Monitoring of Low-Intensity Exposures via Luminescent Bioassays of Different Complexity: Cells, Enzyme Reactions, and Fluorescent Proteins



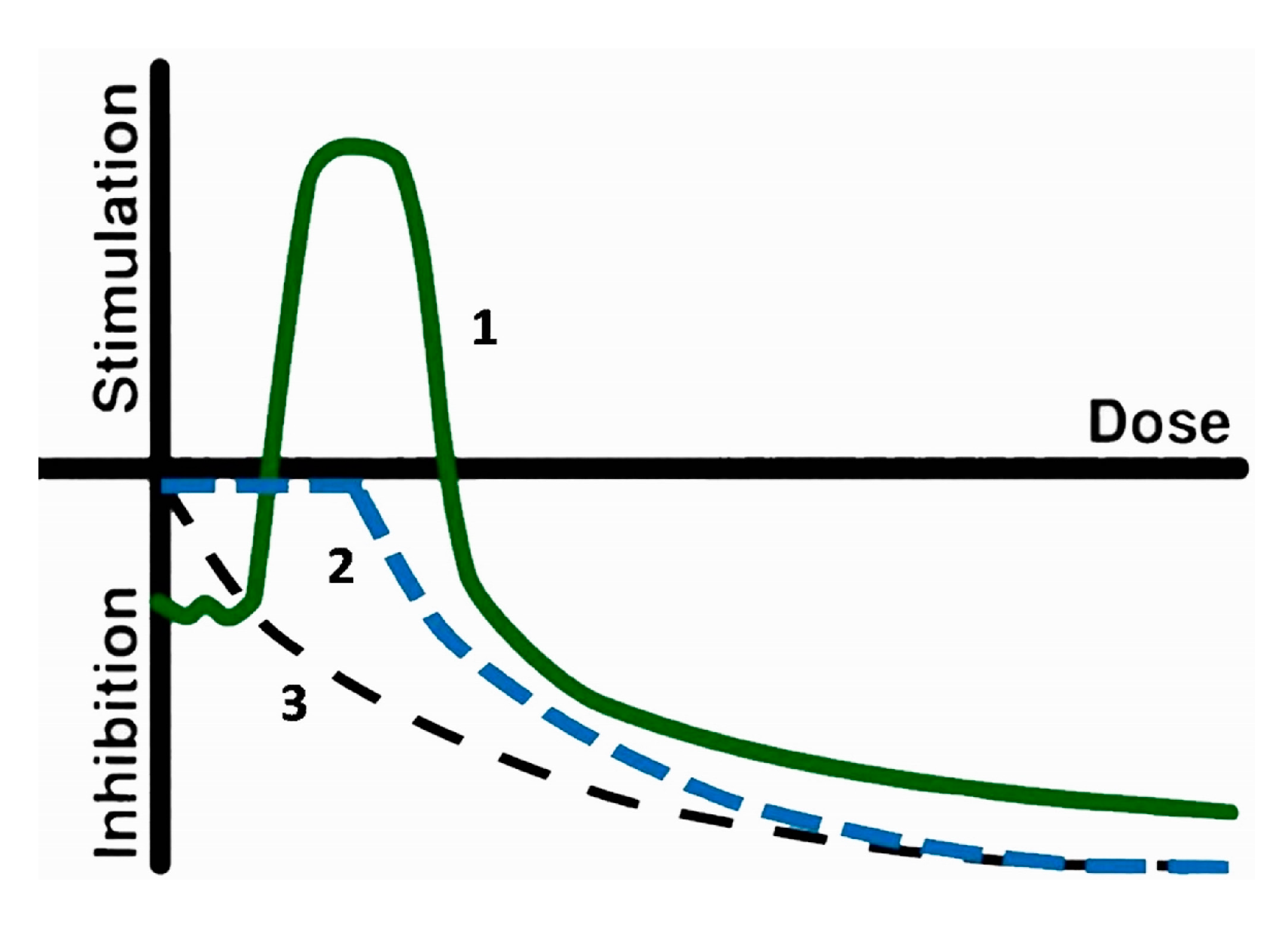{kind=link}
{kind=link}
{kind=link}
{kind=link}
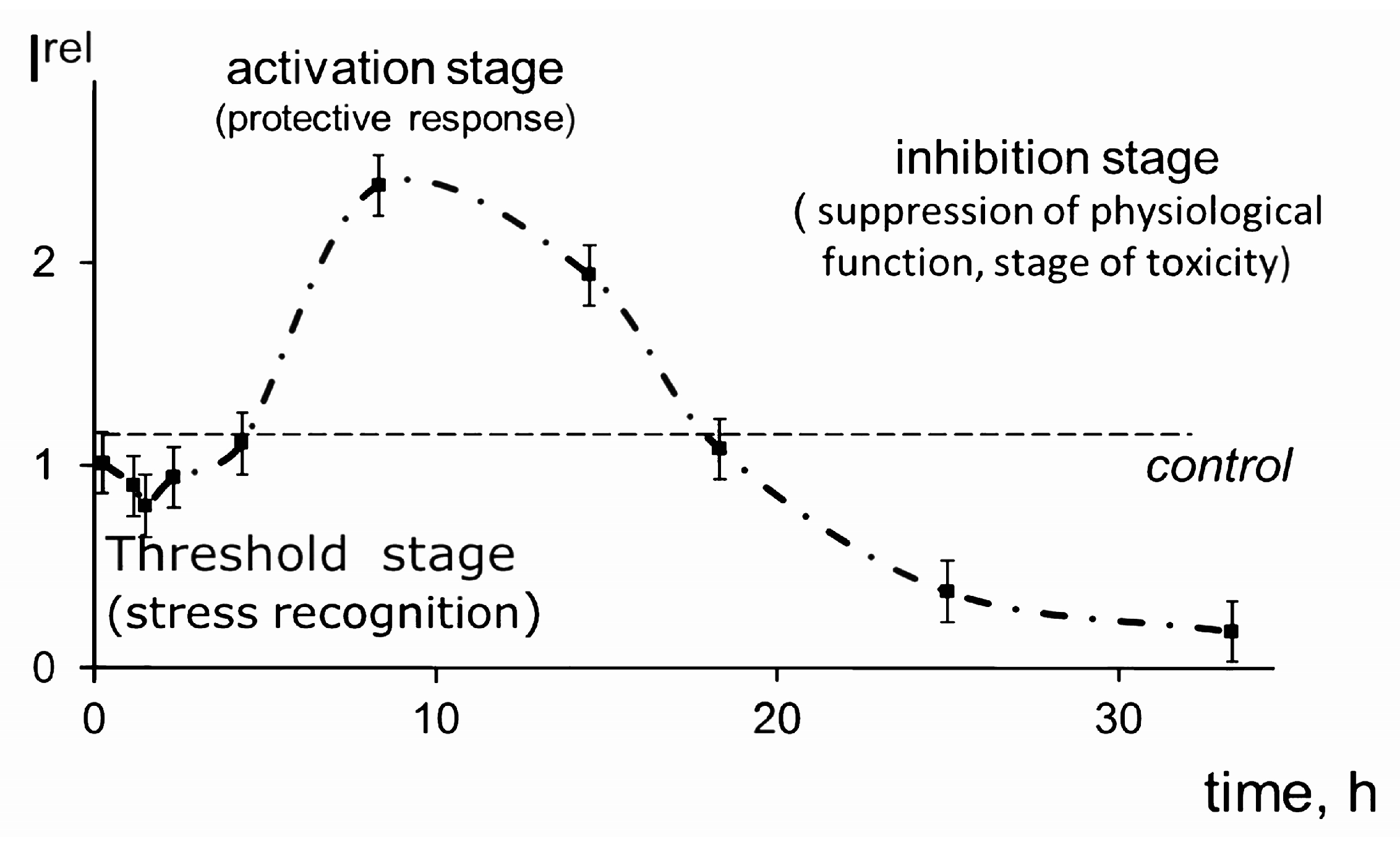{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
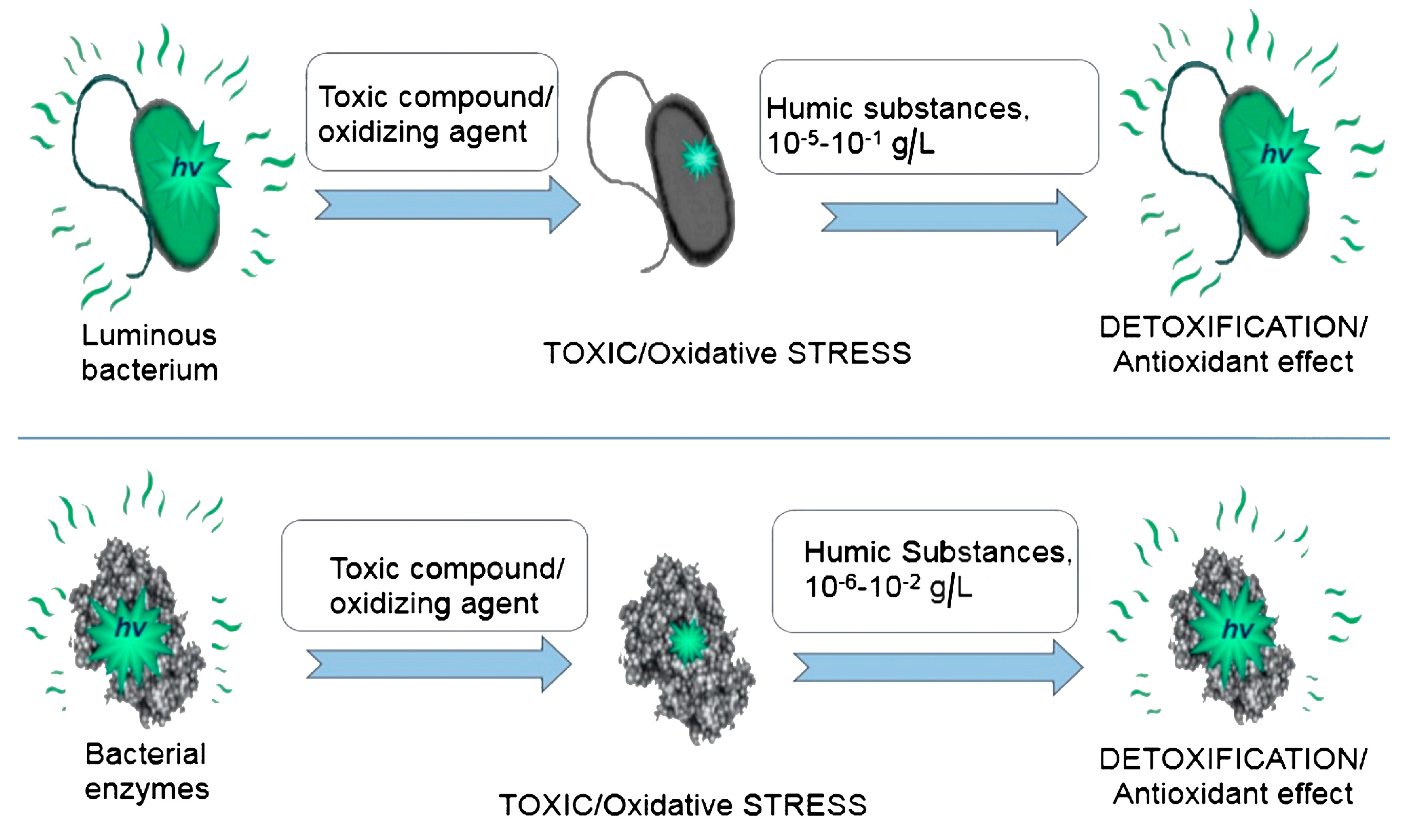{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Coelenteramide-Containing Fluorescent Protein as the Simplest Multicolor Fluorescent Bioassay
3. Luminescence Bioassays as Tools for Studying Low-Dose Radiation Effects
4. Low-Concentration Effects of Bioactive Compounds: Antioxidant Activity via Bioluminescence Bioassays
- High rates of test procedure, which
- provide statistical reliability of the bioassay results and
- can exclude the genetic level of analysis, appealing to biochemical, chemical, and physicochemical processes in cells;
- The possibility to compare effects of bioactive compounds at different organization levels—cellular and enzymatic.
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| CLM | Coelenteramide |
| DRIFT | Diffuse reflectance infrared Fourier transform |
| F | Fullerenol |
| FMN | Flavin mononucleotide |
| GT | General toxicity |
| HS | Humic substances |
| NADH | Nicotinamide adenine dinucleotide |
| OxT | Oxidative toxicity |
| ROS | Reactive oxygen species |
References
- Southam, C.M.; Ehrlich, J. Effects of extracts of western red-cedar heartwood on certain wood-decaying fungi in culture. Phytopathology 1943, 33, 517–524. [Google Scholar]
- Calabrese, E.J. Hormesis: Path and Progression to Significance. Int. J. Mol. Sci. 2018, 19, 2871. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormetic mechanisms. Crit. Rev. Toxicol. 2013, 43, 580–606. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormesis: A fundamental concept in biology. Microb. Cell 2014, 1, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Iavicoli, I.; Leso, V.; Fontana, L.; Calabrese, E.J. Nanoparticle exposure and hormetic dose–responses: An update. Int. J. Mol. Sci. 2018, 19, 805. [Google Scholar] [CrossRef] [PubMed]
- Vasilenko, I.Y.; Vasilenko, O.I. Radiation risk when exposed to small doses is negligible. Atomic Energy Bull. 2001, 12, 34–37. (In Russian) [Google Scholar]
- Ilyin, L.A.; Kutsenko, S.A.; Savateev, N.V.; Sofronov, G.A.; Tiunov, L.A. Toxicological problems in mitigation strategies of chemical industries. J. All-Union Mendeleev Chem. Soc. 1990, 35, 440–447. (In Russian) [Google Scholar]
- Tigini, V.; Giansanti, P.; Mangiavillano, A.; Pannocchia, A.; Varese, G.C. Evaluation of toxicity, genotoxicity and environmental risk of simulated textile and tannery wastewaters with a battery of biotests. Ecotoxicol. Environ. Saf. 2011, 74, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L. Bioassays as a tool for evaluating advanced oxidation processes in water and wastewater treatment. Water Res. 2011, 45, 4311–4340. [Google Scholar] [CrossRef]
- Petukhov, V.N.; Fomchenkov, V.M.; Chugunov, V.A.; Kholodenko, V.P. Plant Biotests for Soil and Water Contaminated with Oil and Oil Products. Appl. Biochem. Microbiol. 2000, 36, 564–567. [Google Scholar] [CrossRef]
