The Endocrine Function of Osteocalcin Regulated by Bone Resorption: A Lesson from Reduced and Increased Bone Mass Diseases

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Disorders of Altered Bone Resorption
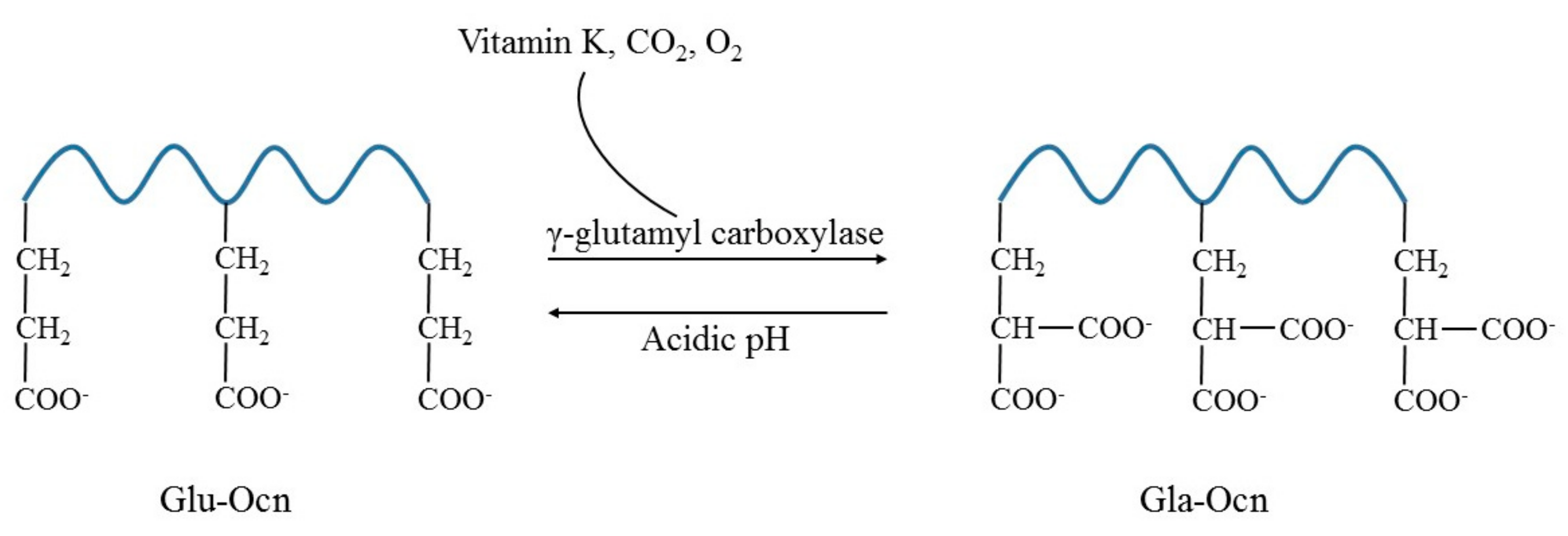
3. Osteocalcin
4. Glucose Metabolism
5. Male Fertility
6. Brain Functions
7. Conclusions
Funding
Conflicts of Interest
References
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Ferron, M.; Hinoi, E.; Karsenty, G.; Ducy, P. Osteocalcin differentially regulates beta cell and adipocyte gene expression and affects the development of metabolic diseases in wild-type mice. Proc. Natl. Acad. Sci. USA 2008, 105, 5266–5270. [Google Scholar] [CrossRef] [PubMed]
- Ferron, M.; Wei, J.; Yoshizawa, T.; Del Fattore, A.; DePinho, R.A.; Teti, A.; Ducy, P.; Karsenty, G. Insulin signaling in osteoblasts integrates bone remodeling and energy metabolism. Cell 2010, 142, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Oury, F.; Sumara, G.; Sumara, O.; Ferron, M.; Chang, H.; Smith, C.E.; Hermo, L.; Suarez, S.; Roth, B.L.; Ducy, P.; et al. Endocrine regulation of male fertility by the skeleton. Cell 2011, 144, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Mera, P.; Laue, K.; Ferron, M.; Confavreux, C.; Wei, J.; Galan-Diez, M.; Lacampagne, A.; Mitchell, S.J.; Mattison, J.A.; Chen, Y.; et al. Osteocalcin Signaling in Myofibers Is Necessary and Sufficient for Optimum Adaptation to Exercise. Cell Metab. 2016, 23, 1078–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mera, P.; Laue, K.; Wei, J.; Berger, J.M.; Karsenty, G. Osteocalcin is necessary and sufficient to maintain muscle mass in older mice. Mol. Metab. 2016, 5, 1042–1047. [Google Scholar] [CrossRef]
- Oury, F.; Khrimian, L.; Denny, C.A.; Gardin, A.; Chamouni, A.; Goeden, N.; Huang, Y.Y.; Lee, H.; Srinivas, P.; Gao, X.B.; et al. Maternal and offspring pools of osteocalcin influence brain development and functions. Cell 2013, 155, 228–241. [Google Scholar] [CrossRef]
- Vaananen, H.K.; Laitala-Leinonen, T. Osteoclast lineage and function. Arch. Biochem. Biophys. 2008, 473, 132–138. [Google Scholar] [CrossRef]
- Hadjidakis, D.J.; Androulakis, II. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]
- Hill, P.A. Bone remodelling. Br. J. Orthod. 1998, 25, 101–107. [Google Scholar] [CrossRef]
- Matsuo, K.; Irie, N. Osteoclast-osteoblast communication. Arch. Biochem. Biophys. 2008, 473, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Rousselle, A.V.; Heymann, D. Osteoclastic acidification pathways during bone resorption. Bone 2002, 30, 533–540. [Google Scholar] [CrossRef]
- Graves, A.R.; Curran, P.K.; Smith, C.L.; Mindell, J.A. The Cl-/H+ antiporter ClC-7 is the primary chloride permeation pathway in lysosomes. Nature 2008, 453, 788–792. [Google Scholar] [CrossRef]
