK/ERK Signaling Pathways Associated with Cell Growth of mPCa
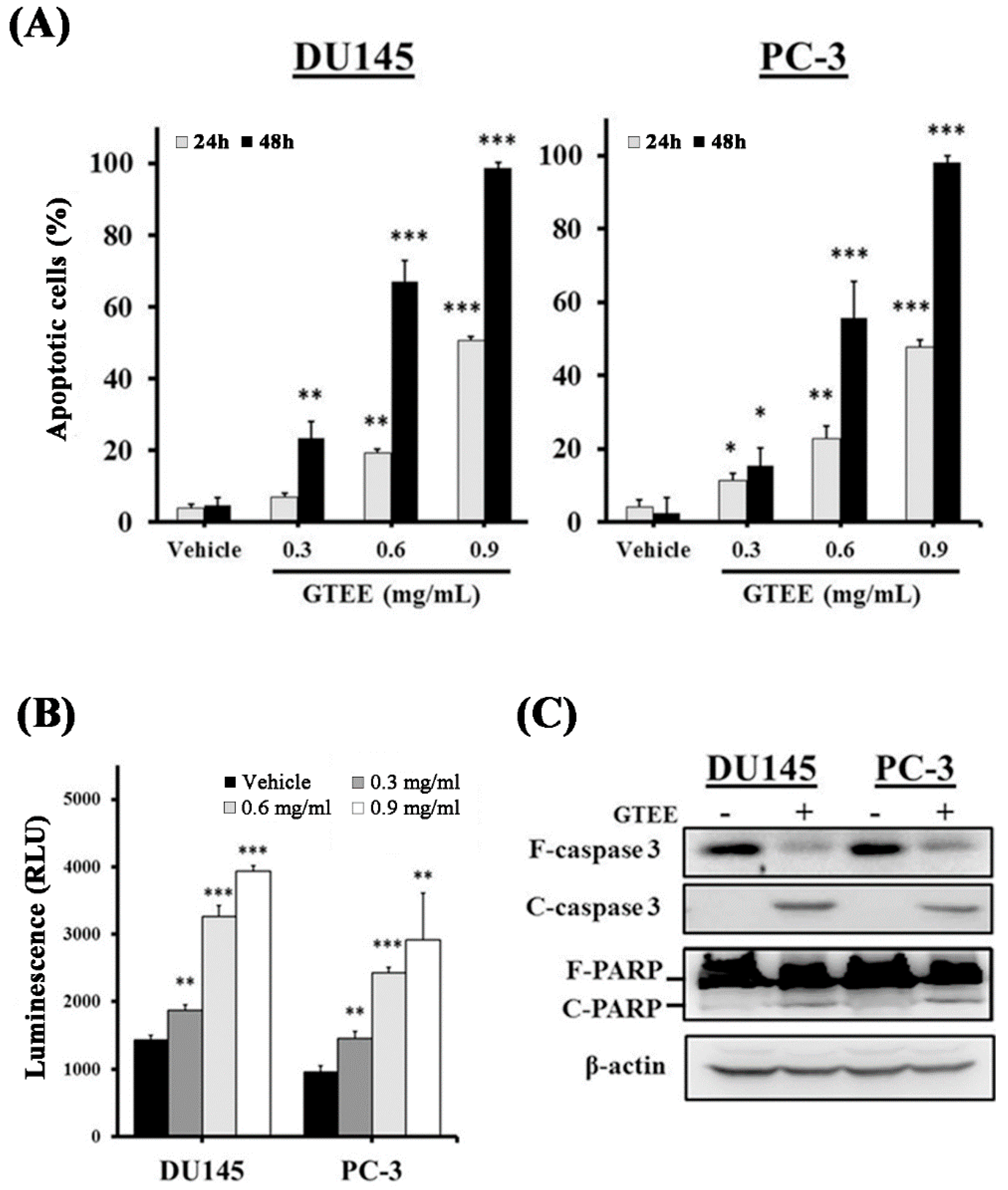
2.4. GTEE Induces Caspase-Dependent Apoptosis in mPCa Cells
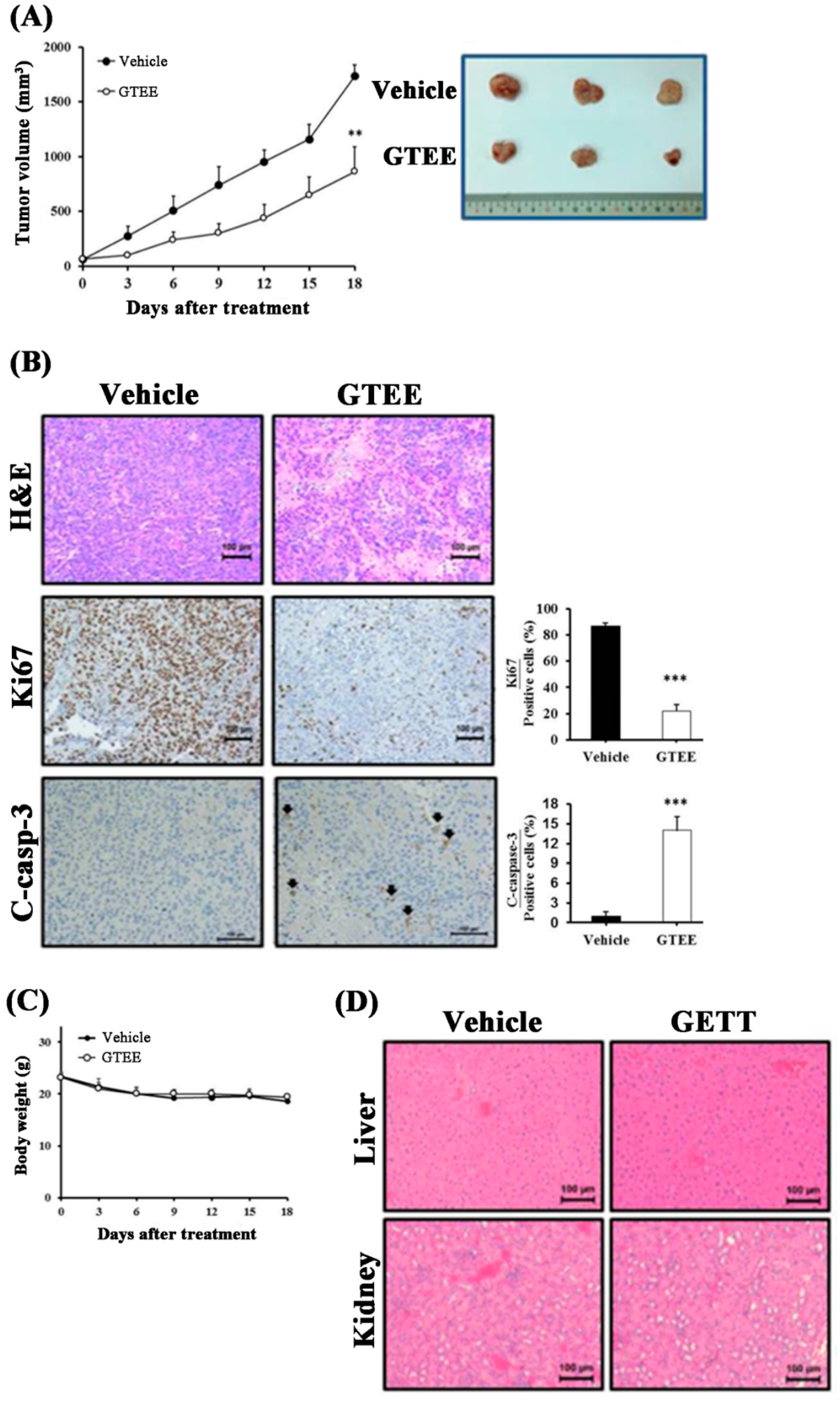
2.5. GTEE Suppresses PC-3 Tumor Growth in a Subcutaneous Xenograft Mouse Model
3. Discussion
4. Materials and Methods
4.1. Preparation of Ganoderma Tsugae Extract
4.2. Cell Lines and Cell Culture
4.3. Cell Proliferation and Progression Assays
4.4. Flow Cytometric Analysis
4.5. Western Blot Analysis
4.6. Animal Experiments
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADT | androgen deprivation therapy |
| FITC | fluorescein isothiocyanate |
| GTEE | Ganoderma tsugae ethanol extract |
| mPCa | metastatic prostate cancer |
| PCa | prostate cancer |
| TCM | traditional Chinese medicine |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.B.; Mostaghel, E.A.; Vessella, R.; Hess, D.L.; Kalhorn, T.F.; Higano, C.S.; True, L.D.; Nelson, P.S. Maintenance of intratumoral androgens in metastatic prostate cancer: A mechanism for castration-resistant tumor growth. Cancer Res. 2008, 68, 4447–4454. [Google Scholar] [CrossRef] [PubMed]
- Mohler, J.L. Castration-recurrent prostate cancer is not androgen-independent. Adv. Exp. Med. Biol. 2008, 617, 223–234. [Google Scholar] [PubMed]
- Mostaghel, E.A.; Page, S.T.; Lin, D.W.; Fazli, L.; Coleman, I.M.; True, L.D.; Knudsen, B.; Hess, D.L.; Nelson, C.C.; Matsumoto, A.M.; et al. Intraprostatic androgens and androgen-regulated gene expression persist after testosterone suppression: Therapeutic implications for castration-resistant prostate cancer. Cancer Res. 2007, 67, 5033–5041. [Google Scholar] [CrossRef] [PubMed]
