MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Influence on Metal Exclusion/Extrusion Mechanisms and Photosynthesis


 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
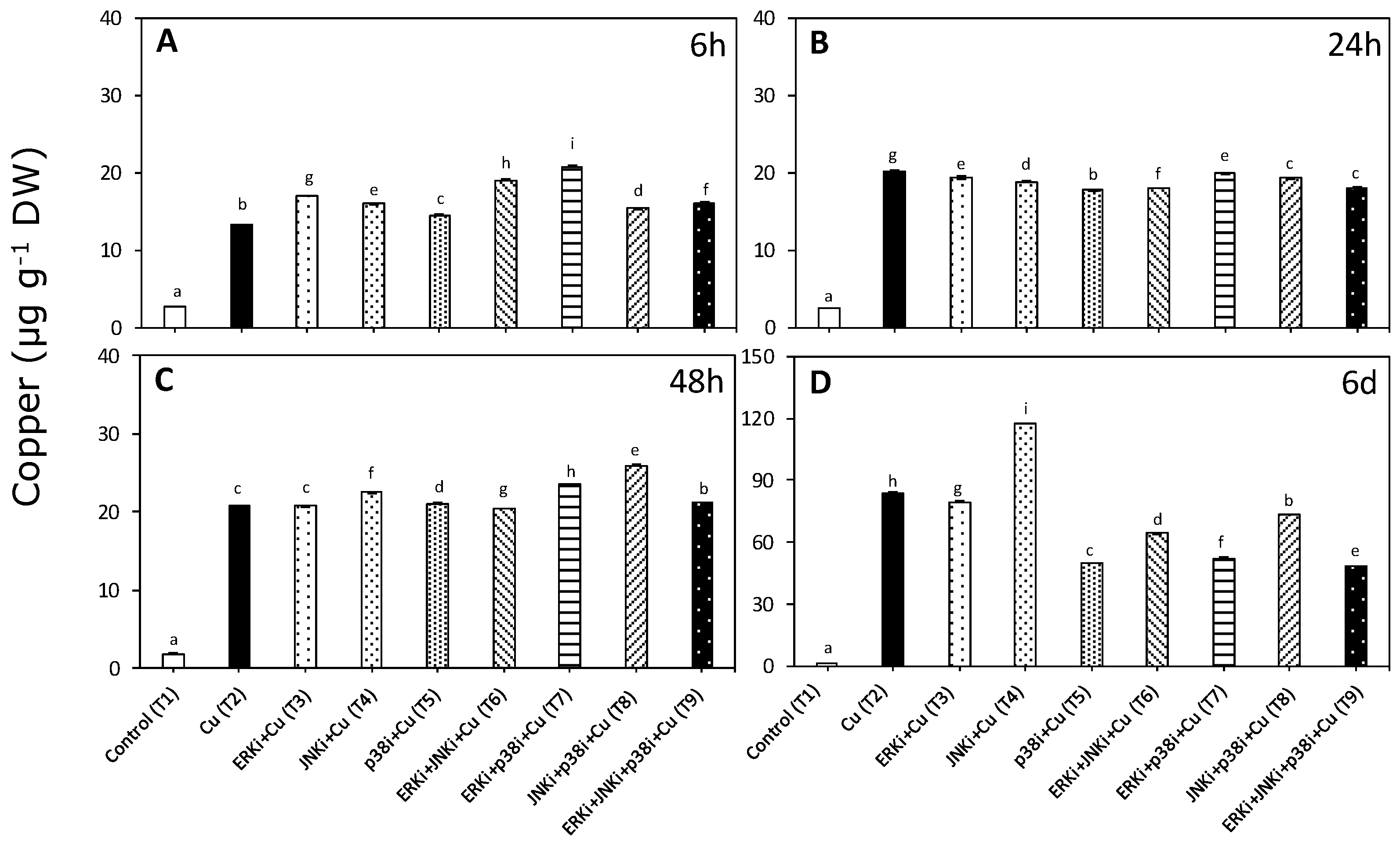
2.1. Copper Accumulation
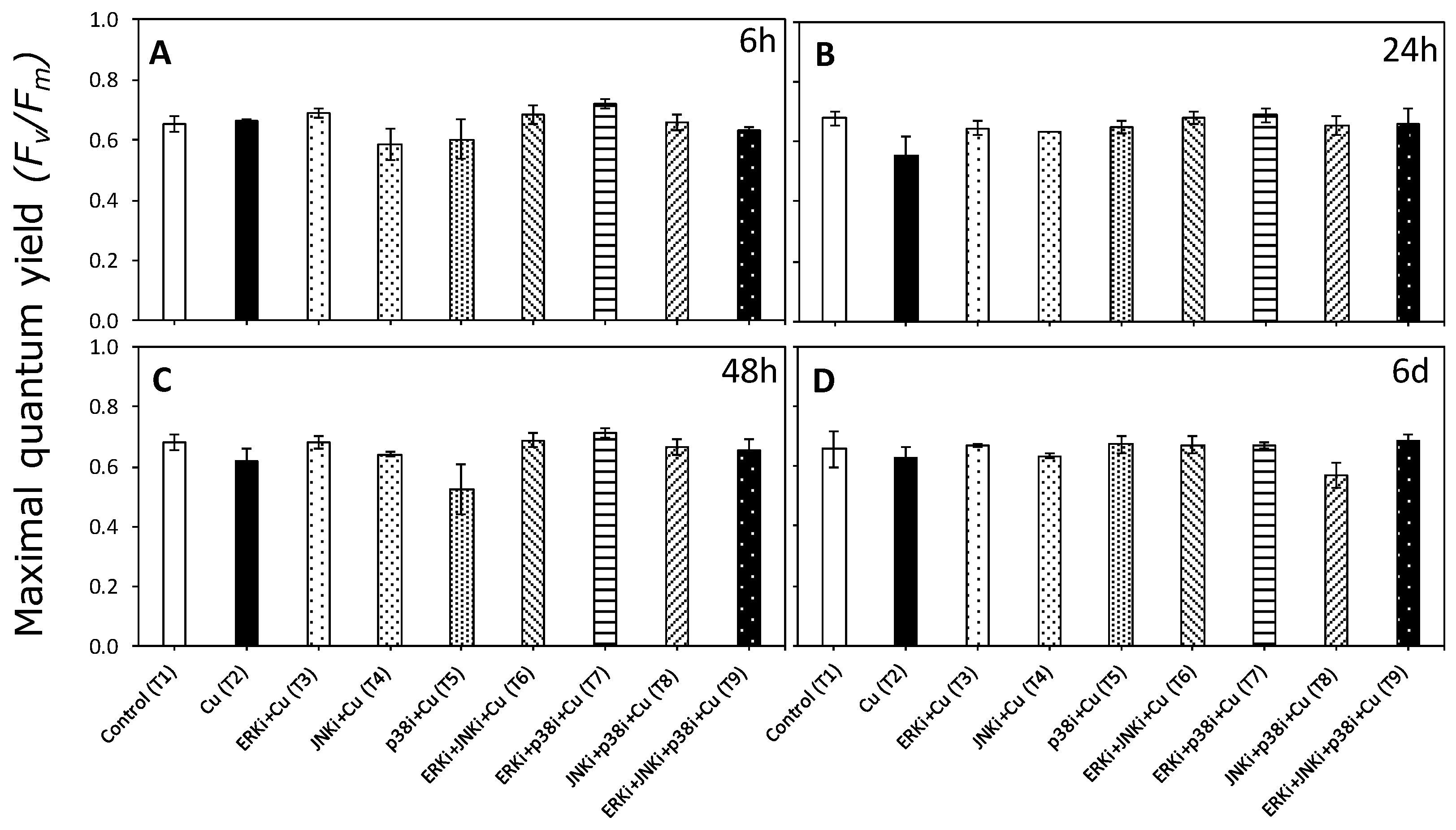
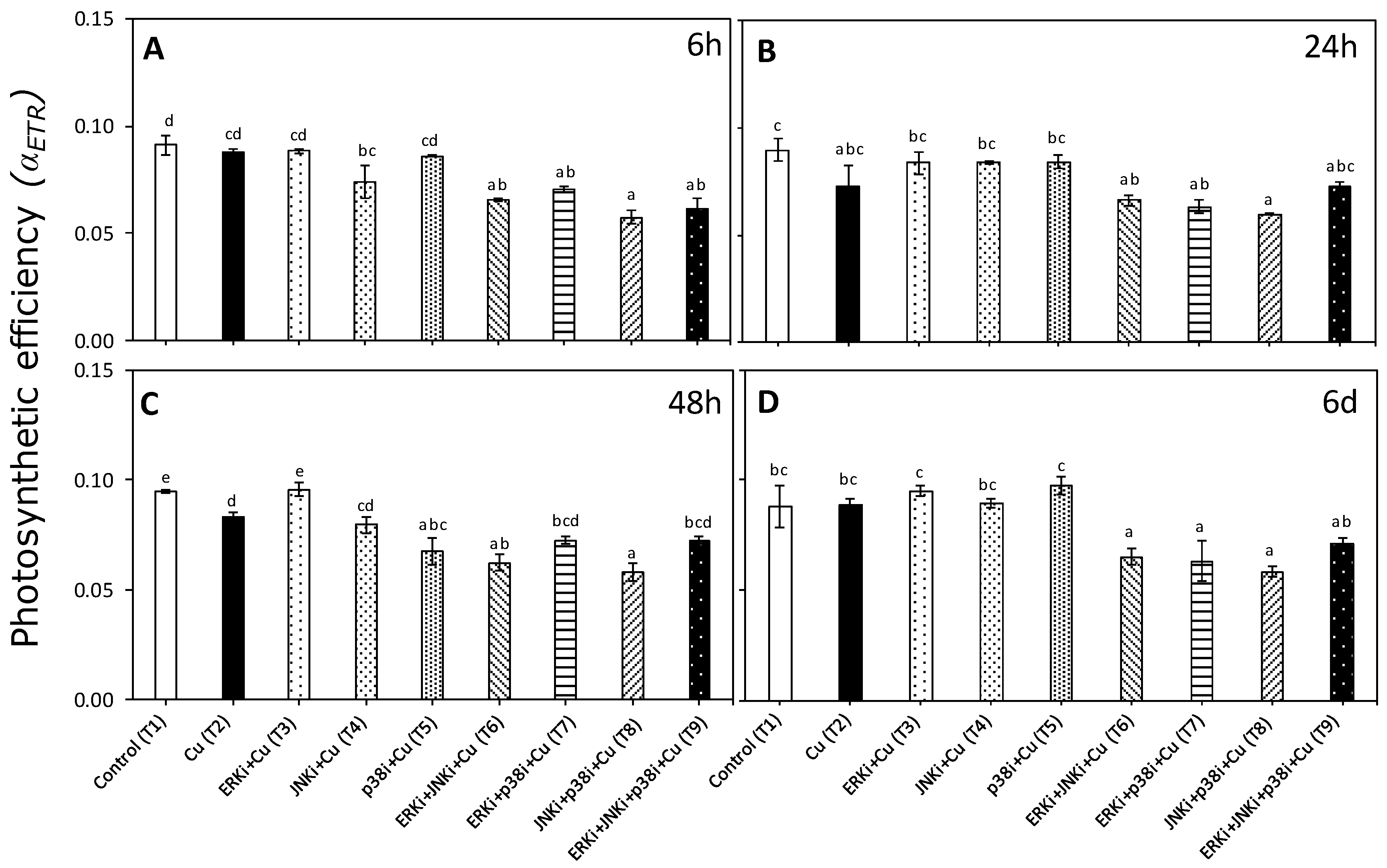
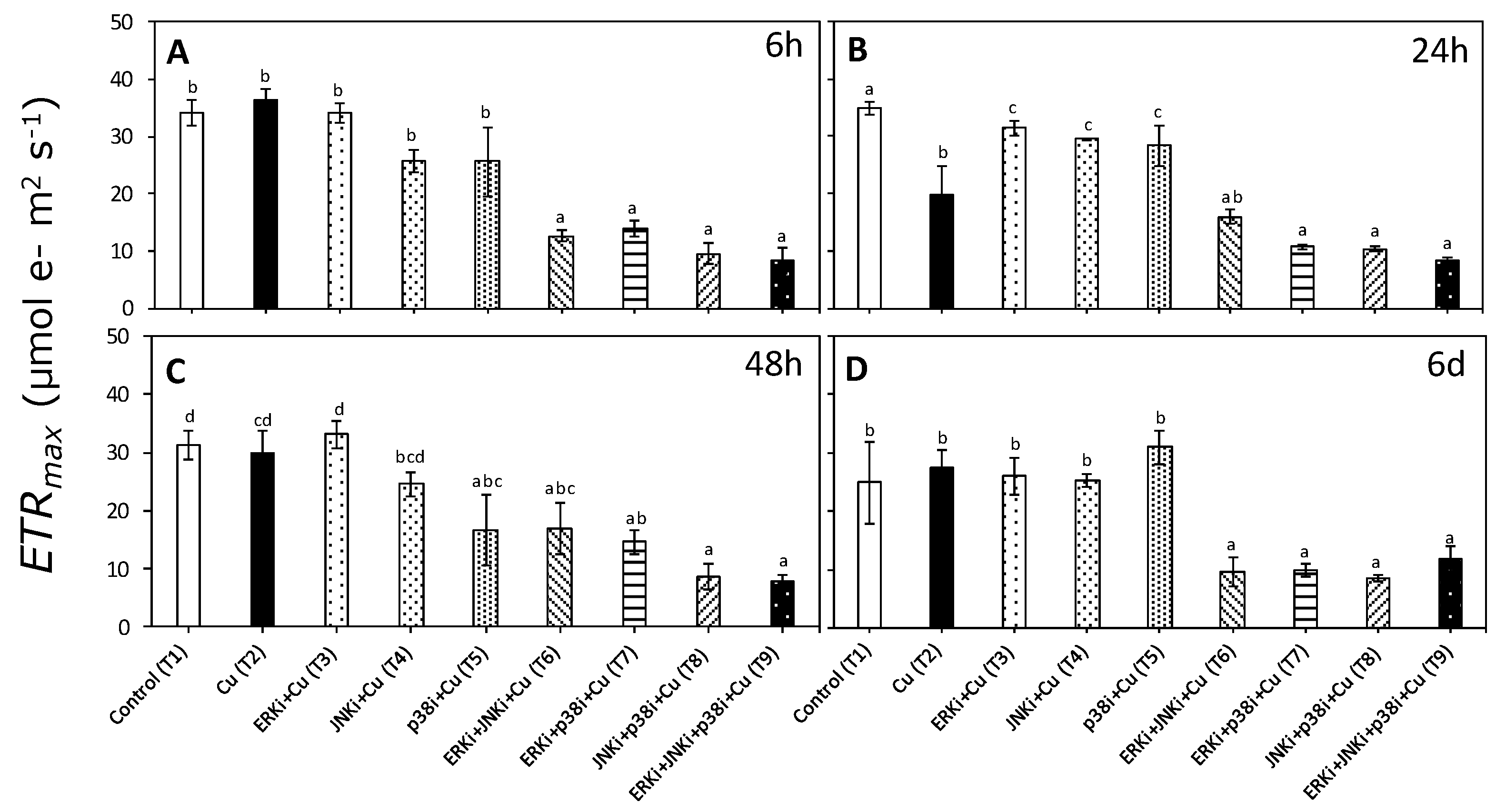
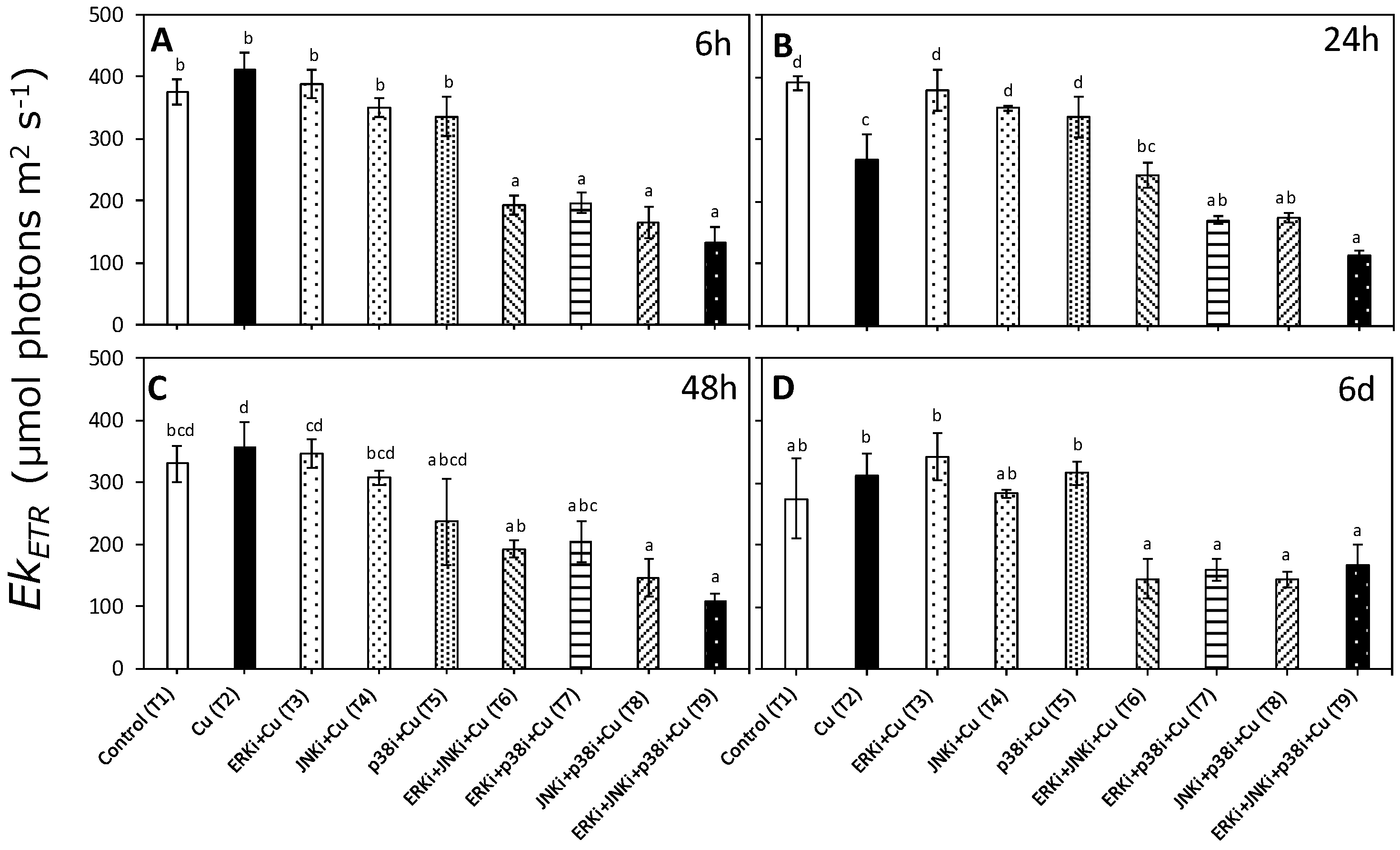
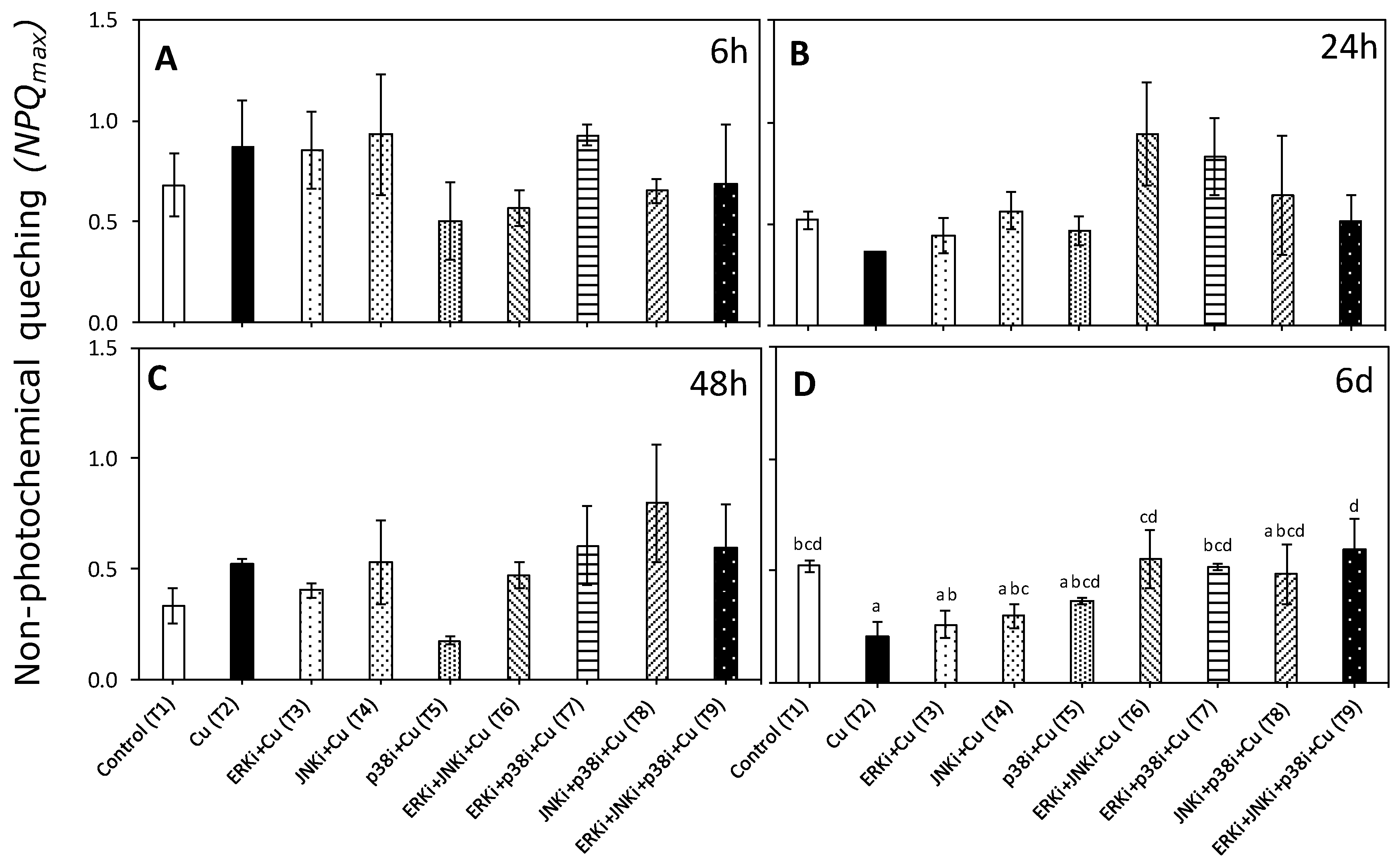
2.2. Photosynthetic Activity According to In Vivo Chlorophyll a Florescence
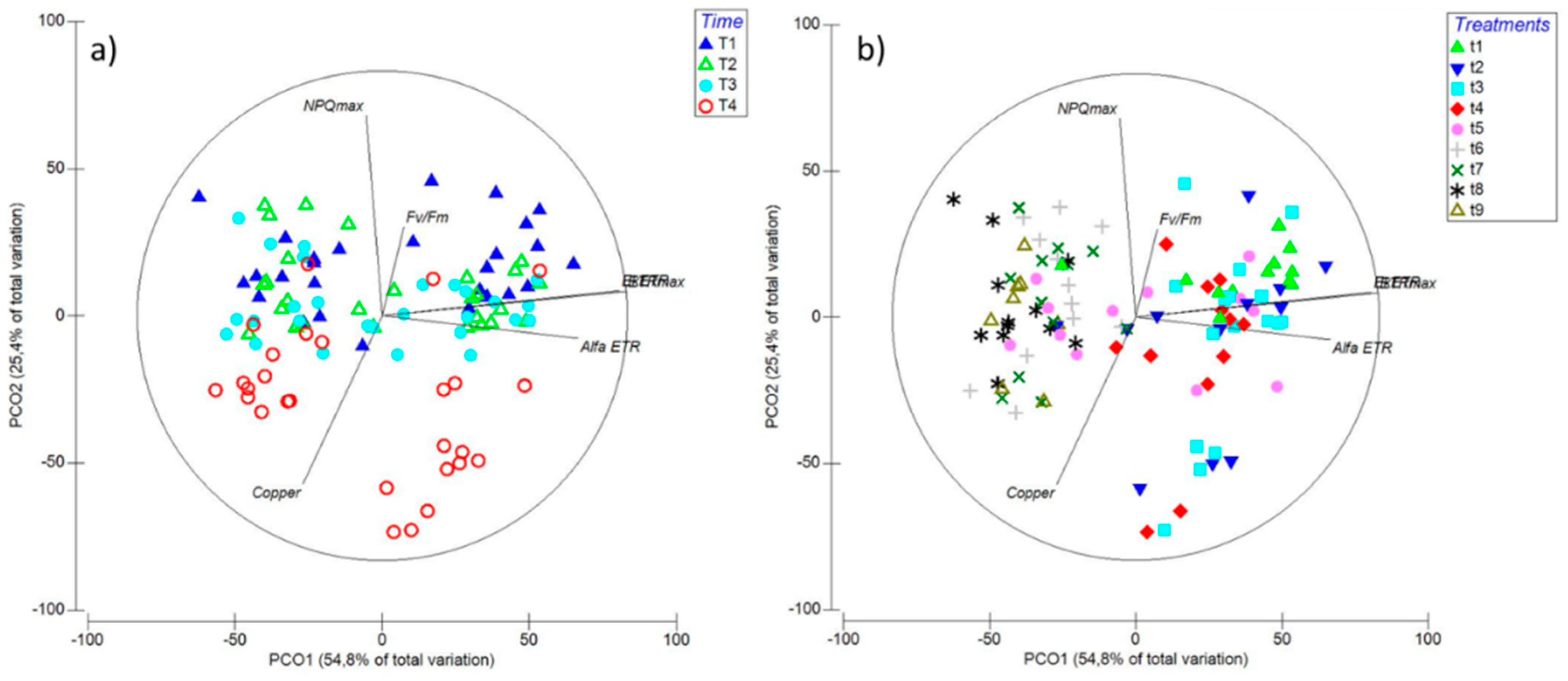
2.3. Principal Component Ordination (PCO) Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Culture Environment Conditions
4.2. Copper and MAPK Inhibitor Treatments
4.3. Intracellular Copper Accumulation
4.4. Photosynthesis and Energy Dissipation through In Vivo Chlorophyll a Fluorescence
4.5. Maximum Quantum Yield of PSII (Fv/Fm).
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| MAPK | mitogen activated protein kinases |
| ERK | Extracellular Signal Regulated Kinases |
| JNK | c-Jun N-terminal Kinases |
| p38 | Cytokinin Specific Binding Protein |
| ROS | reactive oxygen species |
| Fv/Fm | Maximal quantum yield of photosystem II |
| ETRmax | maximal electron transport rate of photosynthesis |
| EkETR | Saturation of the irradiance of electron transport rate |
| αETR | photosynthetic efficiency |
| NPQmax | maximal non-photochemical quenching |
References
- Rodríguez-Rojas, F.; Celis-Pla, P.S.M.; Méndez, L.; Moenne, F.; Muñoz, P.; Lobos, M.; Díaz, P.; Brown, M.; Moenne, A.; Sáez, C.A. MAPK pathway under chronic copper excess in green macroalgae (Chlorophyta): Involvement in the regulation of detoxification mechanisms. Int. J. Mol. Sci. 2019, in press. [Google Scholar]
- Song, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen-activated protein kinase signaling in plants under abiotic stress. Plant Signal. Behav. 2011, 6, 196–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzawa, A.; Ichijo, H. Redox control of cell fate by MAP kinase: Physiological roles of ASK1-MAP kinase pathway in stress signaling. Biochim. Et Biophys. Acta 2008, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Shi, X.; Jiang, B.H.; Fang, J. Insulin-like growth factor-I (IGF-I) induces epidermal growth factor receptor transactivation and cell proliferation through reactive oxygen species. Free Radic. Biol. Med. 2007, 42, 1651–1660. [Google Scholar] [CrossRef] [PubMed]