- Donnelly, K.C.; Chen, J.C.; Huebner, H.J.; Brown, K.W.; Autenrieth, R.L.; Bonner, J.S. Utility of four strains of white-rot fungi for the detoxification of 2,4,6-trinitrotoluene in liquid culture. Environ. Toxicol. Chem. 1997, 16, 1105–1110. [Google Scholar] [CrossRef]
- Kudryasheva, N.S.; Tarasova, A.S. Pollutant toxicity and detoxification by humic substances: Mechanisms and quantitative assessment via luminescent Biomonitoring. Environ. Sci. Pollut. Res. Int. 2015, 22, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Girotti, S.; Ferri, E.N.; Fumo, M.G.; Maiolini, E. Monitoring of environmental pollutants by bioluminescent bacteria. Anal. Chim. Acta 2008, 608, 2–29. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.; Kratasyuk, V.; Esimbekova, E.; Vetrova, E.; Nemtseva, E.; Kudinova, I. Development of bioluminescent bioindicators for analyses of environmental pollution. Field Anal. Chem. Tech. 1998, 2, 277–280. [Google Scholar] [CrossRef]
- Roda, A.; Pasini, P.; Mirasoni, M.; Michchelini, E.; Guardigli, M. Biotechnological application of bioluminescence and chemiluminescence. Trends Biotech. 2004, 22, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Bulich, A.A.; Isenberg, D.L. Use of the luminescent bacterial system for rapid assessment of aquatic toxicity. ISA Trans. 1981, 20, 29–33. [Google Scholar] [PubMed]
- Abbas, M.; Adil, M.; Ehtisham-Ul-Haque, S.; Munir, B.; Yameen, M.; Ghaffar, A.; Shar, G.A.; Asif Tahir, M.; Iqbal, M. Vibrio fischeri bioluminescence inhibition assay for ecotoxicity assessment: A review. Sci. Total Environ. 2018, 626, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, E.; Kudryasheva, N.; Kuznetsov, A.; Mogil’naya, O.; Stom, D. Bioluminescent monitoring of detoxification processes: Activity of humic substances in quinone solutions. J. Photochem. Photobiol. B 2007, 88, 131–136. [Google Scholar] [CrossRef]
- Kudryasheva, N.S.; Rozhko, T.V. Effect of low-dose ionizing radiation on luminous marine bacteria: Radiation hormesis and toxicity. J. Environ. Radioact. 2015, 142, 68–77. [Google Scholar] [CrossRef]
- Kratasyuk, V.A.; Esimbekova, E.N. Applications of luminous bacteria enzymes in toxicology. Comb. Chem. High Throughput Screen. 2015, 18, 952–959. [Google Scholar] [CrossRef] [Green Version]
- Rozhko, T.V.; Kudryasheva, N.S.; Kuznetsov, A.M.; Vydryakova, G.A.; Bondareva, L.G.; Bolsunovsky, A.Y. Effect of low-level α-radiation on bioluminescent assay systems of various complexity. Photochem. Photobiol. Sci. 2007, 6, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Selivanova, M.A.; Mogilnaya, O.A.; Badun, G.A.; Vydryakova, G.A.; Kuznetsov, A.M.; Kudryasheva, N.S. Effect of tritium on luminous marine bacteria and enzyme reactions. J. Environ. Radioact. 2013, 120, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Esimbekova, E.N.; Kondik, A.M.; Kratasyuk, V.A. Bioluminescent enzymatic rapid assay of water integral toxicity. Environ. Monit. Assess. 2013, 185, 5909–5916. [Google Scholar] [CrossRef] [PubMed]
