The Artificial Promoter rMdAG2I Confers Flower-specific Activity in Malus




Abstract
:1. Introduction
2. Results and Discussion
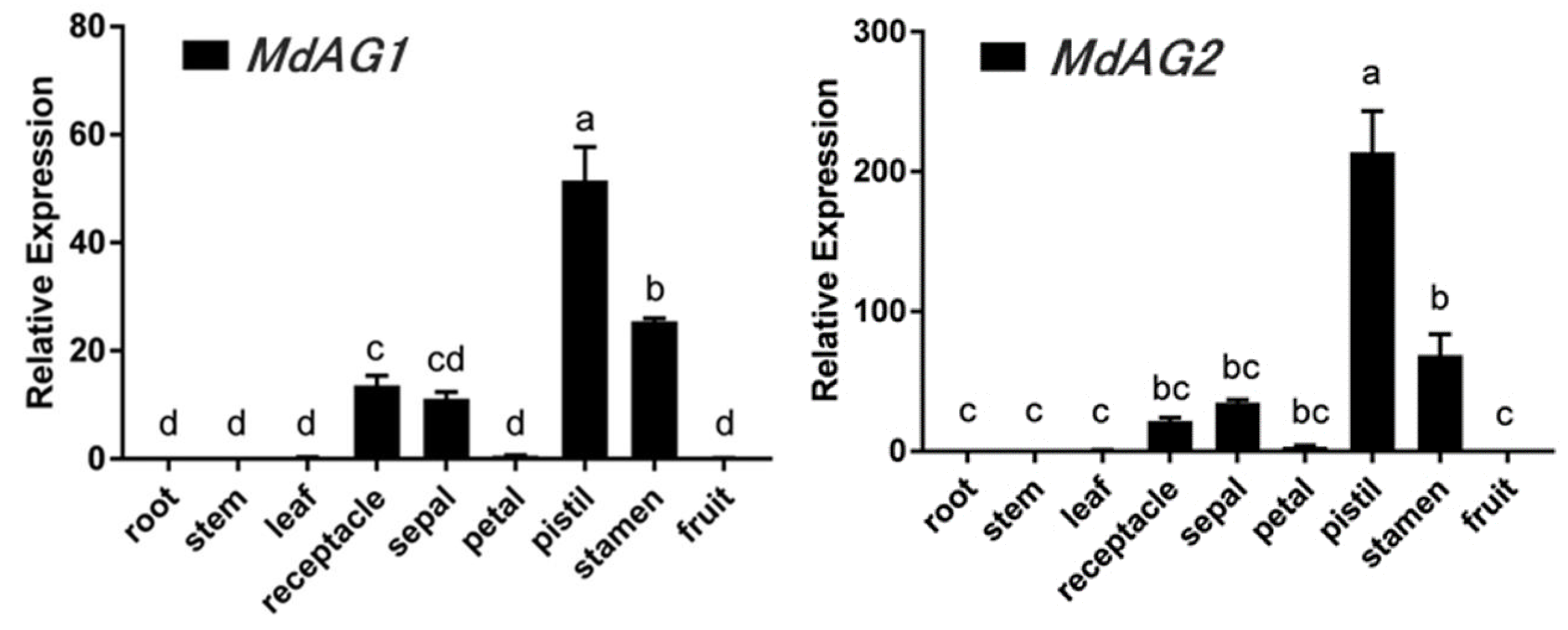
2.1. Isolation and Expression Analysis of MdAG1 and MdAG2
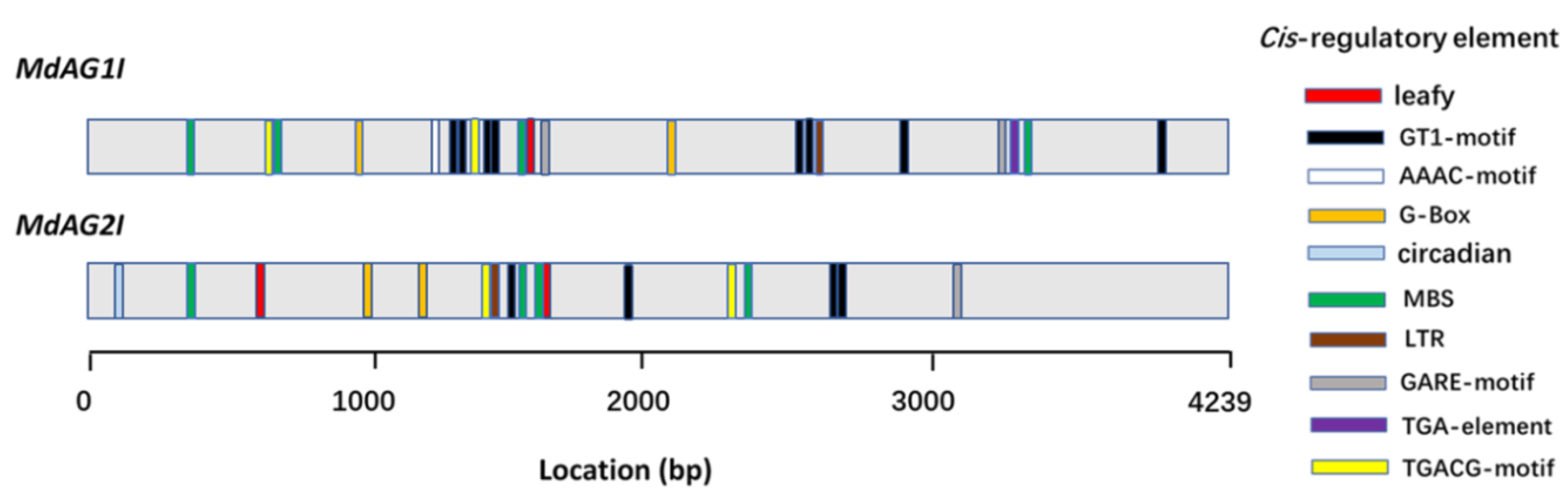
2.2. Cloning and Analysis of the Second Introns of MdAG1 and MdAG2
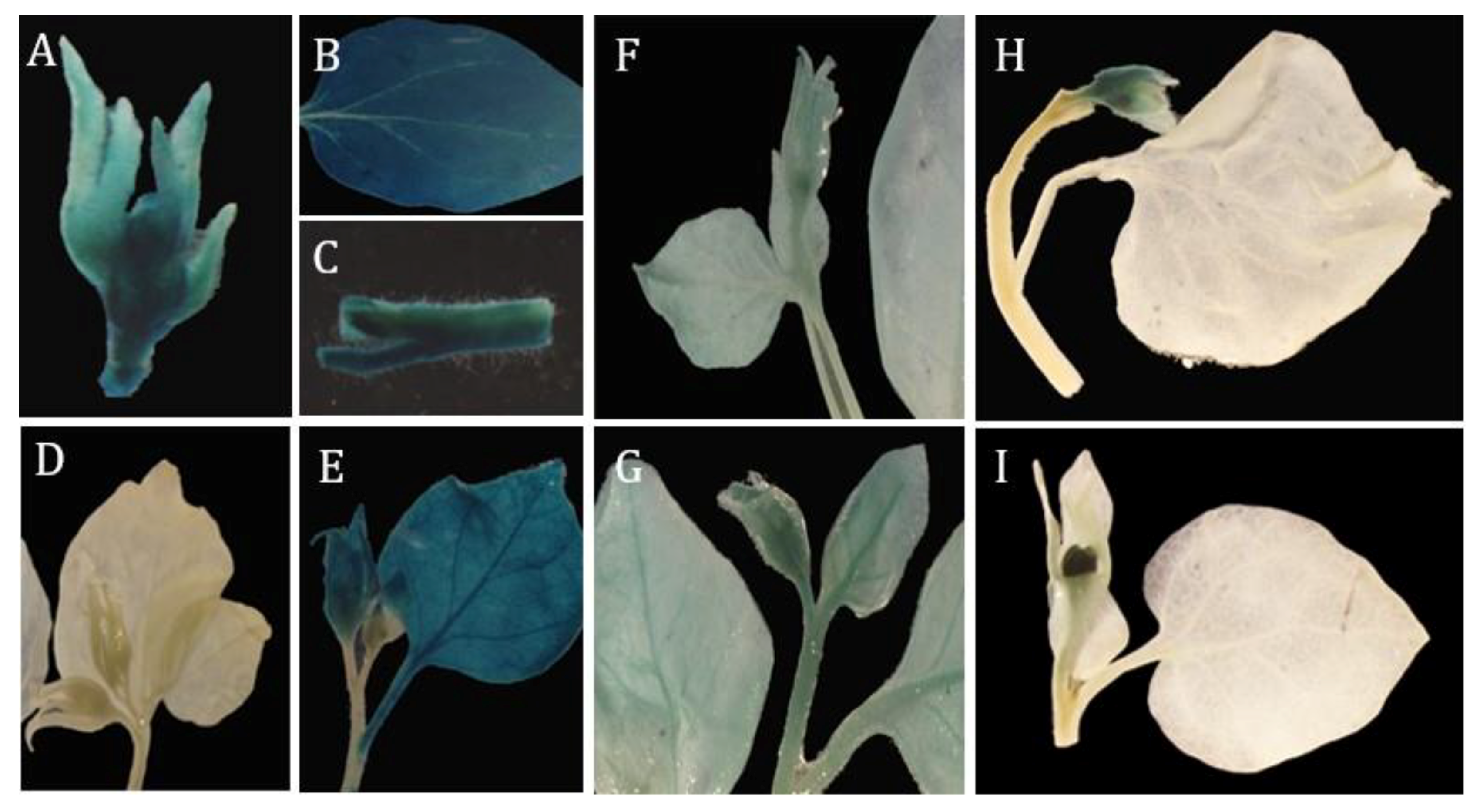
2.3. Floral-Specific Expression of rMdAG2I::GUS in Tobacco
2.4. Optimization of Agrobacterium-Mediated Transient Transformation Conditions for Malus Flowers
2.5. rMdAG2I::GUS Confers Flower-Specific Expression in Malus
3. Materials and Methods
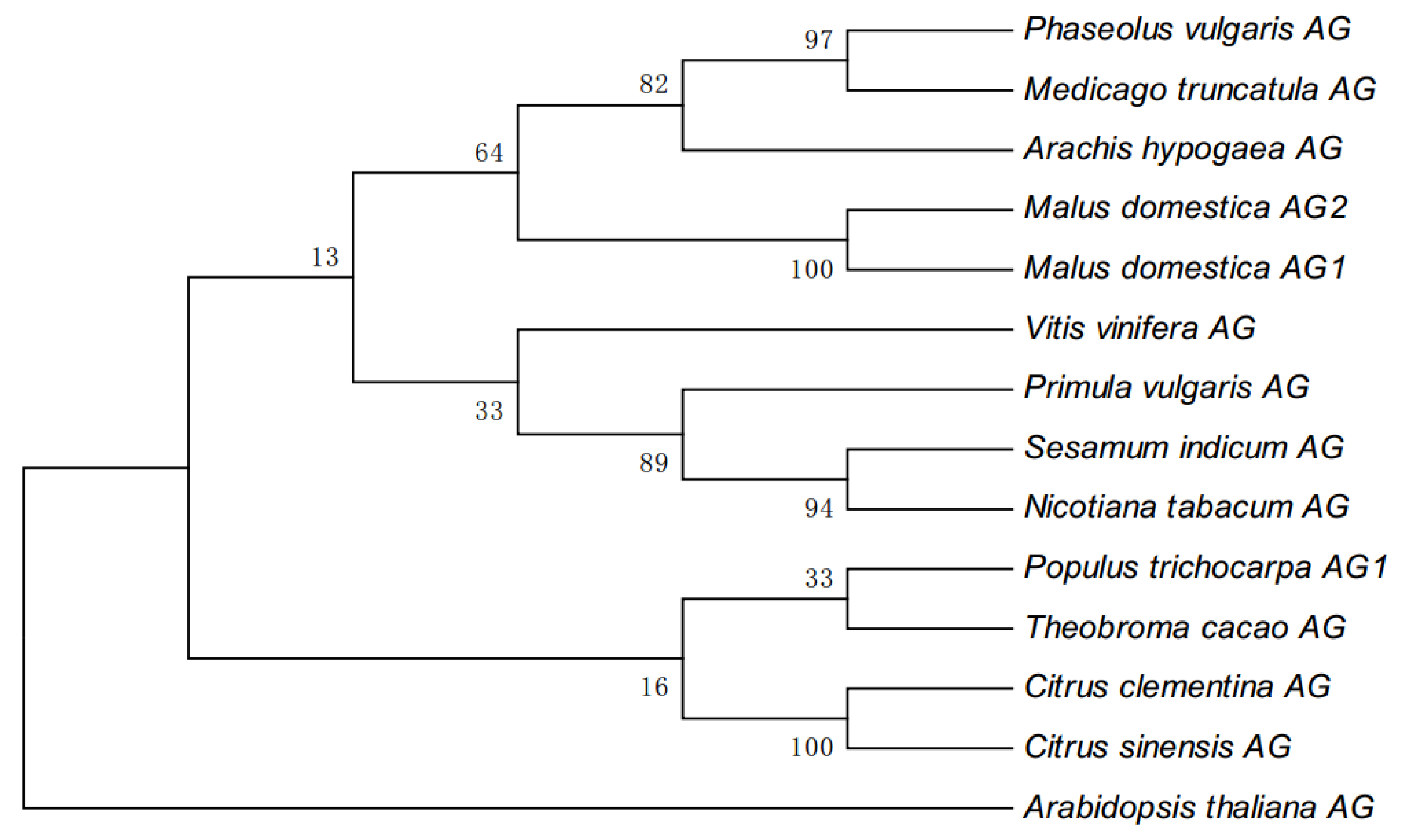
3.1. Phylogenetic Analysis
3.2. RNA Extraction and qRT-PCR
3.3. Cloning of MdAG1 and MdAG2 Second-Intron Sequences
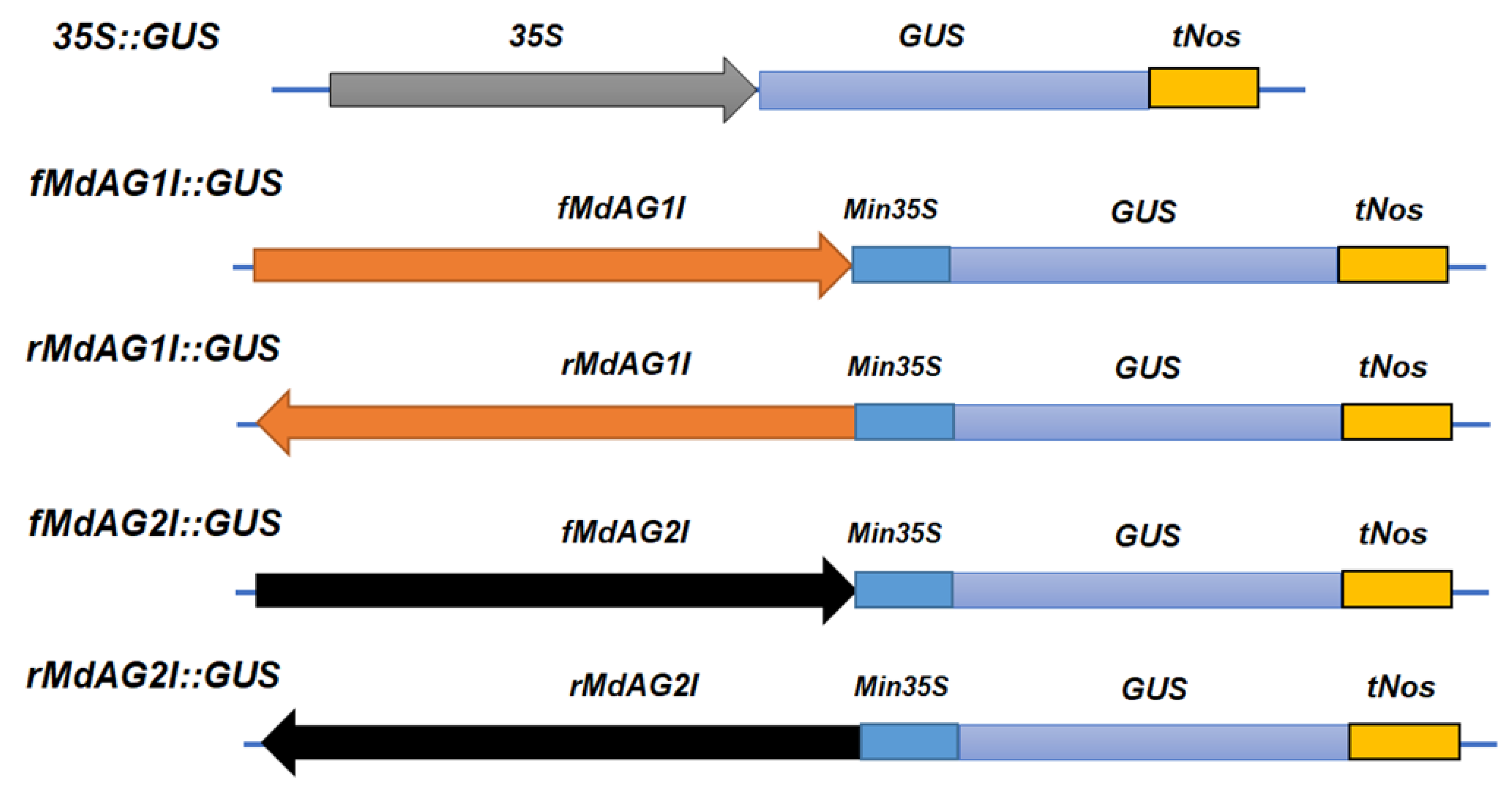
3.4. Plasmid Construction
3.5. Stable Transformation of Tobacco
3.6. Vacuum Infiltration of Malus Micromalus Flowers
3.7. Particle Bombardment of ‘Golden Delicious’ Apple Flowers
3.8. GUS Staining Assays
Author Contributions
Funding
Conflicts of Interest
References
- Phipps, J.B.; Robertso, K.R.; Smith, P.G.; Rohrer, J.R. A checklist of the subfamily Maloideae (Rosaceae). Can. J. Bot. 1988, 68, 2209–2269. [Google Scholar] [CrossRef]
- Lisandru, T.T.; Adrian FÜstÖs, A.; Adelina DumitraŞ, A.; Viorel Mitre, V. Flower development of ornamental Crabapple according to BBCH scale. Bulletin UASVM Hort. 2017, 74, 147–148. [Google Scholar] [CrossRef]
- Del Duca, S.; Aloisi, I.; Parrotta, L.; Cai, G. Cytoskeleton, Transglutaminase and gametophytic self-incompatibility in the Malinae (Rosaceae). Int. J. Mol. Sci. 2019, 20, 209. [Google Scholar] [CrossRef] [PubMed]
- Broothaerts, W.; Janssens, G.A.; Proost, P.; Broekaert, W.F. cDNA cloning and molecular analysis of two self-incompatibility alleles from apple. Plant Mol. Biol. 1995, 27, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Mir, J.; Ahmed, N.; Singh, D.B.; Sheemar, G.; Hamid, A.; Zaffer, S.; Shafi, W. Molecular identification of S-alleles associated with self-incompatibility in apple (Malus spp.) genotypes. Indian J. Agric. Sci. 2016, 86, 78–81. [Google Scholar]
