PTEN, A Target of Microrna-374b, Contributes to the Radiosensitivity of Canine Oral Melanoma Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
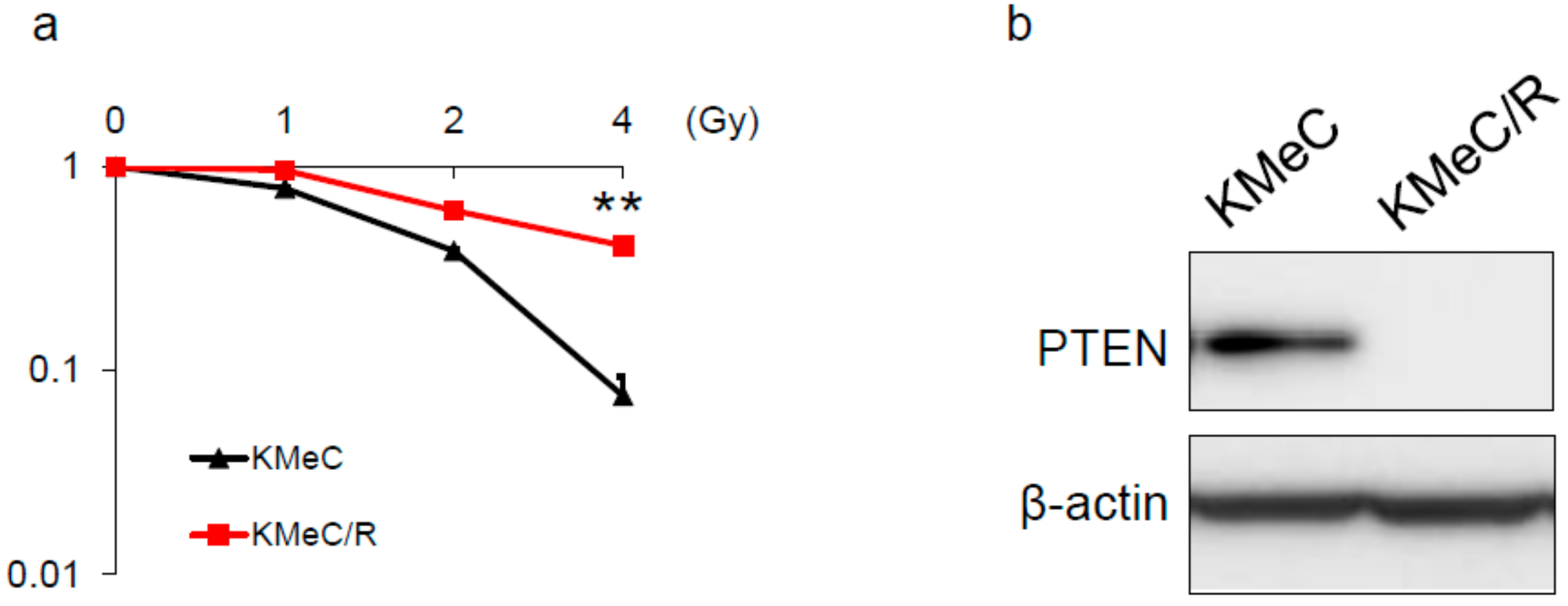
2.1. Development of Radioresistant CoMM Cells
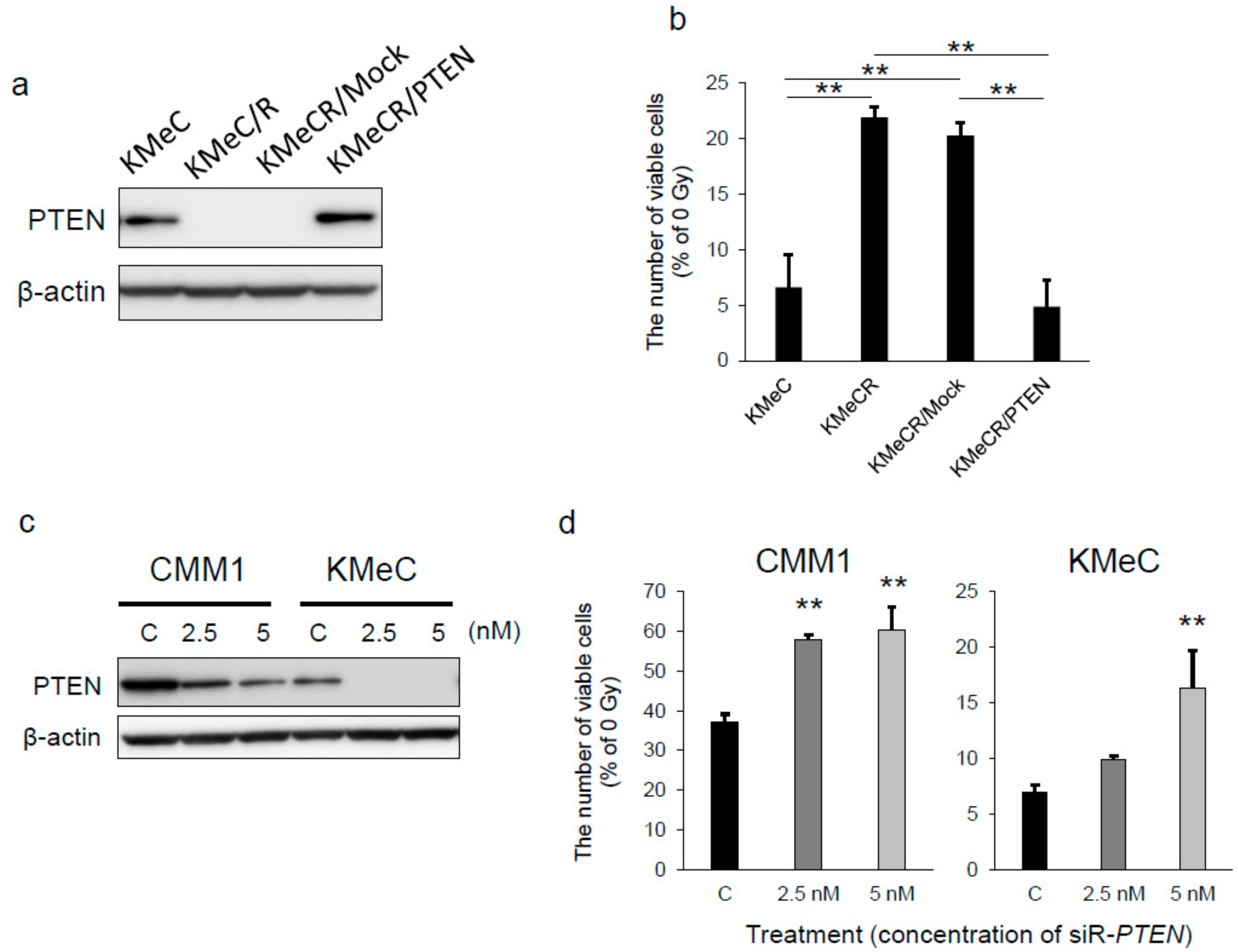
2.2. Overexpression of PTEN Canceled the Radioresistance of KMeC/R Cells and Silencing of PTEN Enhanced that of KMeC Cells
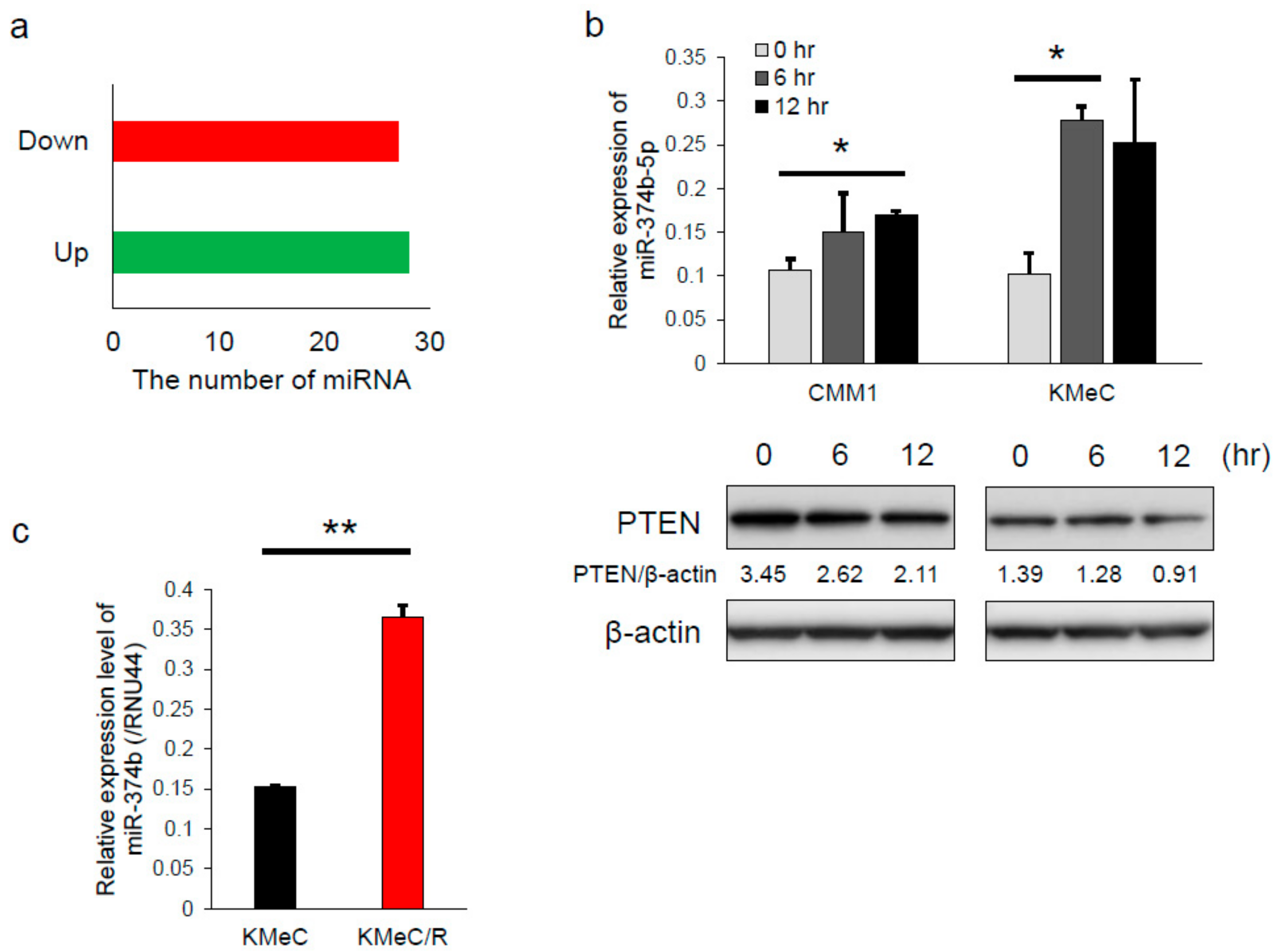
2.3. Differential Expression of miRNAs Associated with Irradiation
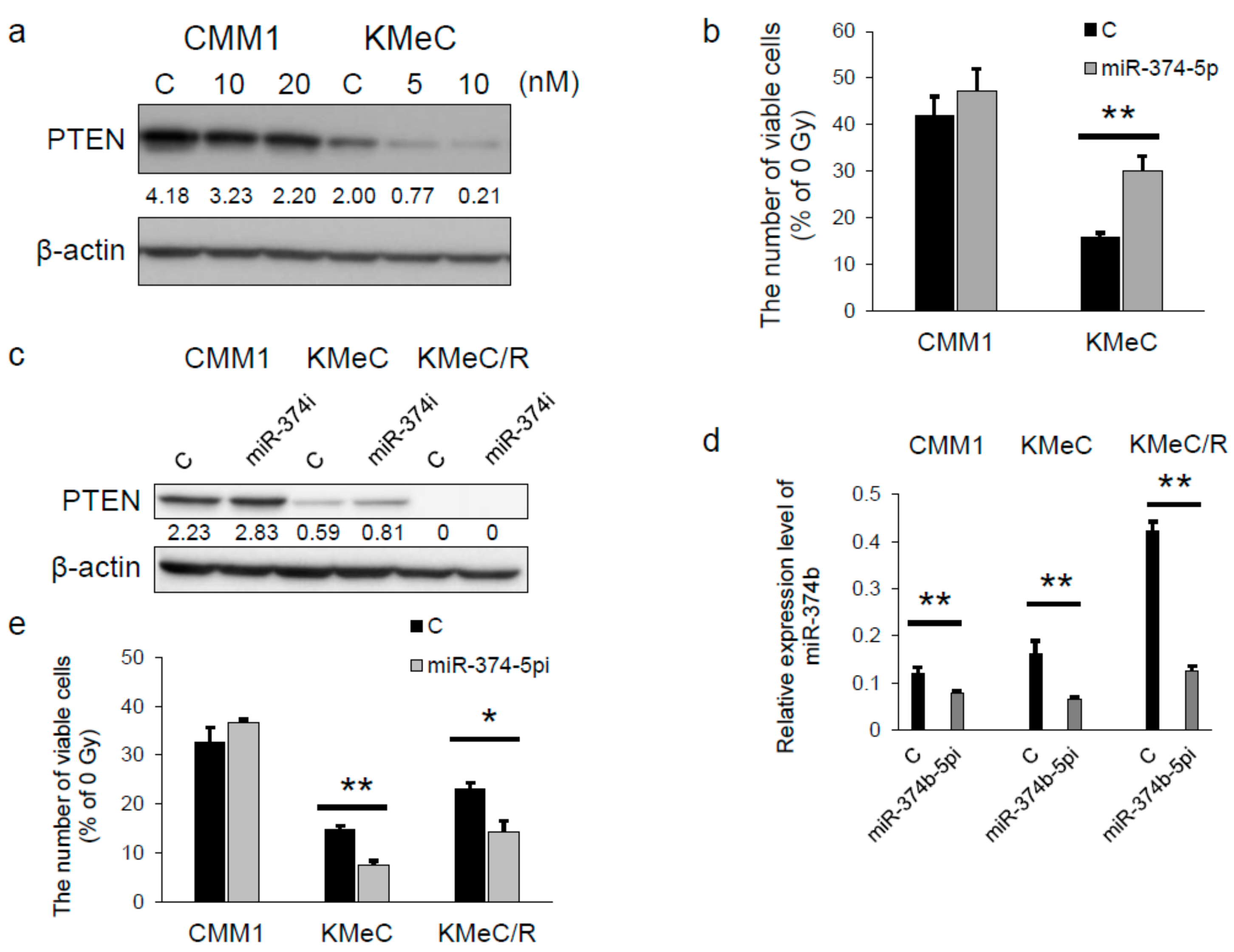
2.4. miR-374b-5p Reduced the Expression Level of PTEN and Conferred the Radioresistance
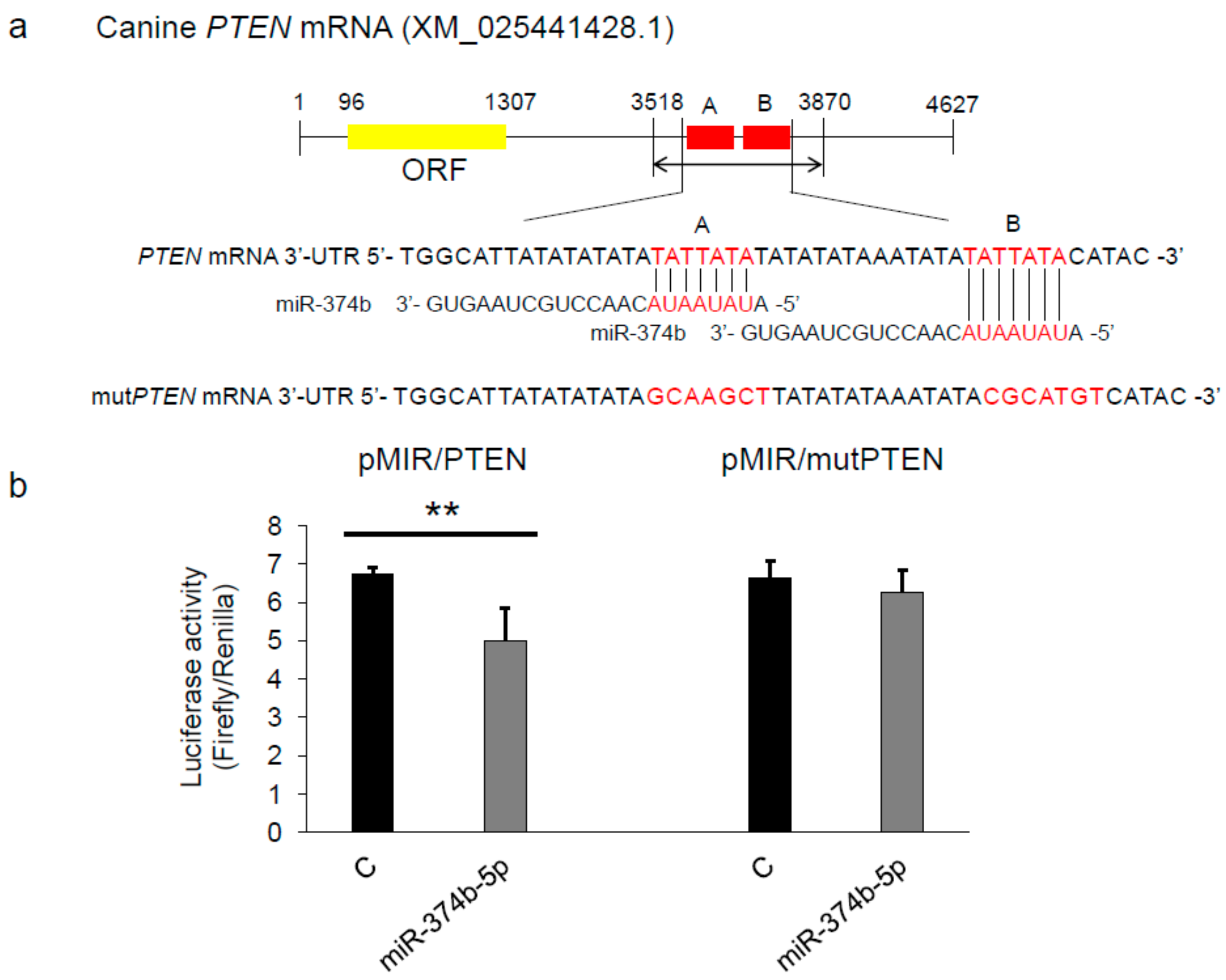
2.5. miR-374b-5p Directly Targeted PTEN
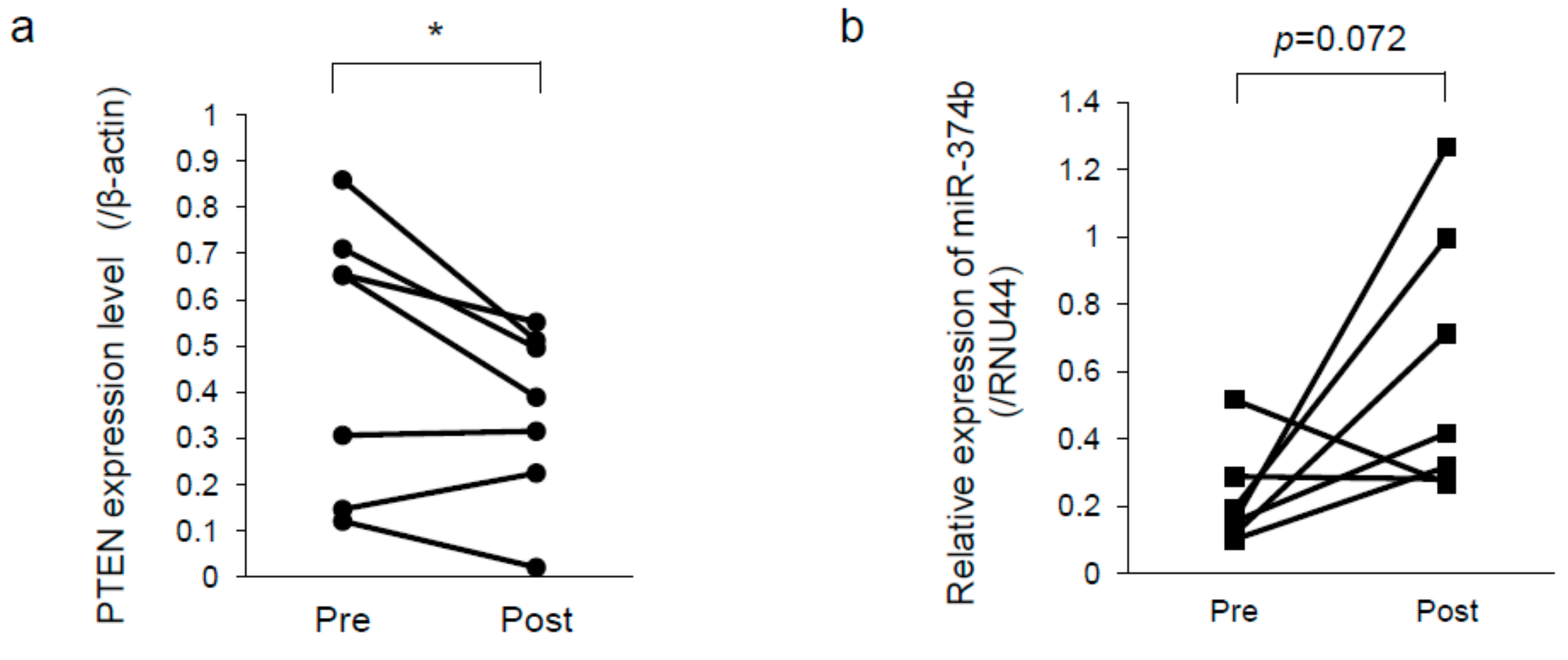
2.6. PTEN was Downregulated and miR-374b Upregulated in Recurrent Melanoma
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Cell Viability
4.2. Cell Count and Clonogenic Assay
4.3. Development of the PTEN Overexpressed CoMM Cells
4.4. Reagents and Antibodies
4.5. Western Blotting
4.6. MicroRNA Microarray and Quantitative RT-PCR (qRT-PCR) Using Real-Time PCR