- Efremenko, E.N.; Maslova, O.V.; Kholstov, A.V.; Senko, O.V.; Ismailov, A.D. Biosensitive element in the form of immobilized luminescent photobacteria for detecting ecotoxicants in aqueous flow-through systems. Luminescence 2016, 31, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Ismailov, A.D.; Aleskerova, L.E. Photobiosensors containing luminescent bacteria. Biochemistry 2015, 80, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Rastogi, N.K.; Thakur, M.S. Development of immobilized biophotonic beads consisting of Photobacterium leiognathi for the detection of heavy metals and pesticide. J. Hazard Mater. 2012, 225–226, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S. Bioluminescence and exogenous compounds: Physicochemical basis for bioluminescence assay. J. Photochem. Photobiol. B 2006, 83, 77–86. [Google Scholar] [CrossRef]
- Kirillova, T.N.; Kudryasheva, N.S. Effect of heavy atom in bioluminescent reactions. Anal. Bioanal. Chem. 2007, 387, 2009–2016. [Google Scholar] [CrossRef]
- Kirillova, T.N.; Gerasimova, M.A.; Nemtseva, E.V.; Kudryasheva, N.S. Effect of halogenated fluorescent compounds on bioluminescent reactions. Anal. Bioanal. Chem. 2011, 400, 343–351. [Google Scholar] [CrossRef]
- Nemtseva, E.V.; Kudryasheva, N.S. The mechanism of electronic excitation in bacterial bioluminescent reaction. Russ. Chem. Rev. 2007, 76, 91–100. [Google Scholar] [CrossRef]
- Vetrova, E.V.; Kudryasheva, N.S.; Kratasyuk, V.A. Redox compounds influence on the NAD(P)H:FMN-oxidoreductase-luciferase bioluminescent system. Photochem. Photobiol. Sci. 2007, 6, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M.; Smith, M.T. Epidemiology. Environment and disease risks. Science 2010, 330, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P. The exposome: From concept to utility. Int. J. Epidemiol. 2012, 41, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Siroux, V.; Agier, L.; Slama, R. The exposome concept: A challenge and a potential driver for environmental health research. Eur. Respir. Rev. 2016, 25, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P. Complementing the Genome with an “Exposome”: The Outstanding Challenge of Environmental Exposure Measurement in Molecular Epidemiology. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef] [PubMed]
- Zubova, N.N.; Bulavina, A.Y.; Savitsky, A.P. Spectral and physicochemical properties of green (GFP) and red (drFP583) fluorescent proteins. Usp. Biol. Khim. 2003, 43, 163–224. (In Russian) [Google Scholar]
- Stepanenko, O.V.; Verkhusha, V.V.; Kuznetsova, I.M.; Turoverov, K.K. Fluorescent proteins: Physical-chemical properties and application in cell biology. TSITOLOGIYA 2007, 49, 395–420. [Google Scholar]
- Kumar, A.; Pal, D. Green fluorescent protein and their applications in advance research. Res. J. Appl. Sci. Eng. Tech. 2016, 1, 42–46. [Google Scholar]
- Remington, S.J. Green fluorescent protein: A perspective. Protein Sci. 2011, 20, 1509–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cody, C.W.; Prasher, D.C.; Wester, W.W.; Prendergast, F.G.; Ward, W.W. Chemical structure of the hexapeptide chromophore of the Aequorea green-fluorescent protein. Biochemistry 1993, 32, 1212–1218. [Google Scholar] [CrossRef]
- Ormö, M.; Cubitt, A.B.; Kallio, K.; Gross, L.A.; Tsien, R.Y.; Remington, S.J. Crystal structure of the Aequorea victoria green fluorescent protein. Science 1996, 273, 1392–1395. [Google Scholar] [CrossRef] [PubMed]