- Guyton, K.Z.; Liu, Y.; Gorospe, M.; Xu, Q.; Holbrook, N.J. Activation of mitogen-activated protein kinase by H2O2. Role in cell survival following oxidant injury. J. Biol. Chem. 1996, 271, 4138–4142. [Google Scholar] [CrossRef]
- Kamata, H.; Honda, S.; Maeda, S.; Chang, L.; Hirata, H.; Karin, M. Reactive oxygen species promote TNFalpha-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell 2005, 120, 649–661. [Google Scholar] [CrossRef]
- Jalmi, S.K.; Bhagat, P.K.; Verma, D.; Noryang, S.; Tayyeba, S.; Singh, K.; Sharma, D.; Sinha, A.K. Traversing the Links between Heavy Metal Stress and Plant Signaling. Front. Plant Sci. 2018, 9, 21. [Google Scholar] [CrossRef]
- Moenne, A.; González, A.; Sáez, C.A. Mechanisms of metal tolerance in marine macroalgae, with emphasis on copper tolerance in Chlorophyta and Rhodophyta. Aquat. Toxicol. 2016, 176, 30–37. [Google Scholar] [CrossRef]
- Kungolos, A.; Emmanouil, C.; Tsiridis, V.; Tsiropoulos, N. Evaluation of toxic and interactive toxic effects of three agrochemicals and copper using a battery of microbiotests. Sci. Total Environ. 2009, 407, 4610–4615. [Google Scholar] [CrossRef]
- Ferreira, J.G.; Andersen, J.H.; Borja, A.; Bricker, S.B.; Camp, J.; da Silva, M.C.; Garces, E.; Heiskanen, A.S.; Humborg, C.; Ignatiades, L.; et al. Overview of eutrophication indicators to assess environmental status within the European Marine Strategy Framework Directive. Estuar. Coast. Shelf Sci. 2011, 93, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Celis-Plá, P.S.M.; Brown, M.T.; Santillán-Sarmiento, A.; Korbee, N.; Sáez, C.A.; Figueroa, F.L. Ecophysiological and metabolic responses to interactive exposure to nutrients and copper excess in the brown macroalga Cystoseira tamariscifolia. Mar. Pollut. Bull. 2018, 128, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C.A.; González, A.; Contreras, R.A.; Moody, A.J.; Moenne, A.; Brown, M.T. A novel field transplantation technique reveals intra-specific metal-induced oxidative responses in strains of Ectocarpus siliculosus with different pollution histories. Environ. Pollut. 2015, 199, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C.A.; Pérez-Matus, A.; Lobos, M.G.; Oliva, D.; Vásquez, J.A.; Bravo, M. Environmental assessment in a shallow subtidal rocky habitat: Approach coupling chemical and ecological tools. Chem. Ecol. 2012, 28, 1–15. [Google Scholar] [CrossRef]