- Garratt, M.P.; Breeze, T.D.; Boreux, V.; Fountain, M.T.; McKerchar, M.; Webber, S.M.; Coston, D.J.; Jenner, N.; Dean, R.; Westbury, D.B.; et al. Apple pollination: Demand depends on variety and supply depends on pollinator identity. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Sapir, G.; Baras, Z.; Azmon, G.; Goldway, M.; Shafir, S.; Allouche, A.; Stern, E.; Stern, R.A. Synergistic effects between bumblebees and honey bees in apple orchards increase cross pollination, seed number and fruit size. Sci. Hortic. 2017, 219, 107–117. [Google Scholar] [CrossRef]
- Russo, L.; Park, M.G.; Blitzer, E.J.; Danforth, B.N. Flower handling behavior and abundance determine the relative contribution of pollinators to seed set in apple orchards. Agric. Ecosyst. Environ. 2017, 246, 102–108. [Google Scholar] [CrossRef]
- Robbie, F.A.; Atkinson, C.J. Wood and tree age as factors influencing the ability of apple flowers to set fruit. J. Hortoc. Sci. 1994, 69, 609–623. [Google Scholar] [CrossRef]
- Kelsey, D.F.; Brown, S.K. ‘McIntosh Wijcik’: A columnar mutation of ‘McIntosh’ apple proving useful in physiology and breeding research. J. Am. Pomol. Soc. 1992, 46, 83–87. [Google Scholar]
- Fisher, D.V. The ‘Wijcik Spur McIntosh’. Fruit Varieties J. 1995, 49, 212–213. [Google Scholar]
- Kim, D.H.; Park, S.; Lee, J.Y.; Ha, S.H.; Lim, S.H. Enhancing flower color through simultaneous expression of the B-peru and mPAP1 transcription factors under control of a flower-specific promoter. Int. J. Mol. Sci. 2019, 19, 309. [Google Scholar] [CrossRef] [PubMed]