- Frank, L.A. Ca2+-Regulated photoproteins: Effective immunoassay reporters. Sensors 2010, 10, 11287–11300. [Google Scholar] [CrossRef] [PubMed]
- Krasitskaya, V.V.; Burakova, L.P.; Pyshnaya, I.A.; Frank, L.A. Bioluminescent reporters for identification of gene allelic variants. Russ. J. Bioorg. Chem. 2012, 38, 298–305. [Google Scholar] [CrossRef]
- Shimomura, O.; Teranishi, K. Light-emitters involved in the luminescence of coelenterazine. Luminescence 2000, 15, 51–58. [Google Scholar] [CrossRef]
- Li, Z.S.; Zhao, X.; Zou, L.Y.; Ren, A.M. The Dynamics Simulation and Quantum Calculation Investigation about Luminescence Mechanism of Coelenteramide. Photochem. Photobiol. 2013, 89, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Malikova, N.P.; Stepanyuk, G.A.; Frank, L.A.; Markova, S.V.; Vysotski, E.S.; Lee, J. Spectral tuning of obelin bioluminescence by mutations of Trp92. FEBS Lett. 2003, 554, 184–188. [Google Scholar] [CrossRef]
- Belogurova, N.V.; Kudryasheva, N.S.; Alieva, R.R. Activity of upper electron-excited states in bioluminescence of coelenterates. J. Mol. Struct. 2009, 924, 148–152. [Google Scholar] [CrossRef]
- Sharifian, S.; Homaei, A.; Hemmati, R.; Luwor, R.B.; Khajeh, K. The emerging use of bioluminescence in medical research. Biomed. Pharmacother. 2018, 101, 74–86. [Google Scholar] [CrossRef]
- Lee, J. Perspectives on Bioluminescence Mechanisms. Photochem. Photobiol. 2017, 93, 389–404. [Google Scholar] [CrossRef]
- Alieva, R.R.; Tomilin, F.N.; Kuzubov, A.A.; Ovchinnikov, S.G.; Kudryasheva, N.S. Ultraviolet fluorescence of coelenteramide and coelenteramide-containing fluorescent proteins. Experimental and theoretical study. J. Photochem. Photobiol. B 2016, 162, 318–323. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.F.; Ferre, N.; Liu, Y.J. QM/MM Study on the Light Emitters of Aequorin Chemiluminescence, Bioluminescence, and Fluorescence: A General Understanding of the Bioluminescence of Several Marine Organisms. Chem. Eur. J. 2013, 19, 8466–8472. [Google Scholar] [CrossRef] [PubMed]
- Min, C.G.; Li, Z.S.; Ren, A.M.; Zou, L.Y.; Guo, J.F.; Goddard, J.D. The fluorescent properties of coelenteramide, a substrate of aequorin and obelin. J. Photochem. Photobiol. A 2013, 251, 182–188. [Google Scholar] [CrossRef]
- Van Oort, B.; Eremeeva, E.V.; Koehorst, R.B.M.; Laptenok, S.P.; van Amerongen, H.; van Berkel, W.J.H.; Malikova, N.P.; Markova, S.V.; Vysotski, E.S.; Visser, A.J.W.G.; et al. Picosecond Fluorescence Relaxation Spectroscopy of the Calcium-Discharged Photoproteins Aequorin and Obelin. Biochemistry 2009, 48, 10486–10491. [Google Scholar] [CrossRef] [PubMed]
- Belogurova, N.V.; Kudryasheva, N.S.; Alieva, R.R.; Sizykh, A.G. Spectral components of bioluminescence of aequorin and obelin. J. Photochem. Photobiol. B 2008, 92, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Belogurova, N.V.; Kudryasheva, N.S. Discharged photoprotein Obelin: Fluorescence peculiarities. J. Photochem. Photobiol. B 2010, 101, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Alieva, R.R.; Belogurova, N.V.; Petrova, A.S.; Kudryasheva, N.S. Fluorescence properties of Ca2+-independent discharged obelin and its application prospects. Anal. Bioanal. Chem. 2013, 405, 3351–3358. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Liu, Y.J. Photoluminescence Rainbow from Coelenteramide—A Theoretical Study. Photochem. Photobiol. 2018, 95, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Alieva, R.R.; Belogurova, N.V.; Petrova, A.S.; Kudryasheva, N.S. Effects of alcohols on fluorescence intensity and color of a discharged-obelin-based biomarker. Anal. Bioanal. Chem. 2014, 406, 2965–2974. [Google Scholar] [CrossRef] [PubMed]
