Insights into the Functions of LncRNAs in Drosophila

 ,
,
Abstract
:1. Introduction
2. Conserved Secondary Structure Could Account for Functional Similarities
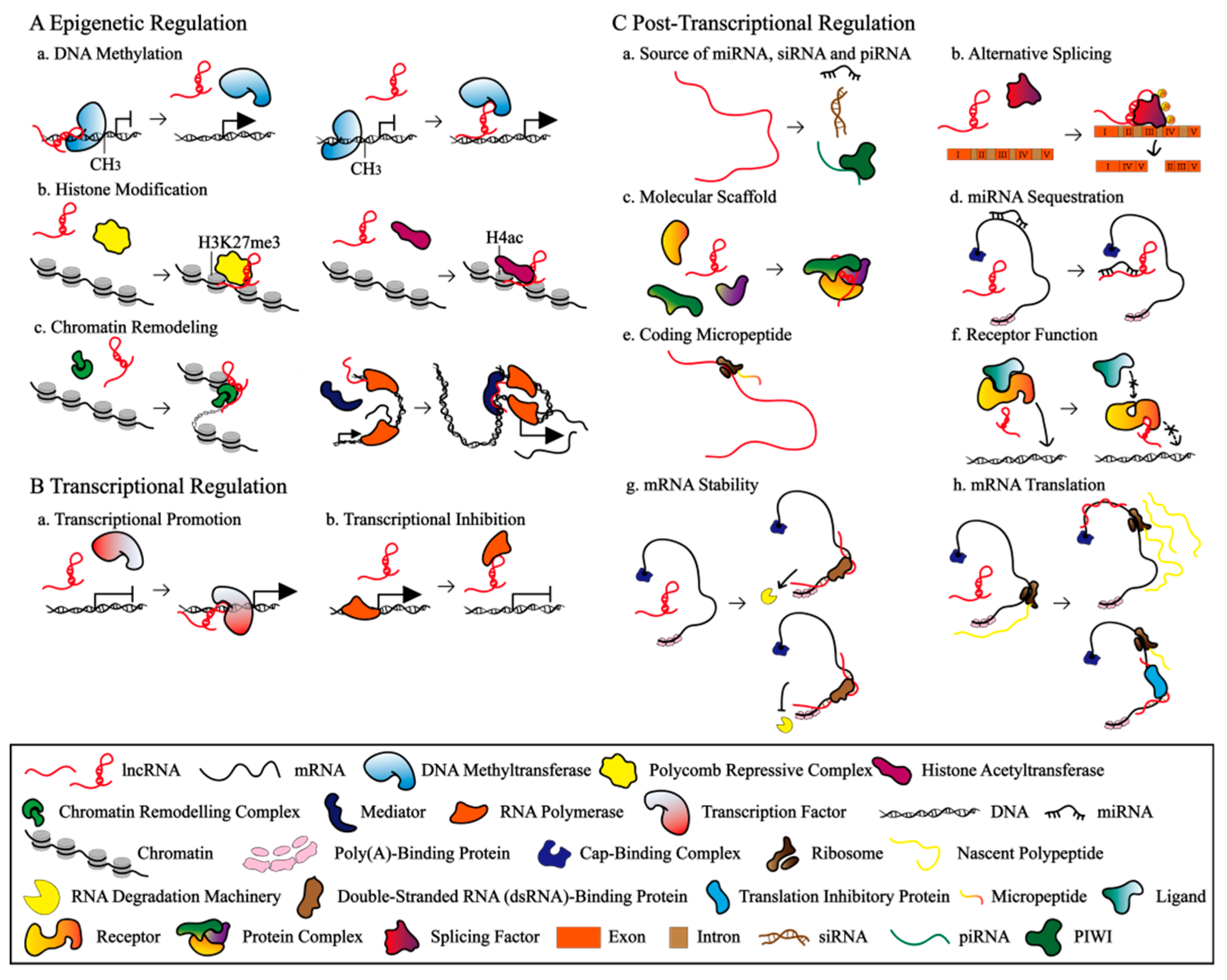
3. Three Main Mechanisms of LncRNAs
3.1. Epigenetic Regulation
3.2. Transcriptional Regulation
3.3. Post-Transcriptional Regulation
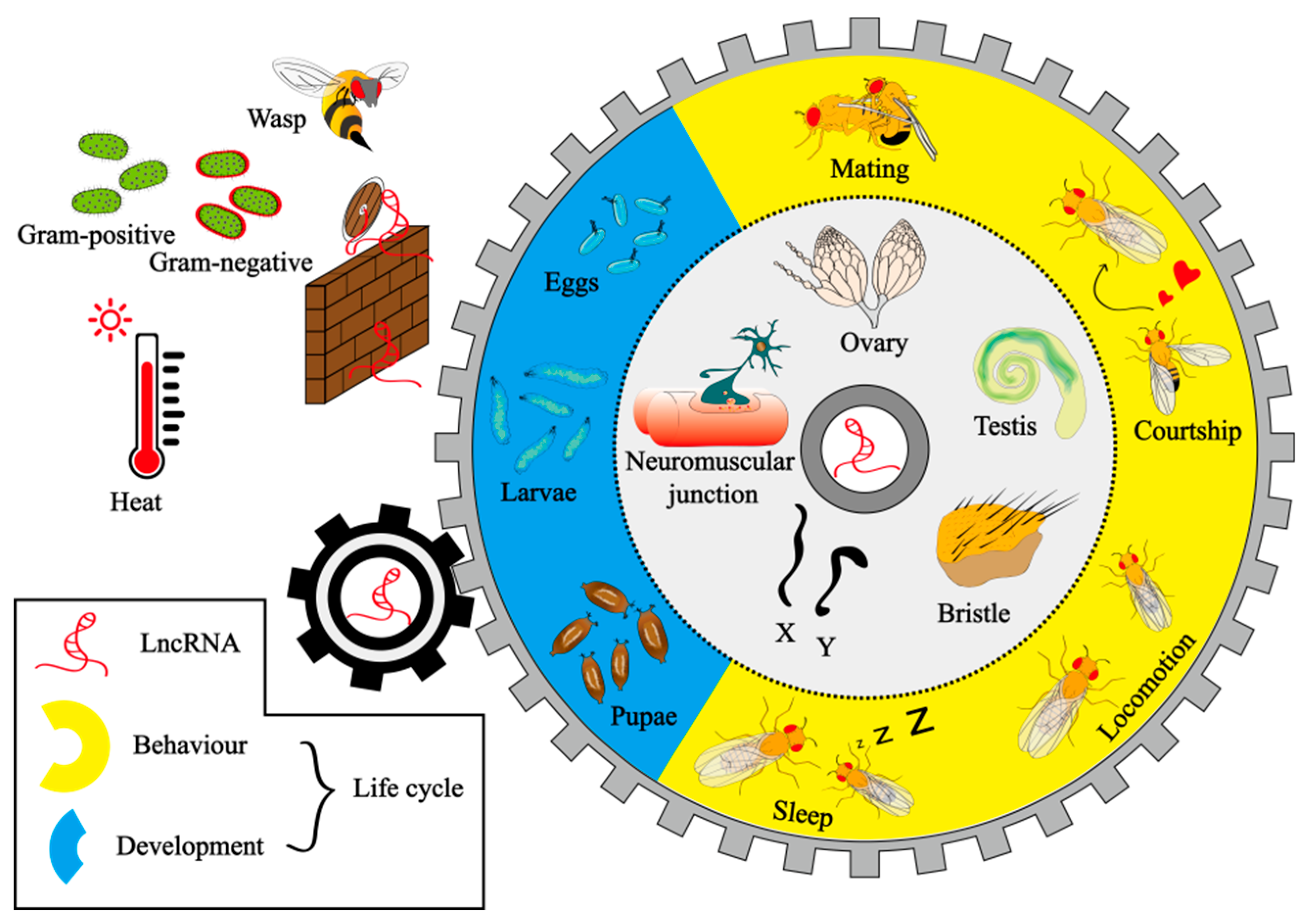
4. LncRNAs Mainly Associated with Drosophila Gonads and Development by Transcriptomics Analysis
4.1. Gonads
4.2. Development
4.3. Ageing and Neurogenesis
5. Biological Functions of LncRNAs in Drosophila
5.1. Embryonic Development
5.1.1. LncRNA-bxd
5.1.2. LncRNA-lincX
5.1.3. LncRNA-acal
5.1.4. LncRNA-ASTR
5.1.5. AAGAG Repeats RNAs
5.1.6. LncRNA-vg-PRE/TRE
5.1.7. LncRNA-7SK
5.2. Neurodegenerative Disease
5.2.1. LncRNA-hsrω
5.2.2. LncRNA-CR18854
5.3. Behaviour
5.3.1. LincRNA-yar
5.3.2. LncRNA-CRG
5.3.3. LncRNA-Sphinx
5.3.4. LncRNA-iab-8
5.4. Gonads
5.4.1. LncRNA-msa
5.4.2. LncRNA-oskar
5.5. Sex Determination and Dosage Compensation
5.5.1. LncRNA-SxlPe-R1 and R2
5.5.2. LncRNA-roX1 and roX2
5.6. Bristle Morphogenesis
5.6.1. LncRNA-bereft
5.6.2. LncRNA-SMRG
5.7. Immunometabolism and Stress Resistance
5.7.1. LincRNA-IBIN
5.7.2. LncRNA-hsrω
6. Drosophila LncRNA-related Databases
7. Techniques and Methods of LncRNAs Study in Drosophila
7.1. LncRNA Identification
7.2. Function Verification
7.3. Mechanism Exploration
8. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Lan, W.; Li, M.; Zhao, K.; Liu, J.; Wu, F.; Pan, Y.; Wang, J. LDAP: A web server for lncRNA-disease association prediction. Bioinformatics 2016, 33, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Kung, J.T.Y.; Colognori, D.; Lee, J.T. Long Noncoding RNAs: Past, Present, and Future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, S.U.; Grote, P.; Herrmann, B.G. Mechanisms of long noncoding RNA function in development and disease. Cell. Mol. Life Sci. 2016, 73, 2491–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and abundance analysis of human lncRNAs at single-cell and single-molecule resolution. Genome Biol. 2015, 16, 20. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, H.; Fang, S.; Kang, Y.; Wu, W.; Hao, Y.; Li, Z.; Bu, D.; Sun, N.; Zhang, M.Q.; et al. NONCODE 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Res. 2015, 44, D203–D208. [Google Scholar] [CrossRef] [PubMed]
- Maass, P.G.; Rump, A.; Schulz, H.; Stricker, S.; Schulze, L.; Platzer, K.; Aydin, A.; Tinschert, S.; Goldring, M.B.; Luft, F.C.; et al. A misplaced lncRNA causes brachydactyly in humans. J. Clin. Investig. 2012, 122, 3990–4002. [Google Scholar] [CrossRef] [Green Version]
- Askarian-Amiri, M.E.; Crawford, J.; French, J.D.; Smart, C.E.; Smith, M.A.; Clark, M.B.; Ru, K.; Mercer, T.R.; Thompson, E.R.; Lakhani, S.R.; et al. SNORD-host RNA Zfas1 is a regulator of mammary development and a potential marker for breast cancer. RNA 2011, 17, 878–891. [Google Scholar] [CrossRef]
- Roote, J.; Prokop, A. How to design a genetic mating scheme: A basic training package for Drosophila genetics. G3 (Bethesda) 2013, 3, 353–358. [Google Scholar] [CrossRef]
- Rogoyski, O.M.; Pueyo, J.I.; Couso, J.P.; Newbury, S.F. Functions of long non-coding RNAs in human disease and their conservation in Drosophila development. Biochem. Soc. Trans. 2017, 45, 895–904. [Google Scholar] [CrossRef]
- Lee, S.; Bang, S.M.; Lee, J.W.; Cho, K.S. Evaluation of traditional medicines for neurodegenerative diseases using Drosophila models. Evid.-Based Complement. Altern. Med. 2014, 2014, 967462. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Krueger, B.J.; Sedore, S.C.; Brogie, J.E.; Rogers, J.T.; Rajendra, T.K.; Saunders, A.; Matera, A.G.; Lis, J.T.; Uguen, P.; et al. The Drosophila 7SK snRNP and the essential role of dHEXIM in development. Nucleic Acids Res. 2012, 40, 5283–5297. [Google Scholar] [CrossRef] [PubMed]