- Sheppard, C. Introduction to World Seas: An Environmental Evaluation. In World Seas: An Environmental Evaluation (Second Edition); Academic Press: New York, NY, USA, 2018. [Google Scholar]
- Ratkevicius, N.; Correa, J.A.; Moenne, A. Copper accumulation, synthesis of ascorbate and activation of ascorbate peroxidase in Enteromorpha compressa (L.) Grev. (Chlorophyta) from heavy metal-enriched environments in northern Chile. Plant Cell Environ. 2003, 26, 1599–1608. [Google Scholar] [CrossRef]
- Sáez, C.A.; Lobos, M.G.; Macaya, E.; Oliva, D.; Quiroz, W.; Brown, M.T. Variation in patterns of metal accumulation in thallus parts of Lessonia trabeculata (Laminariales; Phaeophyceae): Implications for biomonitoring. PLoS ONE 2012, 7, e50170. [Google Scholar] [CrossRef] [PubMed]
- Sáez, C.A.; Roncarati, F.; Moenne, A.; Moody, J.A.; Brown, M.T. Copper-induced intra-specific oxidative damage and antioxidant responses in strains of the brown alga Ectocarpus siliculosus with different pollution histories. Aquat. Toxicol. 2015, 159, 81–89. [Google Scholar] [CrossRef]
- Roncarati, F.; Sáez, C.A.; Greco, M.; Gledhill, M.; Bitonti, M.B.; Brown, M.T. Response differences between Ectocarpus siliculosus populations to copper stress involve cellular exclusion and induction of the phytochelatin biosynthetic pathway. Aquat. Toxicol. 2015, 159, 167–175. [Google Scholar] [CrossRef]
- Laporte, D.; Valdés, N.; González, A.; Sáez, C.A.; Zúñiga, A.; Navarrete, A.; Meneses, C.; Moenne, A. Copper-induced overexpression of genes encoding antioxidant system enzymes and metallothioneins involve the activation of CaMs, CDPKs and MEK1/2 in the marine alga Ulva compressa. Aquat. Toxicol. 2016, 177, 433–440. [Google Scholar] [CrossRef]
- Gómez, M.; González, A.; Sáez, C.A.; Moenne, A. Copper-induced membrane depolarizations involve the induction of mosaic TRP channels, which activate VDCC leading to calcium increases in Ulva compressa. Front. Plant Sci. 2016, 7, 754. [Google Scholar] [CrossRef]