- Petrova, A.S.; Alieva, R.R.; Belogurova, N.V.; Tirranen, L.S.; Kudryasheva, N.S. Variation of spectral characteristics of coelenteramide-containing fluorescent protein from Obelia longissima exposed to Dimethyl sulfoxide. Russ. Phys. J. 2016, 59, 87–92. [Google Scholar] [CrossRef]
- Alexandrova, M.; Rozhko, T.; Vydryakova, G.; Kudryasheva, N. Effect of americium-241 on luminous bacteria. Role of peroxides. J. Environ. Radioact. 2011, 102, 407–411. [Google Scholar] [CrossRef]
- Selivanova, M.A.; Rozhko, T.V.; Devyatlovskaya, A.N.; Kudryasheva, N.S. Comparison of chronic low-dose effects of alpha-and beta-emitting radionuclides on marine bacteria. Cent. Eur. J. Biol. 2014, 9, 951–959. [Google Scholar] [CrossRef]
- Alieva, R.R.; Kudryasheva, N.S. Variability of fluorescence spectra of coelenteramide-containing proteins as a basis for toxicity monitoring. Talanta 2017, 170, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Burlakova, E.B.; Konradov, A.A.; Maltseva, E.X. Effect of extremely weak chemical and physical stimuli on biological systems. Biophysics (Moscow) 2004, 49, 522–534. [Google Scholar]
- Feinendegen, L.E.; Pollycove, M.; Neumann, R.D. Whole-body responses to low-level radiation exposure: New concepts in mammalian radiobiology. Exp. Hematol. 2007, 35, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Feinendegen, L.E. Evidence for beneficial low level radiation effects and radiation hormesis. Br. J. Radiol. 2005, 78, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Seymour, C. Implications for human and environmental health of low doses of ionising radiation. J. Environ. Radioact. 2014, 133, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.R.; Jung, P.M.; Choi, J.; Lee, J.-W. Radiation sensitivity of bacteria and virus in porcine xenoskin for dressing agent. Radiat. Phys. Chem. 2012, 81, 1259–1262. [Google Scholar] [CrossRef]
- Mesquita, N.; Portugal, A.; Piñar, G.; Loureiro, J.; Coutinho, A.P.; Trovão, J.; Nunes, I.; Botelho, M.L.; Freitas, H. Flow cytometry as a tool to assess the effects of gamma radiation on the viability, growth and metabolic activity of fungal spores. Int. Biodeter. Biodegr. 2013, 84, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.; Kadam, A.A.; Govindwar, S.P.; Kumar, P.; Varshney, L. An insight into the influence of low dose irradiation pretreatment on the microbial decolouration and degradation of Reactive Red-120 dye. Chemosphere 2013, 90, 1348–1358. [Google Scholar] [CrossRef]
- Xavier, M.P.; Dauber, C.; Mussio, P.; Delgado, E.; Maquieira, A.; Soria, A.; Curuchet, A.; Márquez, R.; Méndez, C.; López, T. Use of mild irradiation doses to control pathogenic bacteria on meat trimmings for production of patties aiming at provoking minimal changes in quality attributes. Meat. Sci. 2014, 98, 383–391. [Google Scholar] [CrossRef]