- Pease, B.; Borges, A.C.; Bender, W. Non-coding RNAs of the Ultrabithorax domain of the Drosophila bithorax complex. Genetics 2013, 195, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Piccolo, L.L.; Yamaguchi, M. RNAi of arcRNA hsrω affects sub-cellular localization of Drosophila FUS to drive neurodiseases. Exp. Neurol. 2017, 292, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, Y.; Nakamura, A.; Tanaka, R.; Suda, K.; Azuma, Y.; Kushimura, Y.; Lo Piccolo, L.; Yoshida, H.; Mizuta, I.; Tokuda, T.; et al. Genetic screening of the genes interacting with Drosophila FIG4 identified a novel link between CMT-causing gene and long noncoding RNAs. Exp. Neurol. 2018, 310, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Maeda, R.K.; Sitnik, J.L.; Frei, Y.; Prince, E.; Gligorov, D.; Wolfner, M.F.; Karch, F. The lncRNA male-specific abdominal plays a critical role in Drosophila accessory gland development and male fertility. PLoS Genet. 2018, 14, e1007519. [Google Scholar] [CrossRef] [PubMed]
- Lakhotia, S.C.; Mallik, M.; Singh, A.K.; Ray, M. The large noncoding hsrω-n transcripts are essential for thermotolerance and remobilization of hnRNPs, HP1 and RNA polymerase II during recovery from heat shock in Drosophila. Chromosoma 2012, 121, 49–70. [Google Scholar] [CrossRef]
- Valanne, S.; Salminen, T.S.; Järvelä-Stölting, M.; Vesala, L.; Rämet, M. Immune-inducible non-coding RNA molecule lincRNA-IBIN connects immunity and metabolism in Drosophila melanogaster. PLoS Pathog. 2019, 15, e1007504. [Google Scholar] [CrossRef]
- Johnsson, P.; Lipovich, L.; Grandér, D.; Morris, K.V. Evolutionary conservation of long non-coding RNAs; sequence, structure, function. Biochim. et Biophys. Acta (BBA)—Gen. Subj. 2014, 1840, 1063–1071. [Google Scholar] [CrossRef]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef]
- Feyder, M.; Goff, L.A. Investigating long noncoding RNAs using animal models. J. Clin. Investig. 2016, 126, 2783–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandivier, L.E.; Anderson, S.J.; Foley, S.W.; Gregory, B.D. The conservation and function of RNA secondary structure in plants. Annu. Rev. Plant Biol. 2016, 67, 463–488. [Google Scholar] [CrossRef] [PubMed]
- Wutz, A.; Rasmussen, T.P.; Jaenisch, R. Chromosomal silencing and localization are mediated by different domains of Xist RNA. Nat. Genet. 2002, 30, 167. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Merry, C.R.; Forrest, M.E.; Sabers, J.N.; Beard, L.; Gao, X.-H.; Hatzoglou, M.; Jackson, M.W.; Wang, Z.; Markowitz, S.D.; Khalil, A.M. DNMT1-associated long non-coding RNAs regulate global gene expression and DNA methylation in colon cancer. Hum. Mol. Genet. 2015, 24, 6240–6253. [Google Scholar] [CrossRef] [PubMed]
- Canzio, D.; Nwakeze, C.L.; Horta, A.; Rajkumar, S.M.; Coffey, E.L.; Duffy, E.E.; Duffié, R.; Monahan, K.; O’Keeffe, S.; Simon, M.D.; et al. Antisense lncRNA Transcription Mediates DNA Demethylation to Drive Stochastic Protocadherin α Promoter Choice. Cell 2019, 177, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Postepska-Igielska, A.; Giwojna, A.; Gasri-Plotnitsky, L.; Schmitt, N.; Dold, A.; Ginsberg, D.; Grummt, I. LncRNA Khps1 regulates expression of the proto-oncogene SPHK1 via triplex-mediated changes in chromatin structure. Mol. Cell 2015, 60, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Duan, X.; Holmes, J.A.; Li, W.; Lee, S.H.; Tu, Z.; Zhu, C.; Salloum, S.; Lidofsky, A.; Schaefer, E.A.; et al. A Long Noncoding RNA Regulates Hepatitis C Virus Infection Through Interferon Alpha-Inducible Protein 6. Hepatology 2019, 69, 1004–1019. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, Y.; Li, T.; Ma, Z.; Jia, H.; Chen, Q.; Zhao, Y.; Zhai, L.; Zhong, R.; Li, C. Long non-coding RNA Linc-RAM enhances myogenic differentiation by interacting with MyoD. Nat. Commun. 