- González, A.; Cabrera Mde, L.; Mellado, M.; Cabello, S.; Márquez, S.; Morales, B.; Moenne, A. Copper-induced intracellular calcium release requires extracellular calcium entry and activation of L-type voltage-dependent calcium channels in Ulva compressa. Plant Signal. Behav. 2012, 7, 728–732. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Cabrera Mde, L.; Henriquez, M.J.; Contreras, R.A.; Morales, B.; Moenne, A. Cross talk among calcium, hydrogen peroxide, and nitric oxide and activation of gene expression involving calmodulins and calcium-dependent protein kinases in Ulva compressa exposed to copper excess. Plant Physiol. 2012, 158, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.; González, A.; Sáez, C.A.; Morales, B.; Moenne, A. Copper-induced activation of TRP channels promotes extracellular calcium entry and activation of CaMs and CDPKs leading to copper entry and membrane depolarization in Ulva compressa. Front. Plant Sci. 2015, 6, 182. [Google Scholar] [PubMed]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Conde-Álvarez, R.; Gómez, I. Relations between electron transport rates determined by pulse amplitude modulated chlorophyll fluorescence and oxygen evolution in macroalgae under different light conditions. Photosynth. Res. 2003, 75, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Celis-Pla, P.S.M.; Korbee, N.; Gomez-Garreta, A.; Figueroa, F.L. Seasonal photoacclimation patterns in the intertidal macroalga Cystoseira tamariscifolia (Ochrophyta). Sci. Mar. 2014, 78, 377–388. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.-D., Caldwell, M.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1995; pp. 49–70. [Google Scholar]
- Celis-Plá, P.S.M.; Bouzon, Z.L.; Hall-Spencer, J.M.; Schmidt, E.C.; Korbee, N.; Figueroa, F.L. Seasonal biochemical and photophysiological responses in the intertidal macroalga Cystoseira tamariscifolia (Ochrophyta). Mar. Environ. Res. 2016, 115, 89–97. [Google Scholar] [CrossRef]
- Kumar, K.S.; Han, Y.-S.; Choo, K.-S.; Kong, J.-A.; Han, T. Chlorophyll fluorescence based copper toxicity assessment of two algal species. Toxicol. Environ. Health Sci. 2009, 1, 17–23. [Google Scholar] [CrossRef]
- Gao, G.; Liu, Y.; Li, X.; Feng, Z.; Xu, Z.; Wu, H.; Xu, J. Expected CO2-induced ocean acidification modulates copper toxicity in the green tide alga Ulva prolifera. Environ. Exp. Bot. 2017, 135, 63–72. [Google Scholar] [CrossRef]
- Kim, D.-Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef]