- Luckey, T.D. Hormesis with Ionizing Radiation; CRC Press, Incorporated: Boca Raton, FL, USA, 1980; p. 225. [Google Scholar]
- Kudryasheva, N.S.; Petrova, A.S.; Dementyev, D.V.; Bondar, A.A. Exposure of luminous marine bacteria to low-dose gamma-radiation. J. Environ. Radioact. 2017, 169–170, 64–69. [Google Scholar] [CrossRef]
- Rozhko, T.V.; Badun, G.A.; Razzhivina, I.A.; Guseynov, O.A.; Guseynova, V.E.; Kudryasheva, N.S. On mechanism of biological activation by tritium. J. Environ. Radioact. 2016, 157, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Rozhko, T.V.; Guseynov, O.A.; Guseynova, V.E.; Bondar, A.A.; Devyatlovskaya, A.N.; Kudryasheva, N.S. Is bacterial luminescence response to low-dose radiation associated with mutagenicity? J. Environ. Radioact. 2017, 177, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, R.W. Biochemical aspects of active transport. Annu. Rev. Biochem. 1967, 36, 727–756. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D.C.; Edvards, A.A.; Leonard, A.; Deknut, G.L.; Verschaeve, L.; Natarajan, A.T.; Darrudi, F.; Obe, G.; Palitti, F.; Tanzarella, C.; et al. Chromosomal aberrations in human lymphocytes induced in vitro by very low doses of X-rays. Int. J. Radiat. Biol. 1992, 61, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.E. Impact of 16S rRNA Gene Sequence Analysis for Identification of Bacteria on Clinical Microbiology and Infectious Diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Jia, Z.; Trush, M.A. Defining ROS in Biology and Medicine. React. Oxyg. Species (Apex) 2016, 1, 9–21. [Google Scholar] [CrossRef]
- Vallejo, M.J.; Salazar, L.; Grijalva, M. Oxidative Stress Modulation and ROS-Mediated Toxicity in Cancer: A Review on In Vitro Models for Plant-Derived Compounds. Oxid. Med. Cell. Longev. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Fedorova, G.F.; Menshov, V.A.; Trofimov, A.V.; Tsaplev, Y.B.; Vasil’ev, R.F.; Yablonskaya, O.I. Chemiluminescence of cigarette smoke: Salient features of the phenomenon. Photochem. Photobiol. 2017, 93, 579–589. [Google Scholar] [CrossRef]
- Rozhko, T.; Nogovitsyna, E.; Badun, G.; Lukyanchuk, A.; Kudryasheva, N. Reactive Oxygen Species and Low-Dose Effects of Tritium on Bacterial Cells. J. Environ. Radioact. 2019, 208–209, 106035. [Google Scholar] [CrossRef]
- Audi, G.; Wapstra, A.H.; Thibault, C. The AME2003 atomic mass evaluation (II).Tables, graphs, and references. Nucl. Phys. A 2003, 729, 337–676. [Google Scholar] [CrossRef]
- Kamnev, A.A.; Tugarova, A.V.; Selivanova, M.A.; Tarantilis, P.A.; Polissiou, M.G.; Kudryasheva, N.S. Effects of americium-241 and humic substances on Photobacterium phosphoreum: Bioluminescence and diffuse reflectance FTIR spectroscopic studies. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 100, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Kamnev, A.A.; Tugarova, A.V.; Alexandrova, M.A.; Tarantilis, P.A.; Polissiou, M.G.; Kudryasheva, N.S. Effects of tritium and humic substances on cells of Photobacterium phosphoreum: Bioluminescence and FTIR spectroscopic studies. Program and Abstr. Book, Spectroscopy in the Biological Sciences of the XXI Century. In Proceedings of the Intracellular Fluorescence Spectroscopy (A Satellite Conf. of the 8th Eur. Biophysics Congress), Pecs, Hungary, 20–22 August 2011; Abstr. No. P8. p. 35. [Google Scholar]