2017, 8, 14016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trimarchi, T.; Bilal, E.; Ntziachristos, P.; Fabbri, G.; Dalla-Favera, R.; Tsirigos, A.; Aifantis, I. Genome-wide mapping and characterization of Notch-regulated long noncoding RNAs in acute leukemia. Cell 2014, 158, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Sallam, T.; Jones, M.; Thomas, B.J.; Wu, X.; Gilliland, T.; Qian, K.; Eskin, A.; Casero, D. Transcriptional regulation of macrophage cholesterol efflux and atherogenesis by a long noncoding RNA. Nat. Med. 2018, 24, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Philippe, T.; Geneviève, B.; Wajid, B.; Anne, R.; Joseph, M.; Amine, N. Transcription regulation by the noncoding RNA SRG1 requires Spt2-dependent chromatin deposition in the wake of RNA polymerase II. Mol. Cell. Biol. 2011, 31, 1288–1300. [Google Scholar]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659. [Google Scholar] [CrossRef] [PubMed]
- Tam, O.H.; Aravin, A.A.; Stein, P.; Girard, A.; Murchison, E.P.; Cheloufi, S.; Hodges, E.; Anger, M.; Sachidanandam, R.; Schultz, R.M.; et al. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 2008, 453, 534–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Tomizawa, S.-I.; Mitsuya, K.; Totoki, Y.; Yamamoto, Y.; Kuramochi-Miyagawa, S.; Iida, N.; Hoki, Y.; Murphy, P.J.; Toyoda, A. Role for piRNAs and noncoding RNA in de novo DNA methylation of the imprinted mouse Rasgrf1 locus. Science 2011, 332, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef] [PubMed]
- El Bassit, G.; Patel, R.S.; Carter, G.; Shibu, V.; Patel, A.A.; Song, S.; Murr, M.; Cooper, D.R.; Bickford, P.C.; Patel, N.A. MALAT1 in Human Adipose Stem Cells Modulates Survival and Alternative Splicing of PKC δ II in HT22 Cells. Endocrinology 2016, 158, 183–195. [Google Scholar]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef]
- Anderson, D.M.; Anderson, K.M.; Chang, C.-L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef]
- Matsumoto, A.; Pasut, A.; Matsumoto, M.; Yamashita, R.; Fung, J.; Monteleone, E.; Saghatelian, A.; Nakayama, K.I.; Clohessy, J.G.; Pandolfi, P.P. mTORC1 and muscle regeneration are regulated by the LINC00961-encoded SPAR polypeptide. Nature 2016, 541, 228. [Google Scholar] [CrossRef]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA gas5 is a growth arrest–and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal. 2010, 3, ra8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Y.; Lin, L.; Huang, Q.; He, W.; Zhang, S.; Dong, S.; Wen, Z.; Rao, J.; Liao, W.; et al. The lncRNA MACC1-AS1 promotes gastric cancer cell metabolic plasticity via AMPK/Lin28 mediated mRNA stability of MACC1. Mol. Cancer 2018, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, Z.; Trottier, J.; Barbier, O.; Wang, L. Long noncoding RNA MEG3 induces cholestatic liver injury by interaction with PTBP1 to facilitate shp mRNA decay. Hepatology 2017, 65, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Dimartino, D.; Colantoni, A.; Ballarino, M.; Martone, J.; Mariani, D.; Danner, J.; Bruckmann, A.; Meister, G.; Morlando, M.; Bozzoni, I. The Long Non-coding RNA lnc-31 Interacts with Rock1 mRNA and Mediates Its YB-1-Dependent Translation. Cell Rep. 2018, 23, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, R.; Sun, Y.; Qimuge, N.; Wang, G.; Wang, Y.; Chu, G.; Yu, T.; Yang, G.; Pang, W. Adiponectin AS lncRNA inhibits adipogenesis by transferring from nucleus to cytoplasm and attenuating Adiponectin mRNA translation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245. [Google Scholar] [CrossRef]
- Kaikkonen, M.U.; Lam, M.T.; Glass, C.K. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc. Res. 2011, 90, 430–440. [Google Scholar] [CrossRef] [Green Version]
- Lai, F.; Orom, U.A.; Cesaroni, M.; Beringer, M.; Taatjes, D.J.; Blobel, G.A.; Shiekhattar, R. Activating RNAs associate with Mediator to enhance chromatin architecture and transcription. Nature 2013, 494, 497–501. [Google Scholar] [CrossRef]
- Willingham, A.T.; Orth, A.P.; Batalov, S.; Peters, E.C.; Wen, B.G.; Aza-Blanc, P.; Hogenesch, J.B.; Schultz, P.G. A strategy for probing the function of noncoding RNAs finds a repressor of NFAT. Science 2005, 309, 1570–1573. [Google Scholar] [CrossRef]