- Ramudo Cela, L.; Balea Filgueiras, J.; Vizoso Hermida, J.R.; Martin Herranz, I. Study of cases of abiraterone discontinuation due to toxicity in pre-chemotherapy after 1 year’s experience. J. Oncol. Pharm. Pract. 2017, 8, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Tsao, C.K.; Cutting, E.; Martin, J.; Oh, W.K. The role of cabazitaxel in the treatment of metastatic castration-resistant prostate cancer. Ther. Adv. Urol. 2014, 6, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Boudadi, K.; Antonarakis, E.S. Resistance to Novel Antiandrogen Therapies in Metastatic Castration-Resistant Prostate Cancer. Clin. Med. Insights. Oncol. 2016, 10, 1–9. [Google Scholar] [CrossRef]
- Bishop, K.S.; Kao, C.H.; Xu, Y.; Glucina, M.P.; Paterson, R.R.; Ferguson, L.R. From 2000 years of Ganoderma lucidum to recent developments in nutraceuticals. Phytochemistry 2015, 114, 56–65. [Google Scholar] [CrossRef]
- Shiao, M.S. Natural products of the medicinal fungus Ganoderma lucidum: Occurrence, biological activities, and pharmacological functions. Chem. Rec. 2003, 3, 172–180. [Google Scholar] [CrossRef]
- Kuok, Q.Y.; Yeh, C.Y.; Su, B.C.; Hsu, P.L.; Ni, H.; Liu, M.Y.; Mo, F.E. The triterpenoids of Ganoderma tsugae prevent stress-induced myocardial injury in mice. Mol. Nutr. Food Res. 2013, 57, 1892–1896. [Google Scholar]
- Chen, M.L.; Hsieh, C.C.; Chiang, B.L.; Lin, B.F. Triterpenoids and Polysaccharide Fractions of Ganoderma tsugae Exert Different Effects on Antiallergic Activities. Evid. Based Complement Alternat. Med. 2015, 2015, 10. [Google Scholar]
- Kuo, H.P.; Hsu, S.C.; Ou, C.C.; Li, J.W.; Tseng, H.H.; Chuang, T.C.; Liu, J.Y.; Chen, S.J.; Su, M.H.; Cheng, Y.C.; et al. Ganoderma tsugae extract inhibits growth of HER2-overexpressing cancer cells via modulation of HER2/PI3K/Akt signaling pathway. Evid. Based Complement. Alternat. Med. 2013, 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Kuo, H.P.; Hsieh, H.H.; Li, J.W.; Hsu, W.H.; Chen, S.J.; Su, M.H.; Liu, S.H.; Cheng, Y.C.; Chen, C.Y.; et al. Ganoderma tsugae induces S phase arrest and apoptosis in doxorubicin-resistant lung adenocarcinoma H23/0.3 cells via modulation of the PI3K/Akt signaling pathway. Evid. Based Complement. Alternat. Med. 