- Kornak, U.; Kasper, D.; Bosl, M.R.; Kaiser, E.; Schweizer, M.; Schulz, A.; Friedrich, W.; Delling, G.; Jentsch, T.J. Loss of the ClC-7 chloride channel leads to osteopetrosis in mice and man. Cell 2001, 104, 205–215. [Google Scholar] [CrossRef]
- Everts, V.; Korper, W.; Hoeben, K.A.; Jansen, I.D.; Bromme, D.; Cleutjens, K.B.; Heeneman, S.; Peters, C.; Reinheckel, T.; Saftig, P.; et al. Osteoclastic bone degradation and the role of different cysteine proteinases and matrix metalloproteinases: Differences between calvaria and long bone. J. Bone Miner. Res. 2006, 21, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Del Fattore, A.; Teti, A.; Rucci, N. Bone cells and the mechanisms of bone remodelling. Front. Biosci. (Elite Ed.) 2012, 4, 2302–2321. [Google Scholar] [CrossRef]
- Consensus development conference: Diagnosis, prophylaxis, and treatment of osteoporosis. Am. J. Med. 1993, 94, 646–650. [CrossRef]
- Colangelo, L.; Biamonte, F.; Pepe, J.; Cipriani, C.; Minisola, S. Understanding and managing secondary osteoporosis. Expert Rev. Endocrinol. Metab. 2019, 14, 111–122. [Google Scholar] [CrossRef]
- Garnero, P.; Sornay-Rendu, E.; Chapuy, M.C.; Delmas, P.D. Increased bone turnover in late postmenopausal women is a major determinant of osteoporosis. J. Bone Miner. Res. 1996, 11, 337–349. [Google Scholar] [CrossRef]
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Imaz, I.; Zegarra, P.; Gonzalez-Enriquez, J.; Rubio, B.; Alcazar, R.; Amate, J.M. Poor bisphosphonate adherence for treatment of osteoporosis increases fracture risk: Systematic review and meta-analysis. Osteoporos. Int. 2010, 21, 1943–1951. [Google Scholar] [CrossRef] [PubMed]
- Siris, E.S.; Selby, P.L.; Saag, K.G.; Borgstrom, F.; Herings, R.M.; Silverman, S.L. Impact of osteoporosis treatment adherence on fracture rates in North America and Europe. Am. J. Med. 2009, 122, S3–S13. [Google Scholar] [CrossRef] [PubMed]
- Albers-Schonberg. Rontgernbilder einer seltenen Knockenerkrankung. Munc. Med. Wochenschr. 1904, 5, 365–368. [Google Scholar]
- Del Fattore, A.; Cappariello, A.; Teti, A. Genetics, pathogenesis and complications of osteopetrosis. Bone 2008, 42, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Del Fattore, A.; Peruzzi, B.; Rucci, N.; Recchia, I.; Cappariello, A.; Longo, M.; Fortunati, D.; Ballanti, P.; Iacobini, M.; Luciani, M.; et al. Clinical, genetic, and cellular analysis of 49 osteopetrotic patients: Implications for diagnosis and treatment. J. Med. Genet. 2006, 43, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Pangrazio, A.; Cassani, B.; Guerrini, M.M.; Crockett, J.C.; Marrella, V.; Zammataro, L.; Strina, D.; Schulz, A.; Schlack, C.; Kornak, U.; et al. RANK-dependent autosomal recessive osteopetrosis: Characterization of five new cases with novel mutations. J. Bone Miner. Res. 2012, 27, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Sobacchi, C.; Frattini, A.; Guerrini, M.M.; Abinun, M.; Pangrazio, A.; Susani, L.; Bredius, R.; Mancini, G.; Cant, A.; Bishop, N.; et al. Osteoclast-poor human osteopetrosis due to mutations in the gene encoding RANKL. Nat. Genet. 2007, 39, 960–962. [Google Scholar] [CrossRef]