- Shi, X.; Sun, M.; Wu, Y.; Yao, Y.; Liu, H.; Wu, G.; Yuan, D.; Song, Y. Post-transcriptional regulation of long noncoding RNAs in cancer. Tumor Biol. 2015, 36, 503–513. [Google Scholar] [CrossRef]
- Dey, B.K.; Pfeifer, K.; Dutta, A. The H19 long noncoding RNA gives rise to microRNAs miR-675–3p and miR-675–5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014, 28, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Fang, L. MEG3–4 is a miRNA decoy that regulates IL-1beta abundance to initiate and then limit inflammation to prevent sepsis during lung infection. Sci. Signal. 2018, 11, eaao2387. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.M.; Garcia, J.T.; Hung, T.; Flynn, R.A.; Shen, Y.; Qu, K.; Payumo, A.Y.; Peres-da-Silva, A.; Broz, D.K.; Baum, R.; et al. An inducible long noncoding RNA amplifies DNA damage signaling. Nat. Genet. 2016, 48, 1370–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.B.; Boley, N.; Eisman, R.; May, G.E.; Stoiber, M.H.; Duff, M.O.; Booth, B.W.; Wen, J.; Park, S.; Suzuki, A.M.; et al. Diversity and dynamics of the Drosophila transcriptome. Nature 2014, 512, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Vedelek, V.; Bodai, L.; Grezal, G.; Kovacs, B.; Boros, I.M.; Laurinyecz, B.; Sinka, R. Analysis of Drosophila melanogaster testis transcriptome. BMC Genom. 2018, 19, 697. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Yang, L.; Xiong, T.; Di, C.; Ma, D.; Wu, M.; Xue, Z.; Zhang, X.; Long, L.; Zhang, W.; et al. Critical roles of long noncoding RNAs in Drosophila spermatogenesis. Genome Res. 2016, 26, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Soumillon, M.; Necsulea, A.; Weier, M.; Brawand, D.; Zhang, X.; Gu, H.; Barthès, P.; Kokkinaki, M.; Nef, S.; Gnirke, A.; et al. Cellular Source and Mechanisms of High Transcriptome Complexity in the Mammalian Testis. Cell Rep. 2013, 3, 2179–2190. [Google Scholar] [CrossRef] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Lin, Y.; Wu, J. Long non-coding RNA expression profiling of mouse testis during postnatal development. PLoS ONE 2013, 8, e75750. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.; Zhang, X.; Jia, S.; Chen, S.; Kang, L. Genome-wide identification and developmental expression profiling of long noncoding RNAs during Drosophila metamorphosis. Sci. Rep. 2016, 6, 23330. [Google Scholar] [CrossRef]
- Batut, P.J.; Gingeras, T.R. Conserved noncoding transcription and core promoter regulatory code in early Drosophila development. eLife 2017, 6, e29005. [Google Scholar] [CrossRef] [PubMed]
- Schor, I.E.; Bussotti, G.; Males, M.; Forneris, M.; Viales, R.R.; Enright, A.J.; Furlong, E.E.M. Non-coding RNA Expression, Function, and Variation during Drosophila Embryogenesis. Curr. Biol. CB 2018, 28, 3547–3561. [Google Scholar] [CrossRef] [PubMed]
- Karaiskos, N.; Wahle, P. The Drosophila embryo at single-cell transcriptome resolution. Science 2017, 358, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Lian, T.; Tu, J.; Gaur, U.; Mao, X.; Fan, X.; Li, D.; Li, Y.; Yang, M. LncRNA mediated regulation of aging pathways in Drosophila melanogaster during dietary restriction. Aging (Albany NY) 2016, 8, 2182. [Google Scholar] [CrossRef] [PubMed]
- Davie, K.; Janssens, J.; Koldere, D.; De Waegeneer, M.; Pech, U.; Kreft, L.; Aibar, S.; Makhzami, S.; Christiaens, V.; Bravo Gonzalez-Blas, C.; et al. A Single-Cell Transcriptome Atlas of the Aging Drosophila Brain. Cell 2018, 174, 982–998. [Google Scholar] [CrossRef]
- McCorkindale, A.L.; Wahle, P.; Werner, S.; Jungreis, I.; Menzel, P.; Shukla, C.J.; Abreu, R.L.P.; Irizarry, R.A.; Meyer, I.M.; Kellis, M. A gene expression atlas of embryonic neurogenesis in Drosophila reveals complex spatiotemporal regulation of lncRNAs. Development 2019, 146, dev175265. [Google Scholar] [CrossRef]
- Pettini, T.; Ronshaugen, M.R. Transvection and pairing of a Drosophila Hox long noncoding RNA in the regulation of Sex combs reduced. bioRxiv 2016, 045617. [Google Scholar] [CrossRef]