- Li, Y.; Iqbal, M.; Zhang, Q.; Spelt, C.; Bliek, M.; Hakvoort, H.W.J.; Quattrocchio, F.M.; Koes, R.; Schat, H. Two Silene vulgaris copper transporters residing in different cellular compartments confer copper hypertolerance by distinct mechanisms when expressed in Arabidopsis thaliana. New Phytol. 2017, 215, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.F.; Domingos, R.F.; Hauser, C.; Hutchins, C.M.; Zerges, W.; Wilkinson, K.J. Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. Appl. Environ. Microbiol. 2013, 79, 4774–4785. [Google Scholar] [CrossRef] [PubMed]
- Roelofs, D.; Aarts, M.G.M.; Schat, H.; Van Straalen, N.M. Functional ecological genomics to demonstrate general and specific responses to abiotic stress. Funct. Ecol. 2008, 22, 8–18. [Google Scholar] [CrossRef]
- Li, S.; Wang, W.; Gao, J.; Yin, K.; Wang, R.; Wang, C.; Petersen, M.; Mundy, J.; Qiu, J.-L. MYB75 Phosphorylation by MPK4 Is Required for Light-Induced Anthocyanin Accumulation in Arabidopsis. Plant Cell 2016, 28, 2866–2883. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, C.; Sheng, H.; Wang, Y.; Zeng, J.; Kang, H.; Fan, X.; Sha, L.; Zhang, H.; Zhou, Y. Transcriptome-wide identification and expression analyses of ABC transporters in dwarf polish wheat under metal stresses. Biol. Plant. 2017, 61, 293–304. [Google Scholar] [CrossRef]
- Doan, T.K.P.; Park, K.S.; Kim, H.K.; Park, D.S.; Kim, J.H.; Yoon, T.R. Inhibition of JNK and ERK pathways by SP600125- and U0126-enhanced osteogenic differentiation of bone marrow stromal cells. Tissue Eng. Regen. Med. 2012, 9, 283–294. [Google Scholar] [CrossRef]
- Ludwig, A.A.; Saitoh, H.; Felix, G.; Freymark, G.; Miersch, O.; Wasternack, C.; Boller, T.; Jones, J.D.G.; Romeis, T. Ethylene-mediated cross-talk between calcium-dependent protein kinase and MAPK signaling controls stress responses in plants. Proc. Natl. Acad. Sci. United States Am. 2005, 102, 10736–10741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehlmer, N.; Wurzinger, B.; Stael, S.; Hofmann-Rodrigues, D.; Csaszar, E.; Pfister, B.; Bayer, R.; Teige, M. The Ca2+-dependent protein kinase CPK3 is required for MAPK-independent salt-stress acclimation in Arabidopsis. Plant J. 2010, 63, 484–498. [Google Scholar] [CrossRef]
- Xu, H.-Y.; Zhang, C.; Li, Z.-C.; Wang, Z.-R.; Jiang, X.-X.; Shi, Y.-F.; Tian, S.-N.; Braun, E.; Mei, Y.; Qiu, W.-L.; et al. The MAPK Kinase Kinase GmMEKK1 regulates cell death and defense responses. Plant Physiol. 2018, 178, 907–922. [Google Scholar] [CrossRef]
