CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality

 ,
, 
Abstract
:1. Introduction
2. Results
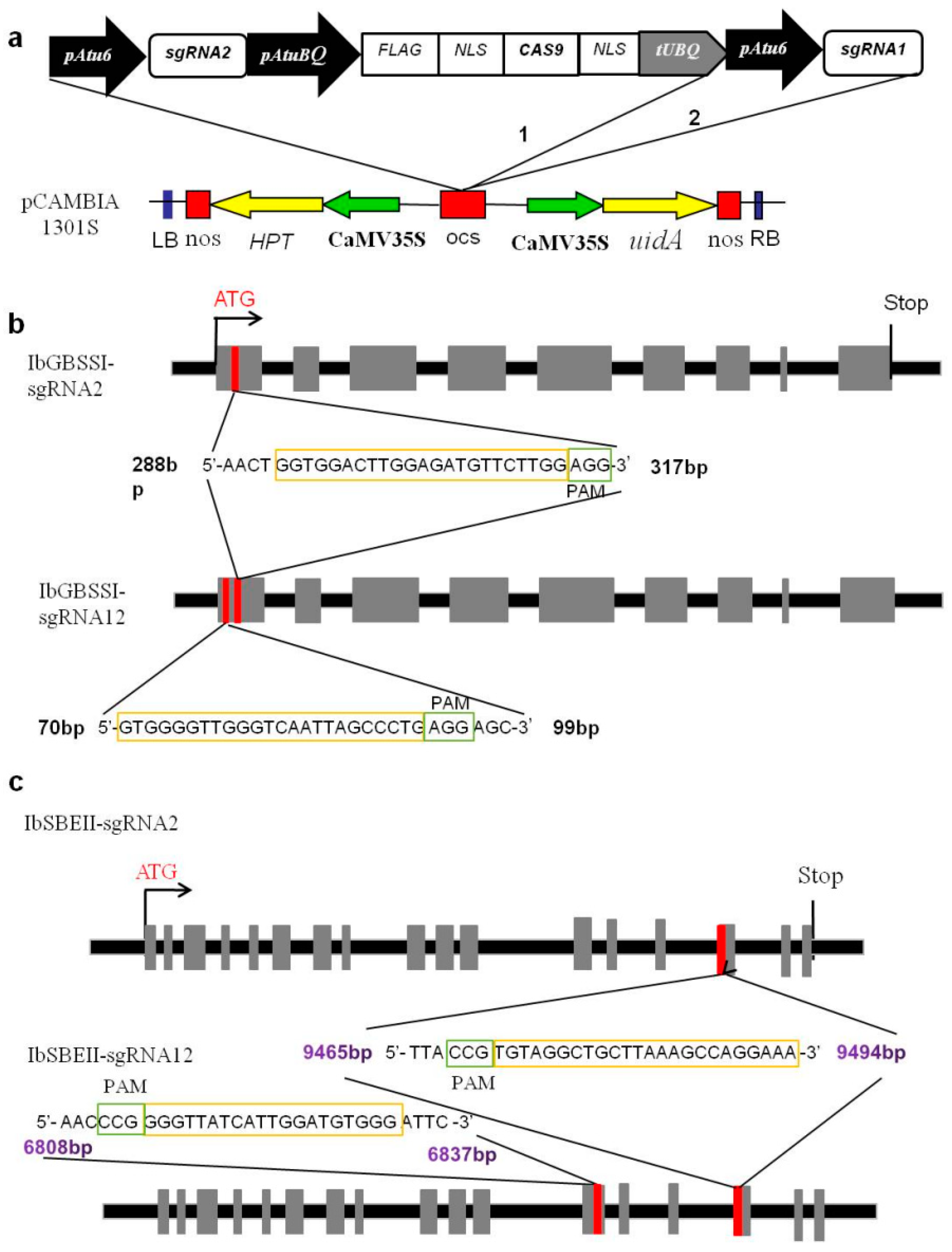
2.1. Single or Dual gRNA Vectors Allow Evaluation of Efficiency for Targeted Mutagenesis
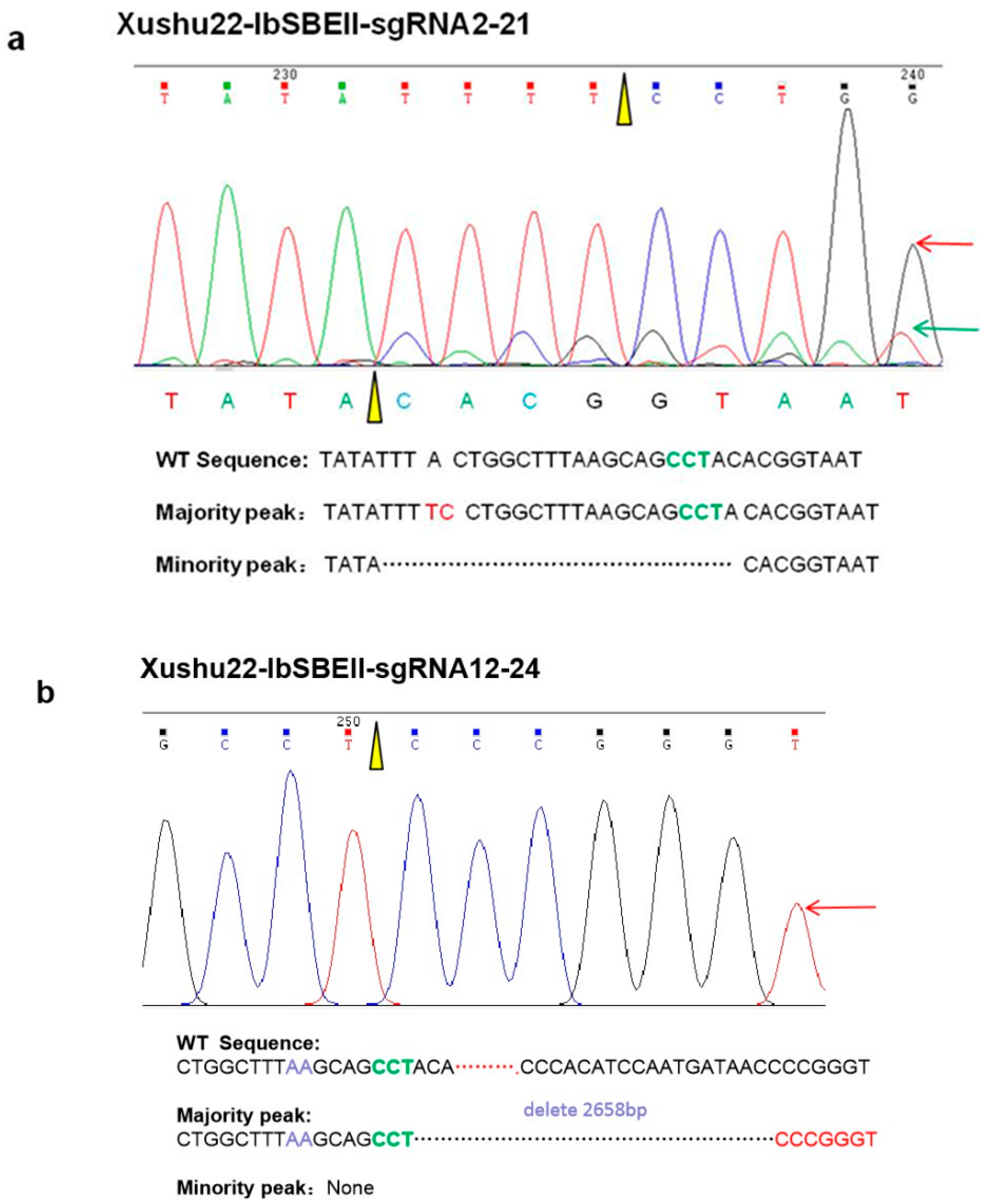
2.2. Efficient Gene Mutagenesis in Sweet Potato
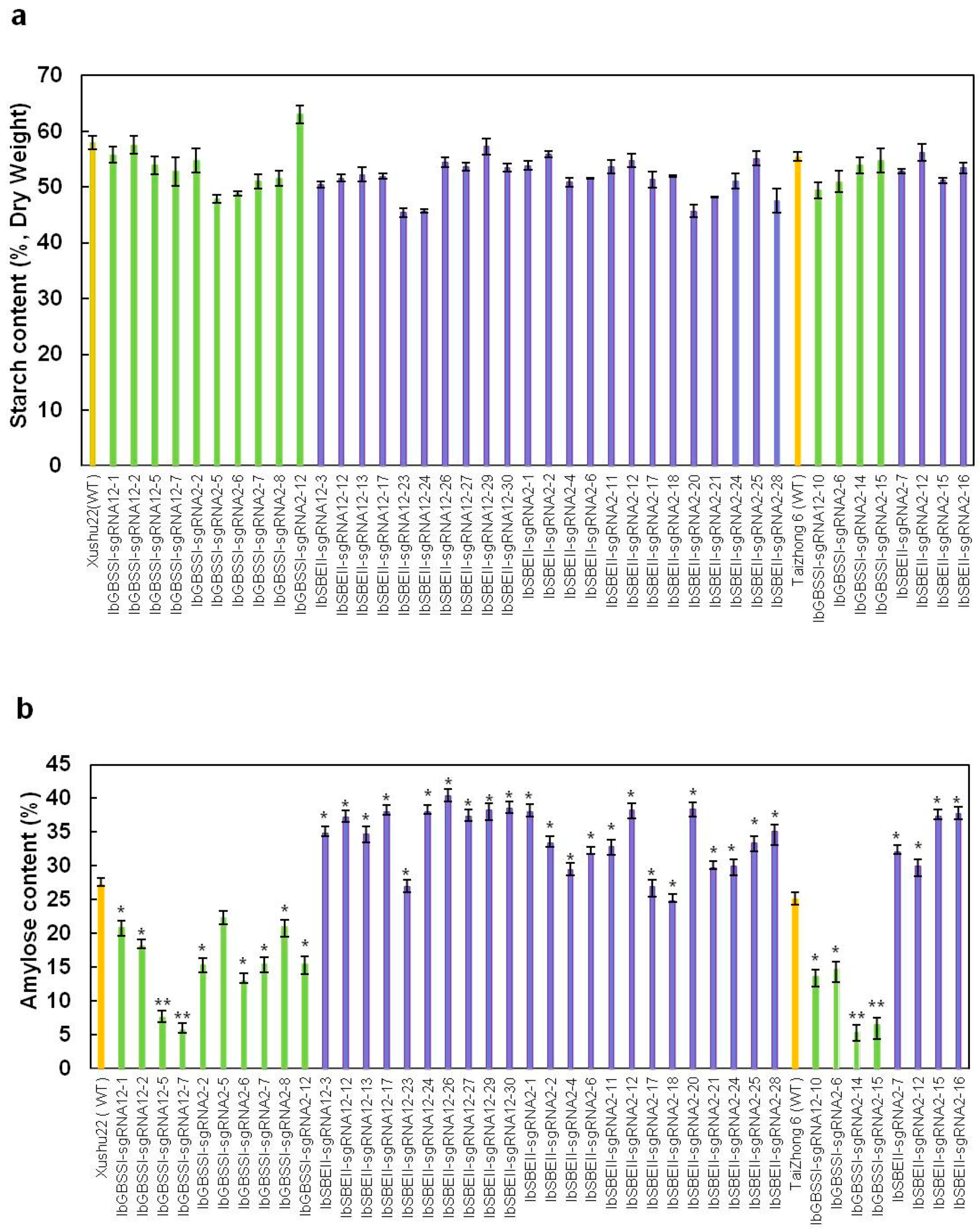
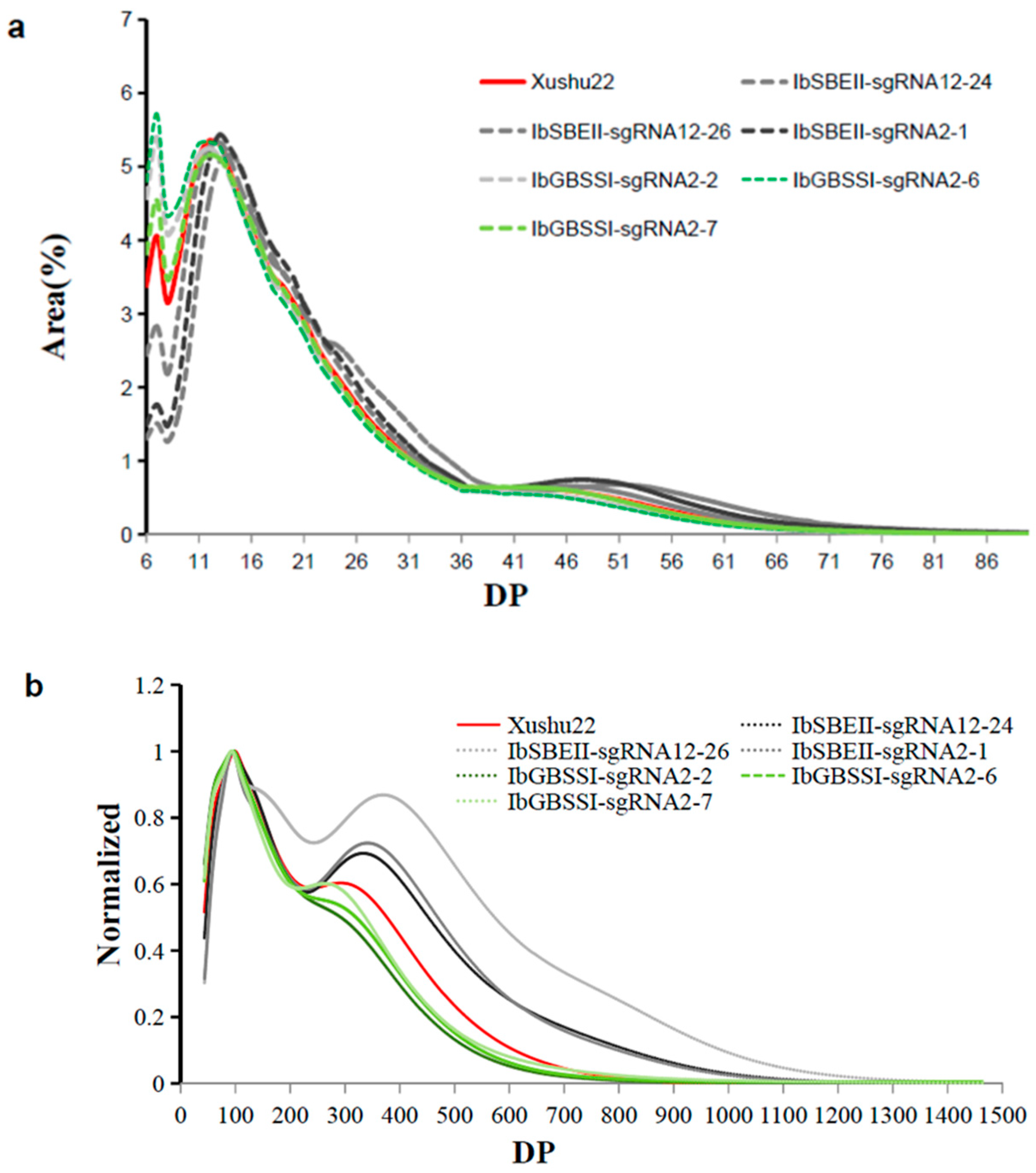