- Verdonk, J.C.; Shibuya, K.; Loucas, H.M.; Colquhoun, T.A.; Underwood, B.A.; Clark, D.G. Flower-specific expression of the Agrobacterium tumefaciens isopentenyltransferase gene results in radial expansion of floral organs in Petunia hybrida. Plant Biotechnol. J. 2008, 6, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Guo, X.; Wang, Q.; Wang, Y.; Zhao, D.; Yao, L.; Wang, S.; Liu, G.; Li, T. Overexpression of MsDREB6.2 results in cytokinin-deficient developmental phenotypes and enhances drought tolerance in transgenic apple plants. Plant J. 2017, 89, 510–526. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hagen, G.; Guilfoyle, T.J. Altered morphology in transgenic tobacco plants that overproduce cytokinins in specific tissues and organs. Dev. Biol. 1992, 153, 386–395. [Google Scholar] [CrossRef]
- Werner, T.; Holst, K.; Pors, Y.; Guivarc’h, A.; Mustroph, A.; Chriqui, D.; Grimm, B.; Schmülling, T. Cytokinin deficiency causes distinct changes of sink and source parameters in tobacco shoots and roots. J. Exp. Bot. 2008, 59, 2659–2672. [Google Scholar] [CrossRef] [Green Version]
- Yanofsky, M.F.; Ma, H.; Bowman, J.L.; Drews, G.N.; Feldmann, K.A.; Meyerowitz, E.M. The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors. Nature 1990, 346, 35–39. [Google Scholar] [CrossRef]
- Drews, G.N.; Bowman, J.L.; Meyerowitz, E.M. Negative regulation of the Arabidopsis homeotic gene AGAMOUS by the APETALA2 product. Cell 1991, 65, 991–1002. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, Z. The second intron of AGAMOUS drives carpel- and stamen-specific expression sufficient to induce complete sterility in Arabidopsis. Plant Cell Rep. 2008, 27, 855–863. [Google Scholar] [CrossRef]
- Yang, Y.; Singer, S.D.; Liu, Z. Two similar but distinct second intron fragments from tobacco AGAMOUS homologous confer identical floral organ-specific expression sufficient for generating complete sterility in plants. Planta 2010, 231, 1159–1169. [Google Scholar] [CrossRef]