- Villa, A.; Guerrini, M.M.; Cassani, B.; Pangrazio, A.; Sobacchi, C. Infantile malignant, autosomal recessive osteopetrosis: The rich and the poor. Calcif. Tissue Int. 2009, 84, 1–12. [Google Scholar] [CrossRef]
- Benichou, O.D.; Laredo, J.D.; de Vernejoul, M.C. Type II autosomal dominant osteopetrosis (Albers-Schonberg disease): Clinical and radiological manifestations in 42 patients. Bone 2000, 26, 87–93. [Google Scholar] [CrossRef]
- Cleiren, E.; Benichou, O.; Van Hul, E.; Gram, J.; Bollerslev, J.; Singer, F.R.; Beaverson, K.; Aledo, A.; Whyte, M.P.; Yoneyama, T.; et al. Albers-Schonberg disease (autosomal dominant osteopetrosis, type II) results from mutations in the ClCN7 chloride channel gene. Hum. Mol. Genet. 2001, 10, 2861–2867. [Google Scholar] [CrossRef] [Green Version]
- De Ridder, R.; Boudin, E.; Mortier, G.; Van Hul, W. Human Genetics of Sclerosing Bone Disorders. Curr. Osteoporos. Rep. 2018, 16, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Markatos, K.; Mavrogenis, A.F.; Karamanou, M.; Androutsos, G. Pycnodysostosis: The disease of Henri de Toulouse-Lautrec. Eur. J. Orthop. Surg. Traumatol. 2018, 28, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Motyckova, G.; Fisher, D.E. Pycnodysostosis: Role and regulation of cathepsin K in osteoclast function and human disease. Curr. Mol. Med. 2002, 2, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Hauschka, P.V.; Lian, J.B.; Cole, D.E.; Gundberg, C.M. Osteocalcin and matrix Gla protein: Vitamin K-dependent proteins in bone. Physiol. Rev. 1989, 69, 990–1047. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A. Gla-containing proteins of bone. Connect. Tissue Res. 1989, 21, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Otsuka, A.A.; Poser, J.W.; Kristaponis, J.; Raman, N. Characterization of a gamma-carboxyglutamic acid-containing protein from bone. Proc. Natl. Acad. Sci. USA 1976, 73, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Poser, J.W.; Raman, N. Primary structure of the gamma-carboxyglutamic acid-containing protein from bovine bone. Proc. Natl. Acad. Sci. USA 1976, 73, 3374–3375. [Google Scholar] [CrossRef]
- Gundberg, C.M.; Clough, M.E. The osteocalcin propeptide is not secreted in vivo or in vitro. J. Bone Miner. Res. 1992, 7, 73–80. [Google Scholar] [CrossRef]
- Gundberg, C.M.; Markowitz, M.E.; Mizruchi, M.; Rosen, J.F. Osteocalcin in human serum: A circadian rhythm. J. Clin. Endocrinol. Metab. 1985, 60, 736–739. [Google Scholar] [CrossRef]
- Morris, D.P.; Stevens, R.D.; Wright, D.J.; Stafford, D.W. Processive post-translational modification. Vitamin K-dependent carboxylation of a peptide substrate. J. Biol. Chem. 1995, 270, 30491–30498. [Google Scholar] [CrossRef]