- Aburai, N.; Abe, K. Metabolic switching: Synergistic induction of carotenogenesis in the aerial microalga, Vischeria helvetica, under environmental stress conditions by inhibitors of fatty acid biosynthesis. Biotechnol. Lett. 2015, 37, 1073–1080. [Google Scholar] [CrossRef]
- Gawroński, P.; Witoń, D.; Vashutina, K.; Bederska, M.; Betliński, B.; Rusaczonek, A.; Karpiński, S. Mitogen-activated protein kinase 4 is a salicylic acid-independent regulator of growth but not of photosynthesis in Arabidopsis. Mol. Plant 2014, 7, 1151–1166. [Google Scholar] [CrossRef] [PubMed]
- Gasulla, F.; Barreno, E.; Parages, M.L.; Cámara, J.; Jiménez, C.; Dörmann, P.; Bartels, D. The role of phospholipase D and MAPK signaling cascades in the adaption of lichen microalgae to desiccation: Changes in membrane lipids and phosphoproteome. Plant Cell Physiol. 2016, 57, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Silvan, J.M.; Reguero, M.; de Pascual-Teresa, S. A protective effect of anthocyanins and xanthophylls on UVB-induced damage in retinal pigment epithelial cells. Food Funct. 2016, 7, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzalka, K.; Prasad, M.N.V. Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Su, J.; Yang, L.; Zhu, Q.; Wu, H.; He, Y.; Liu, Y.; Xu, J.; Jiang, D.; Zhang, S. Active photosynthetic inhibition mediated by MPK3/MPK6 is critical to effector-triggered immunity. PLOS Biol. 2018, 16, e2004122. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Vera, J.; Castro, J.; Dennett, G.; Mellado, M.; Morales, B.; Correa, J.A.; Moenne, A. Co-occurring increases of calcium and organellar reactive oxygen species determine differential activation of antioxidant and defense enzymes in Ulva compressa (Chlorophyta) exposed to copper excess. Plant Cell Environ. 2010, 33, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Schiavon, F.; Betti, M.; Canesi, L.; Ballarin, L. Insight on signal transduction pathways involved in phagocytosis in the colonial ascidian Botryllus schlosseri. J. Invertebr. Pathol. 2013, 112, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.; Heyland, A. Thyroid hormones accelerate initiation of skeletogenesis via mapk (ERK1/2) in larval sea urchins (Strongylocentrotus purpuratus). Front. Endocrinol. 2018, 9, 16. [Google Scholar] [CrossRef]