- Li, W.; Hu, W.; Feng, C.; Chen, L.; Zhuang, W.; Katin-Grazzini, L.; McAvoy, R.J.; Guillard, K.; Li, Y. An AGAMOUS intron-driven cytotoxin leads to flowerless tobacco and produces no detrimental effects on vegetative growth of either tobacco or poplar. Plant Biotechnol. J. 2016, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, C.G.; Vosman, B.; Smulders, M.J.M. Cloning and characterization of four apple MADS box genes isolated from vegetative tissue. J. Exp. Bot. 2002, 53, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Klocko, A.L.; Borejsza-Wysocka, E.; Brunner, A.M.; Shevchenko, O.; Aldwinckle, H.; Strauss, S.H. Transgenic suppression of AGAMOUS genes in apple reduces fertility and increases floral attractiveness. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Z.; Hu, B.; Chen, G.P.; Shi, N.N.; Zhao, Y.; Yin, Q.C.; Liu, J.J. Application of Arabidopsis AGAMOUS second intron for the engineered ablation of flower development in transgenic tobacco. Plant Cell Rep. 2008, 27, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Drews, G.N.; Meyerowitz, E.M. Expression of the Arabidopsis floral homeotic gene AGAMOUS is restricted to specific cell types late in flower development. Plant Cell 1991, 3, 749–758. [Google Scholar] [PubMed]
- Ito, T.; Ng, K.H.; Lim, T.S.; Yu, H.; Meyerowitz, E.M. The Homeotic Protein AGAMOUS Controls Late Stamen Development by Regulating a Jasmonate Biosynthetic Gene in Arabidopsis. Plant Cell 2007, 19, 3516–3529. [Google Scholar] [CrossRef] [PubMed]
- Eccher, G.; Ferrero, S.; Populin, F.; Colombo, L.; Botton, A. Apple (Malus domestica L. Borkh) as an emerging model for fruit development. Plant Biosyst. 2014, 148, 157–168. [Google Scholar] [CrossRef]
- Klee, H.J.; Giovannoni, J.J. Genetics and Control of Tomato Fruit Ripening and Quality Attributes. Annu. Rev. Genet. 2011, 45, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van, D.P.Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic. Acids. Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Busch, M.A.; Bomblies, K.; Weigel, D. Activation of a floral homeotic gene in Arabidopsis. Science 1999, 285, 585–587. [Google Scholar] [CrossRef]