- Ríos-Barrera, L.D.; Gutiérrez-Pérez, I.; Domínguez, M.; Riesgo-Escovar, J.R. acal is a long non-coding RNA in JNK signaling in epithelial shape changes during Drosophila dorsal closure. PLoS Genet. 2015, 11, e1004927. [Google Scholar] [CrossRef]
- Pek, J.W.; Osman, I.; Tay, M.L.; Zheng, R.T. Stable intronic sequence RNAs have possible regulatory roles in Drosophila melanogaster. J. Cell Biol. 2015, 211, 243–251. [Google Scholar] [CrossRef]
- Pathak, R.U.; Mamillapalli, A.; Rangaraj, N.; Kumar, R.P.; Vasanthi, D.; Mishra, K.; Mishra, R.K. AAGAG repeat RNA is an essential component of nuclear matrix in Drosophila. RNA Biol. 2013, 10, 564–571. [Google Scholar] [CrossRef]
- Herzog, V.A.; Lempradl, A.; Trupke, J.; Okulski, H.; Altmutter, C.; Ruge, F.; Boidol, B.; Kubicek, S.; Schmauss, G.; Aumayr, K.; et al. A strand-specific switch in noncoding transcription switches the function of a Polycomb/Trithorax response element. Nat. Genet. 2014, 46, 973–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardiman, K.E.; Brewster, R.; Khan, S.M.; Deo, M.; Bodmer, R. The bereft gene, a potential target of the neural selector gene cut, contributes to bristle morphogenesis. Genetics 2002, 161, 231–247. [Google Scholar] [PubMed]
- Xu, M.; Xiang, Y.; Liu, X.; Bai, B.; Chen, R.; Liu, L.; Li, M. Long noncoding RNA SMRG regulates Drosophila macrochaetes by antagonizing scute through E (spl) mβ. RNA Biol. 2019, 16, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Jenny, A.; Hachet, O.; Zavorszky, P.; Cyrklaff, A.; Weston, M.D.; Johnston, D.S.; Erdelyi, M.; Ephrussi, A. A translation-independent role of oskar RNA in early Drosophila oogenesis. Development 2006, 133, 2827–2833. [Google Scholar] [CrossRef] [PubMed]
- Lo Piccolo, L.; Jantrapirom, S.; Nagai, Y.; Yamaguchi, M. FUS toxicity is rescued by the modulation of lncRNA hsrω expression in Drosophila melanogaster. Sci. Rep. 2017, 7, 15660. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wen, S.; Guo, X.; Bai, B.; Gong, Z.; Liu, X.; Wang, Y.; Zhou, Y.; Chen, X.; Liu, L. The novel long non-coding RNA CRG regulates Drosophila locomotor behavior. Nucleic Acids Res. 2012, 40, 11714–11727. [Google Scholar] [CrossRef] [PubMed]
- Gummalla, M.; Maeda, R.K.; Alvarez, J.J.C.; Gyurkovics, H.; Singari, S.; Edwards, K.A.; Karch, F.; Bender, W. abd-A regulation by the iab-8 noncoding RNA. PLoS Genet. 2012, 8, e1002720. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Meller, V.H. roX RNAs are required for increased expression of X-linked genes in Drosophila melanogaster males. Genetics 2006, 174, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Dai, H.; Chen, S.; Zhang, L.; Long, M. Highly tissue specific expression of Sphinx supports its male courtship related role in Drosophila melanogaster. PLoS ONE 2011, 6, e18853. [Google Scholar] [CrossRef]
- Mulvey, B.B.; Olcese, U.; Cabrera, J.R.; Horabin, J.I. An interactive network of long non-coding RNAs facilitates the Drosophila sex determination decision. Biochim. et Biophys. Acta (BBA)-Gene Regul. Mech. 2014, 1839, 773–784. [Google Scholar] [CrossRef]
- Soshnev, A.A.; Ishimoto, H.; McAllister, B.F.; Li, X.; Wehling, M.D.; Kitamoto, T.; Geyer, P.K. A conserved long noncoding RNA affects sleep behavior in Drosophila. Genetics 2011, 189, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Petruk, S.; Sedkov, Y.; Riley, K.M.; Hodgson, J.; Schweisguth, F.; Hirose, S.; Jaynes, J.B.; Brock, H.W.; Mazo, A. Transcription of bxd noncoding RNAs promoted by trithorax represses Ubx in cis by transcriptional interference. Cell 2006, 127, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Beltran, M.; Garcia de Herreros, A. Antisense non-coding RNAs and regulation of gene transcription. Transcription 2016, 7, 39–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedore, S.C.; Byers, S.A.; Biglione, S.; Price, J.P.; Maury, W.J.; Price, D.H. Manipulation of P-TEFb control machinery by HIV: Recruitment of P-TEFb from the large form by Tat and binding of HEXIM1 to TAR. Nucleic Acids Res. 2007, 35, 4347–4358. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.A.; Poirier, M.A. Protein aggregation and neurodegenerative disease. Nat. Med. 2004, 10, S10. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.A.; Leyns, C.E.G.; Holtzman, D.M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annu. Rev. Cell Dev. Biol. 2018, 34, 545–568. [Google Scholar] [CrossRef]