- Gundberg, C.M.; Weinstein, R.S. Multiple immunoreactive forms of osteocalcin in uremic serum. J. Clin. Investig. 1986, 77, 1762–1767. [Google Scholar] [CrossRef] [PubMed]
- Razzaque, M.S. Osteocalcin: A pivotal mediator or an innocent bystander in energy metabolism? Nephrol. Dial. Transplant. 2011, 26, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Neve, A.; Corrado, A.; Cantatore, F.P. Osteocalcin: Skeletal and extra-skeletal effects. J. Cell. Physiol. 2013, 228, 1149–1153. [Google Scholar] [CrossRef] [PubMed]
- Ivaska, K.K.; Hentunen, T.A.; Vaaraniemi, J.; Ylipahkala, H.; Pettersson, K.; Vaananen, H.K. Release of intact and fragmented osteocalcin molecules from bone matrix during bone resorption in vitro. J. Biol. Chem. 2004, 279, 18361–18369. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Williamson, M.K.; Epstein, D.J. Specific tritium incorporation into gamma-carboxyglutamic acid in proteins. The pH dependence of gamma-proton exchange. J. Biol. Chem. 1981, 256, 1172–1176. [Google Scholar] [PubMed]
- Ducy, P. The role of osteocalcin in the endocrine cross-talk between bone remodelling and energy metabolism. Diabetologia 2011, 54, 1291–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Franco, M.C.; Franco-Diaz de Leon, R.; Villafan-Bernal, J.R. OsteocalcinGPRC6A: An update of its clinical and biological multiorganic interactions (Review). Mol. Med. Rep. 2019, 19, 15–22. [Google Scholar] [PubMed]
- Oldknow, K.J.; MacRae, V.E.; Farquharson, C. Endocrine role of bone: Recent and emerging perspectives beyond osteocalcin. J. Endocrinol. 2015, 225, R1–R19. [Google Scholar] [CrossRef]
- Arnold, K.A.; Eichelbaum, M.; Burk, O. Alternative splicing affects the function and tissue-specific expression of the human constitutive androstane receptor. Nucl. Recept. 2004, 2, 1. [Google Scholar] [CrossRef]
- Rauner, M.; Coudert, A.; Sobacchi, C.; Del Fattore, A. The endocrine role of the skeleton. Int. J. Endocrinol. 2015, 2015, 265151. [Google Scholar] [CrossRef]
- Khrimian, L.; Obri, A.; Ramos-Brossier, M.; Rousseaud, A.; Moriceau, S.; Nicot, A.S.; Mera, P.; Kosmidis, S.; Karnavas, T.; Saudou, F.; et al. Gpr158 mediates osteocalcin’s regulation of cognition. J. Exp. Med. 2017, 214, 2859–2873. [Google Scholar] [CrossRef] [PubMed]
- Kosmidis, S.; Polyzos, A.; Harvey, L.; Youssef, M.; Denny, C.A.; Dranovsky, A.; Kandel, E.R. RbAp48 Protein Is a Critical Component of GPR158/OCN Signaling and Ameliorates Age-Related Memory Loss. Cell Rep. 2018, 25, 959–973. [Google Scholar] [CrossRef] [PubMed]
- Pavlopoulos, E.; Jones, S.; Kosmidis, S.; Close, M.; Kim, C.; Kovalerchik, O.; Small, S.A.; Kandel, E.R. Molecular mechanism for age-related memory loss: The histone-binding protein RbAp48. Sci. Transl. Med. 2013, 5, 200ra115. [Google Scholar] [CrossRef] [PubMed]