4.7. Luciferase Activity Assay
4.8. Clinical Specimens
4.9. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Withrow, S.J. Melanoma. In Small Animal Clinical Oncology, 5th ed.; Withrow, S.J., Vail, D.M., Page, R.L., Eds.; Elsevier: St. Louis, MO, USA, 2013; pp. 321–334. ISBN 978-1-4377-2362-5. [Google Scholar]
- Gillard, M.; Cadieu, E.; De Brito, C.; Abadie, J.; Vergier, B.; Devauchelle, P.; Degorce, F.; Dreano, S.; Primot, A.; Dorso, L.; et al. Naturally occurring melanomas in dogs as models for non-UV pathways of human melanomas. Pigment Cell Melanoma Res. 2014, 27, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, B.; Adissu, H.A.-O.; Wei, B.R.; Michael, H.T.; Merlino, G.; Simpson, R.M. Naturally Occurring Canine Melanoma as a Predictive Comparative Oncology Model for Human Mucosal and Other Triple Wild-Type Melanomas. Int. J. Mol. Sci. 2018, 19, 394. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, M.; Mori, T.; Ito, Y.; Murakami, M.; Sakai, H.; Yanai, T.; Maruo, K. Outcomes of dogs undergoing radiotherapy for treatment of oral malignant melanoma: 111 cases (2006–2012). J. Am. Vet. Med. Assoc. 