- Kamnev, A.A.; Sadovnikova, J.N.; Tarantilis, P.A.; Polissiou, M.G.; Antonyuk, L.P. Responses of Azospirillum brasilense to nitrogen deficiency and to wheat lectin: A diffuse reflectance infrared Fourier transform (DRIFT) spectroscopic study. Microb. Ecol. 2008, 56, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Lee, C.W.; Gu, M.B. Gamma-radiation dose-rate effects on DNA damage and toxicity in bacterial cells. Radiat. Environ. Biophys. 2003, 42, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Petrova, A.S.; Lukonina, A.A.; Badun, G.A.; Kudryasheva, N.S. Fluorescent Coelenteramide-Containing Protein as a Color Bioindicator for Low-Dose Radiation Effects. Anal. Bioanal. Chem. 2017, 409, 4377–4381. [Google Scholar] [CrossRef] [PubMed]
- Petrova, A.S.; Lukonina, A.A.; Dementyev, D.V.; Bolsunovsky, A.Y.; Popov, A.V.; Kudryasheva, N.S. Protein-based fluorescent bioassay for low-dose gamma radiation exposures. Anal. Bioanal. Chem. 2018, 410, 6837–6844. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. Changing distress into eustress: Voices theories on stress. Tex. Med. 1980, 76, 78–80. [Google Scholar]
- Wang, C.R.; Tian, Y.; Wang, X.R.; Yu, H.X.; Lu, X.W.; Wang, C.; Wang, H. Hormesis effects and implicative application in assessment of lead-contaminated soils in roots of Vicia faba seedlings. Chemosphere 2010, 80, 965–971. [Google Scholar] [CrossRef]
- Baldwin, J.; Grantham, V. Radiation hormesis: Historical and current perspectives. J. Nucl. Med. Technol. 2015, 43, 242–246. [Google Scholar] [CrossRef]
- Kaiser, J. Hormesis: Sipping from a poisoned chalice. Science 2003, 302, 376–379. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. The frequency of U-shaped dose responses in the toxicological literature. Toxicol. Sci. 2001, 62, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Kleinhempel, D. Ein beitrag zur theories des huminstoffzustandes. Arch. Agron. Soil Sci. 1970, 14, 3–14. [Google Scholar] [CrossRef]
- Tarasova, A.S.; Stom, D.I.; Kudryasheva, N.S. Effect of humic substances on toxicity of inorganic oxidizer bioluminescent monitoring. Environ. Toxicol. Chem. 2011, 30, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Tarasova, A.S.; Kislan, S.L.; Fedorova, E.S.; Kuznetsov, A.M.; Mogilnaya, O.A.; Stom, D.I.; Kudryasheva, N.S. Bioluminescence as a tool for studying detoxification processes in metal salt solutions involving humic substances. J. Photochem. Photobiol. B 2012, 117, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Tarasova, A.S.; Stom, D.I.; Kudryasheva, N.S. Antioxidant activity of humic substances via bioluminescent monitoring in vitro. Environ. Monit. Assess. 2015, 187, 89. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S.; Kovel, E.S.; Sachkova, A.S.; Vorobeva, A.A.; Isakova, V.G.; Churilov, G.N. Bioluminescent enzymatic assay as a tool for studying antioxidant activity and toxicity of bioactive compounds. J. Photochem. Photobiol. 2017, 93, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Sachkova, A.S.; Kovel, E.S.; Churilov, G.N.; Guseynov, O.A.; Bondar, A.A.; Dubinina, I.A.; Kudryasheva, N.S. On mechanism of antioxidant effect of fullerenols. Biochem. Biophys. Rep. 2017, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sachkova, A.S.; Kovel, E.S.; Churilov, G.N.; Stom, D.I.; Kudryasheva, N.S. Biological activity of carbonic nano-structures—Comparison via enzymatic bioassay. J. Soils Sediments 2018. [Google Scholar] [CrossRef]