- Kaji, H.; Kuroki, Y.; Murakawa, Y.; Funakawa, I.; Funasaka, Y.; Kanda, F.; Sugimoto, T. Effect of alendronate on bone metabolic indices and bone mineral density in patients treated with high-dose glucocorticoid: A prospective study. Osteoporos. Int. 2010, 21, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Yamaguchi, T.; Nawata, K.; Takaoka, S.; Sugimoto, T. Relationships between undercarboxylated osteocalcin and vitamin K intakes, bone turnover, and bone mineral density in healthy women. Clin. Nutr. 2010, 29, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Karimi Fard, M.; Aminorroaya, A.; Kachuei, A.; Salamat, M.R.; Hadi Alijanvand, M.; Aminorroaya Yamini, S.; Karimifar, M.; Feizi, A.; Amini, M. Alendronate improves fasting plasma glucose and insulin sensitivity, and decreases insulin resistance in prediabetic osteopenic postmenopausal women: A randomized triple-blind clinical trial. J. Diabetes. Investig. 2019, 10, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.V.; Schafer, A.L.; Grey, A.; Vittinghoff, E.; Palermo, L.; Lui, L.Y.; Wallace, R.B.; Cummings, S.R.; Black, D.M.; Bauer, D.C.; et al. Effects of antiresorptive therapies on glucose metabolism: Results from the FIT, HORIZON-PFT, and FREEDOM trials. J. Bone Miner. Res. 2013, 28, 1348–1354. [Google Scholar] [CrossRef]
- Toulis, K.A.; Nirantharakumar, K.; Ryan, R.; Marshall, T.; Hemming, K. Bisphosphonates and glucose homeostasis: A population-based, retrospective cohort study. J. Clin. Endocrinol. Metab. 2015, 100, 1933–1940. [Google Scholar] [CrossRef]
- Vestergaard, P.; Rejnmark, L.; Mosekilde, L. Are antiresorptive drugs effective against fractures in patients with diabetes? Calcif. Tissue Int. 2011, 88, 209–214. [Google Scholar] [CrossRef]
- Xuan, Y.; Sun, L.H.; Liu, D.M.; Zhao, L.; Tao, B.; Wang, W.Q.; Zhao, H.Y.; Liu, J.M.; Ning, G. Positive association between serum levels of bone resorption marker CTX and HbA1c in women with normal glucose tolerance. J. Clin. Endocrinol. Metab. 2015, 100, 274–281. [Google Scholar] [CrossRef]
- Yang, S.; Leslie, W.D.; Morin, S.N.; Majumdar, S.R.; Lix, L.M. Antiresorptive therapy and newly diagnosed diabetes in women: A historical cohort study. Diabetes Obes. Metab. 2016, 18, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Urano, T.; Shiraki, M.; Kuroda, T.; Tanaka, S.; Urano, F.; Uenishi, K.; Inoue, S. Low serum osteocalcin concentration is associated with incident type 2 diabetes mellitus in Japanese women. J. Bone Miner. Metab 2018, 36, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Oury, F.; Ferron, M.; Huizhen, W.; Confavreux, C.; Xu, L.; Lacombe, J.; Srinivas, P.; Chamouni, A.; Lugani, F.; Lejeune, H.; et al. Osteocalcin regulates murine and human fertility through a pancreas-bone-testis axis. J. Clin. Investig. 2013, 123, 2421–2433. [Google Scholar] [CrossRef] [PubMed]