- Abe, A.; Numakura, C.; Kijima, K.; Hayashi, M.; Hashimoto, T.; Hayasaka, K. Molecular diagnosis and clinical onset of Charcot-Marie-Tooth disease in Japan. J. Hum. Genet. 2011, 56, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Nichols, C.D.; Becnel, J.; Pandey, U.B. Methods to Assay Drosophila Behavior. JoVE 2012, e3795. [Google Scholar] [CrossRef]
- Lewis, E.B. A gene complex controlling segmentation in Drosophila. Nature 1978, 276, 565–570. [Google Scholar] [CrossRef]
- Sampath, K.; Ephrussi, A. CncRNAs: RNAs with both coding and non-coding roles in development. Development 2016, 143, 1234–1241. [Google Scholar] [CrossRef] [Green Version]
- Gajjar, M.; Candeias, M.M.; Malbert-Colas, L.; Mazars, A.; Fujita, J.; Olivares-Illana, V.; Fåhraeus, R. The p53 mRNA-Mdm2 interaction controls Mdm2 nuclear trafficking and is required for p53 activation following DNA damage. Cancer Cell 2012, 21, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Nagano, H.; Yamagishi, N.; Tomida, C.; Yano, C.; Aibara, K.; Kohno, S.; Abe, T.; Ohno, A.; Hirasaka, K.; Okumura, Y. A novel myogenic function residing in the 5′ non-coding region of Insulin receptor substrate-1 (Irs-1) transcript. BMC Cell Biol. 2015, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Ephrussi, A.; Dickinson, L.K.; Lehmann, R. Oskar organizes the germ plasm and directs localization of the posterior determinant nanos. Cell 1991, 66, 37–50. [Google Scholar] [CrossRef]
- St Johnston, D.; Beuchle, D.; Nusslein-Volhard, C. Staufen, a gene required to localize maternal RNAs in the Drosophila egg. Cell 1991, 66, 51–63. [Google Scholar] [CrossRef]
- Baker, B.S.; Belote, J.M. SEX DETERMINATION AND DOSAGE COMPENSATION IN DROSOPHILA MELANOGASTER. Annu. Rev. Genet. 1983, 17, 345–393. [Google Scholar] [CrossRef]
- Pontier, D.B.; Gribnau, J. Xist regulation and function explored. Hum. Genet. 2011, 130, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Faucillion, M.-L.; Larsson, J. RNA-on-X 1 and 2 in Drosophila melanogaster fulfill separate functions in dosage compensation. PLoS Genet. 2018, 14, e1007842. [Google Scholar] [CrossRef]
- Amaral, P.P.; Leonardi, T.; Han, N.; Viré, E.; Gascoigne, D.K.; Arias-Carrasco, R.; Büscher, M.; Pandolfini, L.; Zhang, A.; Pluchino, S. Genomic positional conservation identifies topological anchor point RNAs linked to developmental loci. Genome Biol. 2018, 19, 32. [Google Scholar] [CrossRef]
- Schweisguth, F. Asymmetric cell division in the Drosophila bristle lineage: From the polarization of sensory organ precursor cells to Notch-mediated binary fate decision. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 299–309. [Google Scholar] [CrossRef]
- Mummery-Widmer, J.L.; Yamazaki, M.; Stoeger, T.; Novatchkova, M.; Bhalerao, S.; Chen, D.; Dietzl, G.; Dickson, B.J.; Knoblich, J.A. Genome-wide analysis of Notch signalling in Drosophila by transgenic RNAi. Nature 2009, 458, 987–992. [Google Scholar] [CrossRef]
- Leader, D.P.; Krause, S.A.; Pandit, A.; Davies, S.A.; Dow, J.A.T. FlyAtlas 2: A new version of the Drosophila melanogaster expression atlas with RNA-Seq, miRNA-Seq and sex-specific data. Nucleic Acids Res. 2017, 46, D809–D815. [Google Scholar] [CrossRef] [PubMed]
- Murali, T.; Pacifico, S.; Yu, J.; Guest, S.; Roberts, G.G.; Finley, R.L. DroID 2011: A comprehensive, integrated resource for protein, transcription factor, RNA and gene interactions for Drosophila. Nucleic Acids Res. 2010, 39, D736–D743. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, G.; Li, J.; Zhang, X.; Huang, S.; Xiang, S.; Hu, X.; Liu, C. CRISPRlnc: A manually curated database of validated sgRNAs for lncRNAs. Nucleic Acids Res. 2018, 47, D63–D68. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Wu, W.; Li, H.; Yuan, J.; Luo, J.; Zhao, Y.; Chen, R. NPInter v3. 0: An upgraded database of noncoding RNA-associated interactions. Database 2016, 2016, baw057. [Google Scholar] [CrossRef] [PubMed]