- Zhao, H.Y.; Liu, W.; Wang, Y.; Dai, N.N.; Gu, J.H.; Yuan, Y.; Liu, X.Z.; Bian, J.C.; Liu, Z.P. Cadmium induces apoptosis in primary rat osteoblasts through caspase and mitogen-activated protein kinase pathways. J. Vet. Sci. 2015, 16, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.L.; Wang, K.L.; Chen, P.L.; Li, Y. The c-Jun N-terminal kinases (JNK)/mitogen-activated Protein Kinase (MAPK) is responsible for the protection of tanshinol (Danshensu) upon H2O2-induced L-6 rat myoblast cell injury. Acta Sci. Vet. 2014, 42, 8. [Google Scholar]
- Khokon, M.A.R.; Salam, M.A.; Jammes, F.; Ye, W.; Hossain, M.A.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Kwak, J.M.; Murata, Y. Two guard cell mitogen-activated protein kinases, MPK9 and MPK12, function in methyl jasmonate-induced stomatal closure in Arabidopsis thaliana. Plant Biol. 2015, 17, 946–952. [Google Scholar] [CrossRef]
- Livanos, P.; Galatis, B.; Gaitanaki, C.; Apostolakos, P. Phosphorylation of a p38-like MAPK is involved in sensing cellular redox state and drives atypical tubulin polymer assembly in angiosperms. Plant Cell Environ. 2014, 37, 1130–1143. [Google Scholar] [CrossRef]
- de Oliveira, E.A.G.; Romeiro, N.C.; Ribeiro, E.D.; Santa-Catarina, C.; Oliveira, A.E.A.; Silveira, V.; de Souza, G.A.; Venancio, T.M.; Cruz, M.A.L. Structural and functional characterization of the protein kinase Mps1 in Arabidopsis thaliana. PLoS ONE 2012, 7, 9. [Google Scholar] [CrossRef]
- Charneco, G.O.; Parages, M.L.; Camarena-Gomez, T.; Jimenez, C. Phosphorylation of MAP Kinases crucially controls the response to environmental stress in Dunaliella viridis. Environ. Exp. Bot. 2018, 156, 203–213. [Google Scholar] [CrossRef]
- Jimenez, C.; Berl, T.; Rivard, C.J.; Edelstein, C.L.; Capasso, J.M. Phosphorylation of MAP kinase-like proteins mediate the response of the halotolerant alga Dunaliella viridis to hypertonic shock. Biochim. Et Biophys. Acta-Mol. Cell Res. 2004, 1644, 61–69. [Google Scholar] [CrossRef]
- Parages, M.L.; Capasso, J.M.; Niell, F.X.; Jiménez, C. Responses of cyclic phosphorylation of MAPK-like proteins in intertidal macroalgae after environmental stress. J. Plant Physiol. 2014, 171, 276–284. [Google Scholar] [CrossRef]