- Hannemann, A.; Breer, S.; Wallaschofski, H.; Nauck, M.; Baumeister, S.E.; Barvencik, F.; Amling, M.; Schinke, T.; Haring, R.; Keller, J. Osteocalcin is associated with testosterone in the general population and selected patients with bone disorders. Andrology 2013, 1, 469–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanazawa, I.; Tanaka, K.; Ogawa, N.; Yamauchi, M.; Yamaguchi, T.; Sugimoto, T. Undercarboxylated osteocalcin is positively associated with free testosterone in male patients with type 2 diabetes mellitus. Osteoporos. Int. 2013, 24, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Legrand, E.; Hedde, C.; Gallois, Y.; Degasne, I.; Boux de Casson, F.; Mathieu, E.; Basle, M.F.; Chappard, D.; Audran, M. Osteoporosis in men: A potential role for the sex hormone binding globulin. Bone 2001, 29, 90–95. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Yang, Y.; Wen, C.Y.; Rong, L.M. Serum Osteocalcin and Testosterone Concentrations in Adult Males with or without Primary Osteoporosis: A Meta-Analysis. Biomed. Res. Int 2017, 2017, 9892048. [Google Scholar] [CrossRef]
- Lui, L.Y.; Stone, K.; Cauley, J.A.; Hillier, T.; Yaffe, K. Bone loss predicts subsequent cognitive decline in older women: The study of osteoporotic fractures. J. Am. Geriatr. Soc. 2003, 51, 38–43. [Google Scholar] [CrossRef]
- Rothman, M.S.; Arciniegas, D.B.; Filley, C.M.; Wierman, M.E. The neuroendocrine effects of traumatic brain injury. J. Neuropsychiatry Clin. Neurosci. 2007, 19, 363–372. [Google Scholar] [CrossRef]
- Tan, Z.S.; Seshadri, S.; Beiser, A.; Zhang, Y.; Felson, D.; Hannan, M.T.; Au, R.; Wolf, P.A.; Kiel, D.P. Bone mineral density and the risk of Alzheimer disease. Arch. Neurol. 2005, 62, 107–111. [Google Scholar] [CrossRef]
- Sohrabi, H.R.; Bates, K.A.; Weinborn, M.; Bucks, R.S.; Rainey-Smith, S.R.; Rodrigues, M.A.; Bird, S.M.; Brown, B.M.; Beilby, J.; Howard, M.; et al. Bone mineral density, adiposity, and cognitive functions. Front. Aging Neurosci. 2015, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Deng, J.; Zhang, M.; Zhou, H.D.; Wang, Y.J. Association between bone mineral density and the risk of Alzheimer’s disease. J. Alzheimers Dis. 2011, 24, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Puig, J.; Blasco, G.; Daunis-i-Estadella, J.; Moreno, M.; Molina, X.; Alberich-Bayarri, A.; Xifra, G.; Pedraza, S.; Ricart, W.; Fernandez-Aranda, F.; et al. Lower serum osteocalcin concentrations are associated with brain microstructural changes and worse cognitive performance. Clin. Endocrinol. (Oxf) 2016, 84, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Bradburn, S.; McPhee, J.S.; Bagley, L.; Sipila, S.; Stenroth, L.; Narici, M.V.; Paasuke, M.; Gapeyeva, H.; Osborne, G.; Sassano, L.; et al. Association between osteocalcin and cognitive performance in healthy older adults. Age Ageing 2016, 45, 844–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirckx, N.; Moorer, M.C.; Clemens, T.L.; Riddle, R.C. The role of osteoblasts in energy homeostasis. Nature Rev. Endocrinol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Merle, B.; Delmas, P.D. Normal carboxylation of circulating osteocalcin (bone Gla-protein) in Paget’s disease of bone. Bone Miner. 1990, 11, 237–245. [Google Scholar] [CrossRef]