- Hong, R.; Hamaguchi, L.; Busch, M.A.; Weigel, D. Regulatory Elements of the Floral Homeotic Gene AGAMOUS Identified by Phylogenetic Footprinting and Shadowing. Plant Cell. 2003, 15, 1296–1309. [Google Scholar] [CrossRef] [PubMed]
- Maizel, A.; Busch, M.A.; Tanahashi, T.; Perkovic, J.; Kato, M.; Mitsuyasu Hasebe, M.; Weigel, D. The floral regulator LEAFY evolves by substitutions in the DNA binding domain. Science 2005, 308, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, W.; Zhao, G.; Fan, X.; Long, H.; Fan, Y.; Shi, M.; Tan, X.; Zhang, L. New insights of salicylic acid into stamen abortion of female flowers in tung tree (Vernicia fordii). Front. Genet. 2019, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yang, D.; Sun, J.; Peng, H.; Yin, C.; Guo, R.; Ning, G.; Hu, H. A novel PhLRR gene promoter is sufficient for engineering male sterility in petunia. Plant Mol. Biol. Rep. 2016, 34, 970. [Google Scholar] [CrossRef]
- Tang, L.; Chu, H.; Yip, W.K.; Yeung, E.C.; Lo, C. An anther-specific dihydroflavonol 4-reductase-like gene (DRL1) is essential for male fertility in Arabidopsis. New Phytol. 2009, 181, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Schwinn, K.E.; Bennett, M.J.; Hunter, D.A.; Waugh, T.L.; Pathirana, N.N.; Brummell, D.A.; Jameson, P.E.; Davies, E.M. Methods for transient assay of gene function in floral tissues. Plant Methods 2007, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wang, J.; Zhang, D.; Wang, Y. A gene regulatory network controlled by BpERF2 and BpMYB102 in Birch under drought conditions. Int. J. Mol. Sci. 2019, 20, 3071. [Google Scholar] [CrossRef]
- Chevreau, E.; Dousset, N.; Joffrion, C.; Richer, A.; Charrier, A.; Vergne, E. Agroinfiltration is a key factor to improve the efficiency of apple and pear transformation. Sci. Hortic. 2019, 251, 150–154. [Google Scholar] [CrossRef]
- Lin, Q.; Li, Z.; Zhang, L.; Tan, X.; Long, H.; Wu, L. High-efficiency regeneration of seedlings from hypocotyl explants of tung tree (Vernicia fordii). Int. J. Agric. Biol. 2016, 18, 370–376. [Google Scholar] [CrossRef]