2.3. The Starch and Amylose Contents and Chain Length Distribution in Mutated Plants
3. Discussion
4. Materials and Methods
4.1. Vector Construction
4.2. Agrobacterium-Mediated Transformation of Sweet Potato
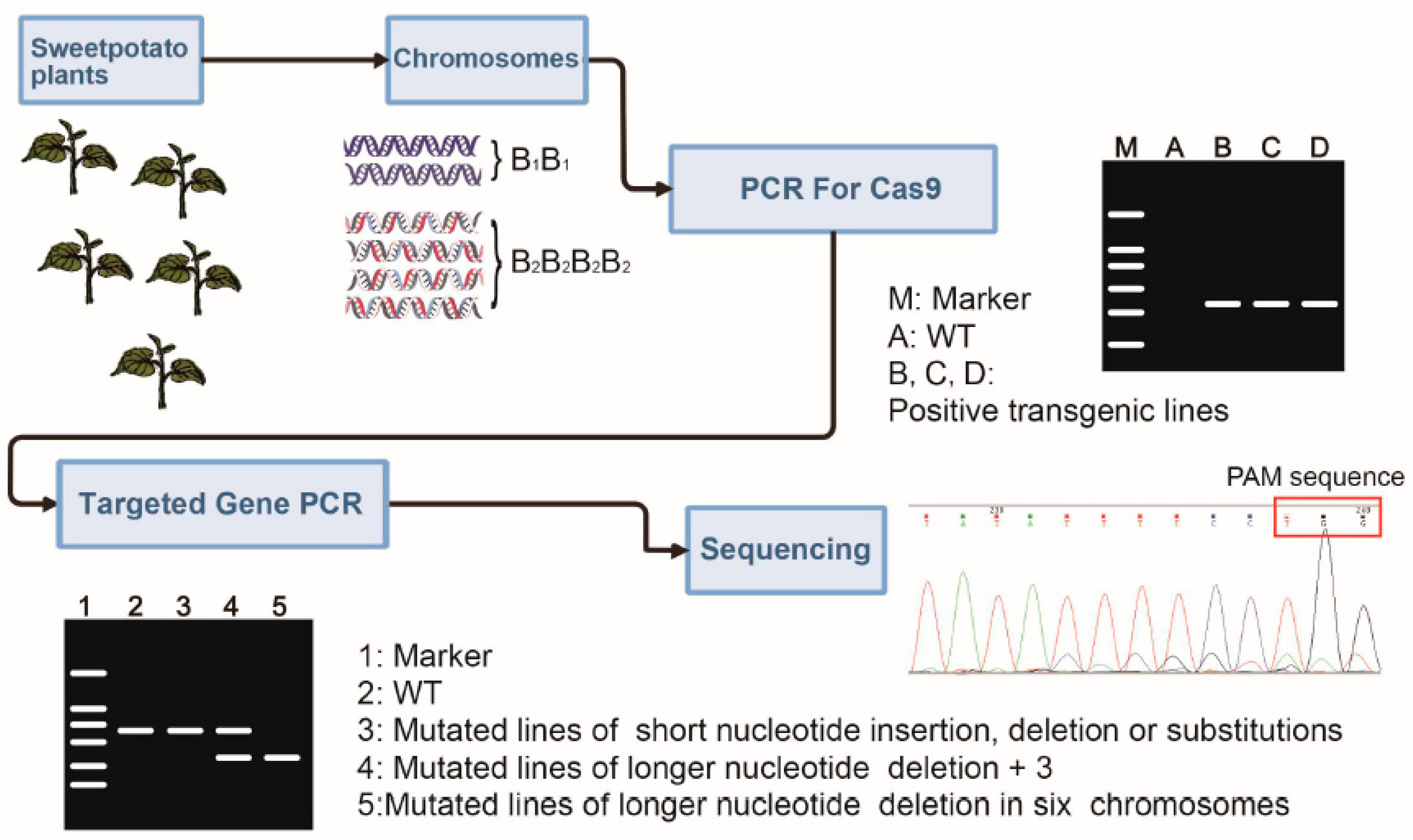
4.3. Detection of the Mutation of Target Genes
4.4. Detection of Total Starch and Amylose Content in the Mutants
4.5. Detection of Chain Length Distribution