- Poser, J.W.; Price, P.A. A method for decarboxylation of gamma-carboxyglutamic acid in proteins. Properties of the decarboxylated gamma-carboxyglutamic acid protein from calf bone. J. Biol. Chem. 1979, 254, 431–436. [Google Scholar]
- Heshmati, H.M.; Riggs, B.L.; Burritt, M.F.; McAlister, C.A.; Wollan, P.C.; Khosla, S. Effects of the circadian variation in serum cortisol on markers of bone turnover and calcium homeostasis in normal postmenopausal women. J. Clin. Endocrinol. Metab. 1998, 83, 751–756. [Google Scholar] [CrossRef]
- Gong, S.; Miao, Y.L.; Jiao, G.Z.; Sun, M.J.; Li, H.; Lin, J.; Luo, M.J.; Tan, J.H. Dynamics and correlation of serum cortisol and corticosterone under different physiological or stressful conditions in mice. PLoS ONE 2015, 10, e0117503. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Osteopetrosis | Genetic Transmission | Gene Mutation | Protein |
|---|---|---|---|
| ARO | Autosomal recessive osteopetrosis | TCIRG1 | α3 subunit V-H+ATPase |
| CLCN7 | Chloride channel 7 | ||
| OSTM1 | Osteopetrosis associated transmembrane protein | ||
| PLEKHM1 | Pleckstrin homology domain containing family M, member I | ||
| SNX10 | Sorting nexin 10 | ||
| TNFSF11 | Receptor activator for nuclear factor κB ligand | ||
| TNFRSF11A | Receptor activator for nuclear factor κB | ||
| IRO | Autosomal recessive osteopetrosis | CAII | Carbonic anhydrase |
| ADO | Autosomal dominant osteopetrosis | CLCN7 | Chloride channel 7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, M.; Battafarano, G.; Pepe, J.; Minisola, S.; Del Fattore, A. The Endocrine Function of Osteocalcin Regulated by Bone Resorption: A Lesson from Reduced and Increased Bone Mass Diseases. Int. J. Mol. Sci. 2019, 20, 4502. https://doi.org/10.3390/ijms20184502
Rossi M, Battafarano G, Pepe J, Minisola S, Del Fattore A. The Endocrine Function of Osteocalcin Regulated by Bone Resorption: A Lesson from Reduced and Increased Bone Mass Diseases. International Journal of Molecular Sciences. 2019; 20(18):4502. https://doi.org/10.3390/ijms20184502
Chicago/Turabian StyleRossi, Michela, Giulia Battafarano, Jessica Pepe, Salvatore Minisola, and Andrea Del Fattore. 2019. "The Endocrine Function of Osteocalcin Regulated by Bone Resorption: A Lesson from Reduced and Increased Bone Mass Diseases" International Journal of Molecular Sciences 20, no. 18: 4502. https://doi.org/10.3390/ijms20184502
APA StyleRossi, M., Battafarano, G., Pepe, J., Minisola, S., & Del Fattore, A. (2019). The Endocrine Function of Osteocalcin Regulated by Bone Resorption: A Lesson from Reduced and Increased Bone Mass Diseases. International Journal of Molecular Sciences, 20(18), 4502. https://doi.org/10.3390/ijms20184502