- Tsuda, K.; Qi, Y.; Nguyen, V.L.; Bethke, G.; Tsuda, Y.; Glazebrook, J.; Katagiri, F. An efficient Agrobacterium-mediated transient transformation of Arabidopsis. Plant J. 2012, 69, 713–719. [Google Scholar] [CrossRef]
- Hwang, H.; Yu, M.; Lai, E. Agrobacterium-mediated plant transformation: Biology and applications. Arabidopsis Book 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Q.; Yang, T.; Wu, Y.; Wang, G.; Yang, F.; Wang, R.; Lin, X.; Li, G. Development of Agrobacterium-mediated transient expression system in Caragana intermedia and characterization of CiDREB1C in stress response. BMC Plant Biol. 2019, 19, 237. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.F.; Li, H.; Qin, R.Y.; Wang, L.; Li, L.; Wei, P.C.; Yang, J.B. Gene targeting using the Agrobacterium tumefaciens-mediated CRISPR-Cas system in rice. Rice 2014, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.G.; Wang, N. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, C.; Hirai, N.; Komori, S.; Wada, M.; Okada, K.; Osakabe, K.; Yamamoto, T.; Osakabe, Y. Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep-UK 2016, 6, 31481. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.Y.; Hu, Y.N.; Lv, Y.D.; Rose, J.K.C.; Sun, Y.Q.; Shen, F.; Wang, Y.; Zhang, X.Z.; Xu, X.F.; Wu, T.; et al. Natural variation underlies differences in ETHYLENE RESPONSE FACTOR17 activity in fruit peel degreening. Plant Physiol. 2018, 176, 2292–2304. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.X.; Fu, D.Q.; Zhu, B.Z.; Liu, H.P.; Shen, X.Y.; Luo, Y.B. Sprout vacuum-infiltration: A simple and efficient agroinoculation method for virus-induced gene silencing in diverse solanaceous species. Plant Cell Rep. 2012, 31, 1713–1722. [Google Scholar] [CrossRef]
- Sapers, G.M.; Garzarella, L.; Pilizota, V. Application of browning inhibitors to cut apple and potato by vacuum and pressure infiltration. Food Sci. 1990, 55, 1049–1053. [Google Scholar] [CrossRef]