- Parages, M.L.; Figueroa, F.L.; Conde-Alvarez, R.M.; Jimenez, C. Phosphorylation of MAPK-like proteins in three intertidal macroalgae under stress conditions. Aquat. Biol. 2014, 22, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, C.; Cossio, B.R.; Rivard, C.J.; Berl, T.; Capasso, J.M. Cell division in the unicellular microalga Dunaliella viridis depends on phosphorylation of extracellular signal-regulated kinases (ERKs). J. Exp. Bot. 2007, 58, 1001–1011. [Google Scholar] [CrossRef]
- Eilers, P.H.C.; Peeters, J.C.H. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol. Model. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- GOWER, J.C. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, K. PERMANOVA+ for Primer: Guide to Software and Statistical Methods; PRIMER-e: Plymouth, UK, 2008; p. 218. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Celis-Plá, P.S.M.; Rodríguez-Rojas, F.; Méndez, L.; Moenne, F.; Muñoz, P.T.; Lobos, M.G.; Díaz, P.; Sánchez-Lizaso, J.L.; Brown, M.T.; Moenne, A.; et al. MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Influence on Metal Exclusion/Extrusion Mechanisms and Photosynthesis. Int. J. Mol. Sci. 2019, 20, 4547. https://doi.org/10.3390/ijms20184547
Celis-Plá PSM, Rodríguez-Rojas F, Méndez L, Moenne F, Muñoz PT, Lobos MG, Díaz P, Sánchez-Lizaso JL, Brown MT, Moenne A, et al. MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Influence on Metal Exclusion/Extrusion Mechanisms and Photosynthesis. International Journal of Molecular Sciences. 2019; 20(18):4547. https://doi.org/10.3390/ijms20184547
Chicago/Turabian StyleCelis-Plá, Paula S. M., Fernanda Rodríguez-Rojas, Lorena Méndez, Fabiola Moenne, Pamela T. Muñoz, M. Gabriela Lobos, Patricia Díaz, José Luis Sánchez-Lizaso, Murray T. Brown, Alejandra Moenne, and et al. 2019. "MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Influence on Metal Exclusion/Extrusion Mechanisms and Photosynthesis" International Journal of Molecular Sciences 20, no. 18: 4547. https://doi.org/10.3390/ijms20184547
APA StyleCelis-Plá, P. S. M., Rodríguez-Rojas, F., Méndez, L., Moenne, F., Muñoz, P. T., Lobos, M. G., Díaz, P., Sánchez-Lizaso, J. L., Brown, M. T., Moenne, A., & Sáez, C. A. (2019). MAPK Pathway under Chronic Copper Excess in Green Macroalgae (Chlorophyta): Influence on Metal Exclusion/Extrusion Mechanisms and Photosynthesis. International Journal of Molecular Sciences, 20(18), 4547. https://doi.org/10.3390/ijms20184547