- Tweedie, S.; Ashburner, M.; Falls, K.; Leyland, P.; McQuilton, P.; Marygold, S.; Millburn, G.; Osumi-Sutherland, D.; Schroeder, A.; Seal, R. FlyBase: Enhancing Drosophila gene ontology annotations. Nucleic Acids Res. 2008, 37, D555–D559. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Bai, B.; Skogerbø, G.; Cai, L.; Deng, W.; Zhang, Y.; Bu, D.; Zhao, Y.; Chen, R. NONCODE: An integrated knowledge database of non-coding RNAs. Nucleic Acids Res. 2005, 33, D112–D115. [Google Scholar] [CrossRef] [PubMed]
- Amaral, P.P.; Clark, M.B.; Gascoigne, D.K.; Dinger, M.E.; Mattick, J.S. lncRNAdb: A reference database for long noncoding RNAs. Nucleic Acids Res. 2010, 39, D146–D151. [Google Scholar] [CrossRef]
- Chen, X.; Hao, Y.; Cui, Y.; Fan, Z.; He, S.; Luo, J.; Chen, R. LncVar: A database of genetic variation associated with long non-coding genes. Bioinformatics 2016, 33, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Liu, C.; Liu, W.; Xiang, Y.; Diao, L.; Guo, A.-Y.; Han, L. LNCediting: A database for functional effects of RNA editing in lncRNAs. Nucleic Acids Res. 2016, 45, D79–D84. [Google Scholar] [CrossRef]
- Yang, J.; Li, J.; Jiang, S.; Zhou, H.; Qu, L. ChIPBase: A database for decoding the transcriptional regulation of long non-coding RNA and microRNA genes from ChIP-Seq data. Nucleic Acids Res. 2012, 41, D177–D187. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.T.; Di, C.; Hu, B.; Zhou, M.; Liu, Y.; Song, N.; Li, Y.; Umetsu, J.; Lu, Z.J. CLIPdb: A CLIP-seq database for protein-RNA interactions. BMC Genom. 2015, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Yu, N.; Choi, I.; Kim, W.; Lee, S. lncRNAtor: A comprehensive resource for functional investigation of long non-coding RNAs. Bioinformatics 2014, 30, 2480–2485. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Wang, Z.; Guo, J. The research strategies for probing the function of long noncoding RNAs. Genomics 2012, 99, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Aprile, M.; Esposito, R.; Ciccodicola, A. RNA-Seq and human complex diseases: Recent accomplishments and future perspectives. Eur. J. Hum. Genet. 2013, 21, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Zhou, X.; Du, X. Circulating long non-coding RNAs in cancer: Current status and future perspectives. Mol. Cancer 2016, 15, 39. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Tibbit, C.; Liu, J. Effective knockdown of Drosophila long non-coding RNAs by CRISPR interference. Nucleic Acids Res. 2016, 44, e84. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Qu, K.; Zhong, F.L.; Artandi, S.E.; Chang, H.Y. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Spitale, R.C.; Chang, H.Y. Technologies to probe functions and mechanisms of long noncoding RNAs. Nat. Struct. Mol. Biol. 2015, 22, 29. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.N.; Sattler, M. Challenges and perspectives for structural biology of lncRNAs–the example of the Xist lncRNA A-repeats. J. Mol. Cell Biol. 2019, mjz086. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Transcripts | Related Genes/Proteins | Function | Reference |
|---|---|---|---|
| AAGAG repeat RNAs | - | Nuclear matrix constituents | [70] |
| acal | Cka, aop, raw | Involved in dorsal closure | [68] |
| ASTR | regena, sisR-1 | Regulates regena gene transcripts | [69] |
| bereft | cut, numb, tramtrack | Involved in bristle morphogenesis | [72] |
| bxd | Ubx | Regulates growth and development | [13] |
| CRG | CASK, RNA pol II | Regulates locomotor activity and climbing ability | [76] |
| CR18854 | dFUS, dFIG4 | Suppresses the rough eye and the loss-of-cone cell phenotype caused by dFIG4 deficiency | [15] |