2015, 247, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.; Parsons, R. PTEN: Life as a tumor suppressor. Exp. Cell Res. 2001, 264, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, X.; Yu, S.; Wang, R.; Zhao, R.; Du, C. microRNA-222 promotes tumor growth and confers radioresistance in nasopharyngeal carcinoma by targeting PTEN. Mol. Med. Rep. 2018, 17, 1305–1310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xue, X.; Peng, H. Therapeutic Delivery of miR-29b Enhances Radiosensitivity in Cervical Cancer. Mol. Ther. 2019, 27, 1183–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirmohammadsadegh, A.; Marini, A.; Nambiar, S.; Hassan, M.; Tannapfel, A.; Ruzicka, T.; Hengge, U.R. Epigenetic silencing of the PTEN gene in melanoma. Cancer Res. 2006, 66, 6546–6552. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Calderon, L.G.; Fonseca-Alves, C.E.; Kobayashi, P.E.; Carvalho, M.; Drigo, S.A.; de Oliveira Vasconcelos, R.; Laufer-Amorim, R. Alterations in PTEN, MDM2, TP53 and AR protein and gene expression are associated with canine prostate carcinogenesis. Res. Vet. Sci. 2016, 106, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borge, K.S.; Nord, S.; Van Loo, P.; Lingjaerde, O.C.; Gunnes, G.; Alnaes, G.I.; Solvang, H.K.; Luders, T.; Kristensen, V.N.; Borresen-Dale, A.L.; et al. Canine Mammary Tumours Are Affected by Frequent Copy Number Aberrations, including Amplification of MYC and Loss of PTEN. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Mori, T.; Hoshino, Y.; Yamada, N.; Maruo, K.; Akao, Y. MicroRNAs as tumour suppressors in canine and human melanoma cells and as a prognostic factor in canine melanomas. Vet. Comp. Oncol. 2013, 11, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.L.; Tsai, Y.M.; Lien, C.T.; Kuo, P.L.; Hung, A.J. The Roles of MicroRNA in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, L.; Rodriguez-Aguayo, C.; Yuan, Y.; Debeb, B.G.; Chen, D.; Sun, Y.; You, M.J.; Liu, Y.; Dean, D.C.; et al. miR-205 acts as a tumour radiosensitizer by targeting ZEB1 and Ubc13. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Bode, A.M.; Cao, Y.; Dong, Z. Regulatory mechanisms and clinical perspectives of miRNA in tumor radiosensitivity. Carcinogenesis 2012, 33, 2220–2227. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Noguchi, S.; Nishiyama, Y.; Matsuyama, S.; Mori, T.; Igase, M.; Mizuno, T.; Shimamura, S.; Shimada, T. MicroRNA-205 enhances the radiosensitivity of canine melanoma cells by inhibiting E2F1. Jpn. J. Vet. Res. 2019, 67, 151–161. [Google Scholar]
- Nguyen, S.M.; Thamm, D.H.; Vail, D.M.; London, C.A. Response evaluation criteria for solid tumours in dogs (v1.0): A Veterinary Cooperative Oncology Group (VCOG) consensus document. Vet. Comp. Oncol. 2015, 13, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Li, G.C.; Cao, X.Y.; Li, Y.N.; Qiu, Y.Y.; Li, Y.N.; Liu, X.J.; Sun, X.X.A.-O.h.o.o. MicroRNA-374b inhibits cervical cancer cell proliferation and induces apoptosis through the p38/ERK signaling pathway by binding to JAM-2. J. Cell Physiol. 2018, 233, 7379–7390. [Google Scholar] [CrossRef] [PubMed]