- Kovel, E.S.; Sachkova, A.S.; Vnukova, N.G.; Churilov, G.N.; Knyazeva, E.M.; Kudryasheva, N.S. Antioxidant Activity and Toxicity of Fullerenols via Bioluminescence Signaling: Role of Oxygen Substituents. Int. J. Mol. Sci. 2019, 20, 2324. [Google Scholar] [CrossRef]
- Kudryasheva, N.; Vetrova, E.; Kuznetsov, A.; Kratasyuk, V.; Stom, D. Bioluminescent assays: Effects of quinones and phenols. Ecotoxicol. Environ. Saf. 2002, 53, 221–225. [Google Scholar] [CrossRef]
- Brisebois, P.P.; Arnold, A.A.; Chabre, Y.M.; Roy, R.; Marcotte, I. Comparative study of the interaction of fullerenol nanoparticles with eukaryotic and bacterial model membranes using solid-state NMR and FTIR spectroscopy. Eur. Biophys. J. 2012, 41, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Voeikov, V.L.; Yablonskaya, O.I. Stabilizing effects of hydrated fullerenes C60 in a wide range of concentrations on luciferase, alkaline phosphatase, and peroxidase in vitro. Electromagn. Biol. Med. 2015, 34, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Yablonskaya, O.I.; Ryndina, T.S.; Voeikov, V.L.; Khokhlov, A.N. A paradoxal effect of hydrated C60-fullerene in an ultralow concentration on the viability and aging of cultivated Chinese hamster cells. Moscow Univ. Biol. Sci. Bull. 2013, 68, 63–68. [Google Scholar] [CrossRef]
- Bensasson, R.V.; Bretteich, M.; Frederiksen, J.; Gottinger, H.; Hirsch, A.; Land, E.J.; Leach, S.; McGarvey, D.J.; Schonberger, H. Reactions of e-aq, CO2−, HO−, O2− and O2(1Dg) with a dendro [60] fullerene and C60[C(COOH)2]n (n = 2–6). Free Radic. Biol. Med. 2000, 29, 26–33. [Google Scholar] [CrossRef]












© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudryasheva, N.S.; Kovel, E.S. Monitoring of Low-Intensity Exposures via Luminescent Bioassays of Different Complexity: Cells, Enzyme Reactions, and Fluorescent Proteins. Int. J. Mol. Sci. 2019, 20, 4451. https://doi.org/10.3390/ijms20184451
Kudryasheva NS, Kovel ES. Monitoring of Low-Intensity Exposures via Luminescent Bioassays of Different Complexity: Cells, Enzyme Reactions, and Fluorescent Proteins. International Journal of Molecular Sciences. 2019; 20(18):4451. https://doi.org/10.3390/ijms20184451
Chicago/Turabian StyleKudryasheva, Nadezhda S., and Ekaterina S. Kovel. 2019. "Monitoring of Low-Intensity Exposures via Luminescent Bioassays of Different Complexity: Cells, Enzyme Reactions, and Fluorescent Proteins" International Journal of Molecular Sciences 20, no. 18: 4451. https://doi.org/10.3390/ijms20184451
APA StyleKudryasheva, N. S., & Kovel, E. S. (2019). Monitoring of Low-Intensity Exposures via Luminescent Bioassays of Different Complexity: Cells, Enzyme Reactions, and Fluorescent Proteins. International Journal of Molecular Sciences, 20(18), 4451. https://doi.org/10.3390/ijms20184451