| hsrω | hnRNPs, HP1, RNA pol II | Responds to heat shock | [17] |
| hsrω | dFUS | Regulates the development of neuromuscular junctions | [14] |
| iab-8 | miR-iab-8, abd-A | Regulates the mating behaviour | [77] |
| IBIN | Toll, BAP, Brahma | Acts as a link between innate immune responses and metabolism | [18] |
| lincX | Scr | Involved in the activation of Scr | [67] |
| msa | miR-iab-8 | Involved in accessory gland development and male fertility | [16] |
| oskar | Bruno, Staufen | Regulates oogenesis | [74] |
| roX1 and roX2 | Msl, Mof, Mle | Involved in dosage compensation | [78] |
| SMRG | scute, E(spl)mβ | Regulates scutellar macrochaetes | [73] |
| sphinx | - | Involved in the regulation of male courtship behaviour | [79] |
| SxlPe-R1 and R2 | Sxl, PcG/trxG | Facilitates sex determination | [80] |
| vg-PRE/TRE | vg, PRC2, E(Z) | Regulates PRC2 activity | [71] |
| yar | - | Regulates sleep behaviour | [81] |
| 7SK | RNA pol II, P-TEFb | Regulates RNA pol II activity via P-TEFb | [12] |
| Name | Website | Description | Reference |
|---|---|---|---|
| FlyBase | http://flybase.org | A database of Drosophila genes and genomes | [105] |
| FlyAtlas 2 | http://www.flyatlas2.org | Gene expression pattern in fly tissues | [101] |
| DVEX | http://www.dvex.org | Single-cell expression atlas of lncRNAs of the stage 6 Drosophila | [63] |
| DroID | http://droidb.org/ | Interaction networks (protein–protein, TF–gene, and miRNA–gene) | [102] |
| NONCODE | http://www.noncode.org/ | Details of annotation of lncRNAs | [106] |
| lncRNAdb | http://www.lncrnadb.org/ | Information on RNAs related to nucleotide sequence, genomic context, gene expression data, structural information, subcellular localization, conservation, and function (validated data) | [107] |
| LncVar | http://bioinfo.ibp.ac.cn/LncVar/ | Systematically integrated information about transcription factor binding sites and m6A modification sites of lncRNAs and comprehensive effects of single nucleotide polymorphism (SNPs) on transcription and modification of lncRNAs | [108] |
| LNCediting | http://bioinfo.life.hust.edu.cn/LNCediting/ | A comprehensive resource for the functional prediction of RNAs editing in lncRNAs | [109] |
| ChIPBase v2.0 | http://rna.sysu.edu.cn/chipbase/ | Transcriptional regulatory networks of non-coding RNAs (ncRNAs) and protein-coding genes (PCGs) | [110] |
| CRISPRlnc | http://www.crisprlnc.org | Manually curated database of validated CRISPR/Cas9 sgRNAs for lncRNAs | [103] |
| NPInter v3.0 | http://www.bioinfo.org/NPInter/ | Experimentally verified interactions between ncRNAs (excluding transfer RNAs (tRNAs) and ribosomal RNAs (rRNAs)), especially lncRNAs and other biomolecules | [104] |
| CLIPdb | http://lulab.life.tsinghua.edu.cn/clipdb/ | Regulatory networks among RNA-binding proteins and various RNA transcripts | [111] |
| lncRNAtor | http://lncrnator.ewha.ac.kr/ | Information related to expression profiles, interacting (binding) proteins, integrated sequence curation, evolutionary scores and coding potential of lncRNAs | [112] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Tian, Y.; Yuan, Y.; Fan, X.; Yang, M.; He, Z.; Yang, D. Insights into the Functions of LncRNAs in Drosophila. Int. J. Mol. Sci. 2019, 20, 4646. https://doi.org/10.3390/ijms20184646
Li K, Tian Y, Yuan Y, Fan X, Yang M, He Z, Yang D. Insights into the Functions of LncRNAs in Drosophila. International Journal of Molecular Sciences. 2019; 20(18):4646. https://doi.org/10.3390/ijms20184646
Chicago/Turabian StyleLi, Keqin, Yuanliangzi Tian, Ya Yuan, Xiaolan Fan, Mingyao Yang, Zhi He, and Deying Yang. 2019. "Insights into the Functions of LncRNAs in Drosophila" International Journal of Molecular Sciences 20, no. 18: 4646. https://doi.org/10.3390/ijms20184646
APA StyleLi, K., Tian, Y., Yuan, Y., Fan, X., Yang, M., He, Z., & Yang, D. (2019). Insights into the Functions of LncRNAs in Drosophila. International Journal of Molecular Sciences, 20(18), 4646. https://doi.org/10.3390/ijms20184646