- Long, Z.W.; Wu, J.H.; Cai, H.; Wang, Y.N.; Zhou, Y. MiR-374b Promotes Proliferation and Inhibits Apoptosis of Human GIST Cells by Inhibiting PTEN through Activation of the PI3K/Akt Pathway. Mol. Cells 2018, 41, 532–544. [Google Scholar] [PubMed]
- Qu, R.; Hao, S.; Jin, X.; Shi, G.; Yu, Q.; Tong, X.; Guo, D. MicroRNA-374b reduces the proliferation and invasion of colon cancer cells by regulation of LRH-1/Wnt signaling. Gene 2018, 642, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.R.; Michael, H.T.; Halsey, C.H.; Peer, C.J.; Adhikari, A.; Dwyer, J.E.; Hoover, S.B.; El Meskini, R.; Kozlov, S.; Weaver Ohler, Z.; et al. Synergistic targeted inhibition of MEK and dual PI3K/mTOR diminishes viability and inhibits tumor growth of canine melanoma underscoring its utility as a preclinical model for human mucosal melanoma. Pigment Cell Melanoma Res. 2016, 29, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Mori, T.; Hoshino, Y.; Maruo, K.; Yamada, N.; Kitade, Y.; Naoe, T.; Akao, Y. MicroRNA-143 functions as a tumor suppressor in human bladder cancer T24 cells. Cancer Lett. 2011, 307, 211–220. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noguchi, S.; Ogusu, R.; Wada, Y.; Matsuyama, S.; Mori, T. PTEN, A Target of Microrna-374b, Contributes to the Radiosensitivity of Canine Oral Melanoma Cells. Int. J. Mol. Sci. 2019, 20, 4631. https://doi.org/10.3390/ijms20184631
Noguchi S, Ogusu R, Wada Y, Matsuyama S, Mori T. PTEN, A Target of Microrna-374b, Contributes to the Radiosensitivity of Canine Oral Melanoma Cells. International Journal of Molecular Sciences. 2019; 20(18):4631. https://doi.org/10.3390/ijms20184631
Chicago/Turabian StyleNoguchi, Shunsuke, Ryo Ogusu, Yusuke Wada, Satoshi Matsuyama, and Takashi Mori. 2019. "PTEN, A Target of Microrna-374b, Contributes to the Radiosensitivity of Canine Oral Melanoma Cells" International Journal of Molecular Sciences 20, no. 18: 4631. https://doi.org/10.3390/ijms20184631
APA StyleNoguchi, S., Ogusu, R., Wada, Y., Matsuyama, S., & Mori, T. (2019). PTEN, A Target of Microrna-374b, Contributes to the Radiosensitivity of Canine Oral Melanoma Cells. International Journal of Molecular Sciences, 20(18), 4631. https://doi.org/10.3390/ijms20184631