- Dai, S.; Zheng, P.; Marmey, P.; Zhang, S.; Tian, W.; Chen, S.; Beachy, R.N. Comparative analysis of transgenic rice plants obtained by Agrobacterium-mediated transformation and particle bombardment. Mol. Breeding 2001, 7, 25–33. [Google Scholar] [CrossRef]
- Azuma, M.; Morimoto, R.; Hirose, M.; Morita, Y.; Hoshino, A.; Iida, S.; Oshima, Y.; Mitsuda, N.; Takagi, M.O.; Shiratake, K. A petal-specific InMYB1 promoter from Japanese morning glory: A useful tool for molecular breeding of floricultural crops. Plant Biotechnol. J. 2016, 14, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Zapata, D.G.; Vargas, J.M.S.; Osorio, K.S.M.; Manjarrez, E.S.; Flores, J.L.C.; Salvador, M.D.C.O.; Figueroa, N.V.D.; Nogué, F.; Corona, A.B. Efficient eiting of the nuclear APT reporter gene in Chlamydomonas reinhardtii via expression of a CRISPR-Cas9 module. Int. J. Mol. Sci. 2019, 20, 1247. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 12, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2011, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Xu, X.; Li, X.; Hu, X.; Wu, T.; Wang, Y.; Xu, X.; Zhang, X.; Han, Z. High miR156 Expression Is Required for Auxin-Induced Adventitious Root Formation via MxSPL26 Independent of PINs and ARFs in Malus xiaojinensis. Front. Plant Sci. 2017, 19, 1059. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vectors | No. of Transgenic Lines | Plant Tissues | Floral Specificity | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Stem | Leaf | Sepal | Petal | Stamen | Carpel | ||||
| 35S::GUS | 8 | + | + | + | + | + | + | No | |
| fMdAG1I::GUS | 28 | + | + | + | + | + | + | No | |
| rMdAG11::GUS | 23 | + | + | + | + | + | + | ||
| fMdAG2I::GUS | 31 | + | + | + | + | + | + | ||
| rMdAG2I::GUS | Type I | 11 | − | − | + | + | + | + | Yes |
| Type II | 9 | − | − | − | − | + | + | ||
| Vacuum Time (min) | Soaking Time (min) | Condition of Treated Flowers |
| 2.0 | 1.0 | No damage, No staining |
| 2.0 | 1.5 | No damage, Blue staining |
| 2.0 | 2.0 | No damage, Blue staining |