2012, 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.C.; Ou, C.C.; Chuang, T.C.; Li, J.W.; Lee, Y.J.; Wang, V.; Liu, J.Y.; Chen, C.S.; Lin, S.C.; Kao, M.C. Ganoderma tsugae extract inhibits expression of epidermal growth factor receptor and angiogenesis in human epidermoid carcinoma cells: In vitro and in vivo. Cancer Lett. 2009, 281, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.C.; Ou, C.C.; Li, J.W.; Chuang, T.C.; Kuo, H.P.; Liu, J.Y.; Chen, C.S.; Lin, S.C.; Su, C.H.; Kao, M.C. Ganoderma tsugae extracts inhibit colorectal cancer cell growth via G2/M cell cycle arrest. J Ethnopharmacol. 2008, 120, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Huang, G.J.; Wu, H.C.; Kao, M.C.; Huang, W.C. Ganoderma tsugae inhibits the SREBP-1/AR axis leading to suppression of cell growth and activation of apoptosis in prostate cancer cells. Molecules 2018, 23, 2539. [Google Scholar] [CrossRef] [PubMed]
- Kaighn, M.E.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L.W. Establishment and characterization of a human prostatic carcinoma cell line (PC-3). Invest Urol 1979, 17, 16–23. [Google Scholar]
- Stone, K.R.; Mickey, D.D.; Wunderli, H.; Mickey, G.H.; Paulson, D.F. Isolation of a human prostate carcinoma cell line (DU 145). Int. J. Cancer 1978, 21, 274–281. [Google Scholar] [CrossRef]
- Grana, X.; Reddy, E.P. Cell cycle control in mammalian cells: Role of cyclins, cyclin dependent kinases (CDKs), growth suppressor genes and cyclin-dependent kinase inhibitors (CKIs). Oncogene 1995, 11, 211–219. [Google Scholar]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef]
- Shukla, S.; Maclennan, G.T.; Hartman, D.J.; Fu, P.; Resnick, M.I.; Gupta, S. Activation of PI3K-Akt signaling pathway promotes prostate cancer cell invasion. Int. J. Cancer 2007, 121, 1424–1432. [Google Scholar] [CrossRef] [PubMed]
- Kreisberg, J.I.; Malik, S.N.; Prihoda, T.J.; Bedolla, R.G.; Troyer, D.A.; Kreisberg, S.; Ghosh, P.M. Phosphorylation of Akt (Ser473) is an excellent predictor of poor clinical outcome in prostate cancer. Cancer Res. 2004, 64, 5232–5236. [Google Scholar] [CrossRef] [PubMed]
- Maroni, P.D.; Koul, S.; Meacham, R.B.; Koul, H.K. Mitogen Activated Protein kinase signal transduction pathways in the prostate. Cell Commun. Signal. 2004, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Berriguete, G.; Fraile, B.; Martinez-Onsurbe, P.; Olmedilla, G.; Paniagua, R.; Royuela, M. MAP Kinases and Prostate Cancer. J. Signal. Transduct. 2012, 2012, 9. [Google Scholar] [CrossRef] [PubMed]
- Boutros, R.; Lobjois, V.; Ducommun, B. CDC25 phosphatases in cancer cells: Key players? Good targets? Nat. Rev. Cancer 2007, 7, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.; Jacks, T.; Pavletich, N.P. The cell cycle and cancer. Proc. Natl. Acad. Sci. USA 1997, 94, 2776–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- McDonald, E.R., 3rd; El-Deiry, W.S. Cell cycle control as a basis for cancer drug development (Review). Int. J. Oncol. 2000, 16, 871–886. [Google Scholar] [CrossRef]
- Buolamwini, J.K. Cell cycle molecular targets in novel anticancer drug discovery. Curr. Pharm. Des. 2000, 6, 379–392. [Google Scholar] [CrossRef]
- Elledge, S.J. Cell cycle checkpoints: Preventing an identity crisis. Science 1996, 274, 1664–1672. [Google Scholar] [CrossRef]
- Li, X.; Chen, Y.T.; Hu, P.; Huang, W.C. Fatostatin displays high antitumor activity in prostate cancer by blocking SREBP-regulated metabolic pathways and androgen receptor signaling. Mol. Cancer Ther. 2014, 13, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, J.B.; Chung, L.W.; Huang, W.C. Anti-cancer efficacy of SREBP inhibitor, alone or in combination with docetaxel, in prostate cancer harboring p53 mutations. Oncotarget 2015, 38, 41018–41032. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Janicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, J.B.; Li, Q.; Shigemura, K.; Chung, L.W.; Huang, W.C. SREBP-2 promotes stem cell-like properties and metastasis by transcriptional activation of c-Myc in prostate cancer. Oncotarget 2016, 7, 12869–12884. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, Y.T.; Josson, S.; Mukhopadhyay, N.K.; Kim, J.; Freeman, M.R.; Huang, W.C. MicroRNA-185 and 342 inhibit tumorigenicity and induce apoptosis through blockade of the SREBP metabolic pathway in prostate cancer cells. PLoS ONE 2013, 8, e70987. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.-C.; Chang, M.-S.; Huang, S.-Y.; Tsai, C.-J.; Kuo, P.-H.; Chang, H.-W.; Huang, S.-T.; Kuo, C.-L.; Lee, S.-L.; Kao, M.-C. Chinese Herbal Medicine Ganoderma tsugae Displays Potential Anti-Cancer Efficacy on Metastatic Prostate Cancer Cells. Int. J. Mol. Sci. 2019, 20, 4418. https://doi.org/10.3390/ijms20184418
Huang W-C, Chang M-S, Huang S-Y, Tsai C-J, Kuo P-H, Chang H-W, Huang S-T, Kuo C-L, Lee S-L, Kao M-C. Chinese Herbal Medicine Ganoderma tsugae Displays Potential Anti-Cancer Efficacy on Metastatic Prostate Cancer Cells. International Journal of Molecular Sciences. 2019; 20(18):4418. https://doi.org/10.3390/ijms20184418
Chicago/Turabian StyleHuang, Wen-Chin, Meng-Shiun Chang, Shih-Yin Huang, Ching-Ju Tsai, Pin-Hung Kuo, Han-Wen Chang, Sheng-Teng Huang, Chao-Lin Kuo, Shou-Lun Lee, and Ming-Ching Kao. 2019. "Chinese Herbal Medicine Ganoderma tsugae Displays Potential Anti-Cancer Efficacy on Metastatic Prostate Cancer Cells" International Journal of Molecular Sciences 20, no. 18: 4418. https://doi.org/10.3390/ijms20184418
APA StyleHuang, W. -C., Chang, M. -S., Huang, S. -Y., Tsai, C. -J., Kuo, P. -H., Chang, H. -W., Huang, S. -T., Kuo, C. -L., Lee, S. -L., & Kao, M. -C. (2019). Chinese Herbal Medicine Ganoderma tsugae Displays Potential Anti-Cancer Efficacy on Metastatic Prostate Cancer Cells. International Journal of Molecular Sciences, 20(18), 4418. https://doi.org/10.3390/ijms20184418