| 2.0 | 3.0 | No damage, Blue staining |
| 3.0 | 1.0 | No damage, No staining |
| 3.0 | 1.5 | No damage, Blue staining |
| 3.0 | 2.0 | Slightly damaged |
| 3.0 | 3.0 | Damaged |
| 4.0 | 1.0 | Damaged |
| 4.0 | 1.5 | Damaged |
| 4.0 | 2.0 | Damaged |
| 4.0 | 3.0 | Damaged |
| Organs | Stem | Sepal | Petal | Stamen | Pistil | Receptacle | |
|---|---|---|---|---|---|---|---|
| Vectors | |||||||
| 35S::GUS | + | + | + | + | + | + | |
| fMdAG1I::GUS | + | + | + | + | + | + | |
| rMdAG1I::GUS | + | − | − | + | − | − | |
| fMdAG2I::GUS | + | + | + | + | + | + | |
| rMdAG2I::GUS | − | + | + | + | + | + | |
| Organ | Stem | Leaf | Sepal | Petal | Stamen | Pistil | Receptacle |
|---|---|---|---|---|---|---|---|
| 35S::GUS | + | + | + | + | + | + | + |
| rMdAG2I::GUS | − | − | + | + | + | + | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, H.; Liu, L.; Fan, X.; Asghar, S.; Li, Y.; Wang, Y.; Xu, X.; Wu, T.; Zhang, X.; Qiu, C.; et al. The Artificial Promoter rMdAG2I Confers Flower-specific Activity in Malus. Int. J. Mol. Sci. 2019, 20, 4551. https://doi.org/10.3390/ijms20184551
Dong H, Liu L, Fan X, Asghar S, Li Y, Wang Y, Xu X, Wu T, Zhang X, Qiu C, et al. The Artificial Promoter rMdAG2I Confers Flower-specific Activity in Malus. International Journal of Molecular Sciences. 2019; 20(18):4551. https://doi.org/10.3390/ijms20184551
Chicago/Turabian StyleDong, Haiqiang, Lanxin Liu, Xingqiang Fan, Sumeera Asghar, Yi Li, Yi Wang, Xuefeng Xu, Ting Wu, Xinzhong Zhang, Changpeng Qiu, and et al. 2019. "The Artificial Promoter rMdAG2I Confers Flower-specific Activity in Malus" International Journal of Molecular Sciences 20, no. 18: 4551. https://doi.org/10.3390/ijms20184551
APA StyleDong, H., Liu, L., Fan, X., Asghar, S., Li, Y., Wang, Y., Xu, X., Wu, T., Zhang, X., Qiu, C., Han, Z., & Li, W. (2019). The Artificial Promoter rMdAG2I Confers Flower-specific Activity in Malus. International Journal of Molecular Sciences, 20(18), 4551. https://doi.org/10.3390/ijms20184551