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jain, A.; Zode, G.; Kasetti, R.B.; Ran, F.A.; Yan, W.; Sharma, T.P.; Bugge, K.; Searby, C.C.; Fingert, J.H.; Zhang, F.; et al. CRISPR-Cas9-based treatment of myocilin-associated glaucoma. Proc. Natl. Acad. Sci. USA 2017, 114, 11199–11204. [Google Scholar] [CrossRef]
- Yin, K.; Gao, C.; Qiu, J.L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Arazoe, T.; Yachie, N.; Banno, S.; Kakimoto, M.; Tabata, M.; Mochizuki, M.; Miyabe, A.; Araki, M.; Hara, K.Y.; et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science 2016, 353, 6305. [Google Scholar] [CrossRef]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. CRISPR for Crop Improvement: An Update Review. Front Plant Sci. 2018, 9, 985. [Google Scholar] [CrossRef]
- Ricroch, A.; Clairand, P.; Harwood, W. Use of CRISPR systems in plant genome editing: Toward new opportunities in agriculture. Emerg. Top. Life Sci. 2017, 1, 169–182. [Google Scholar] [CrossRef]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 232. [Google Scholar] [CrossRef]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.; Yin, X.; Biswal, A.; Coe, R.; Quick, W.P. CRISPR-Cas9-Mediated Genome Editing of Rice Towards Better Grain Quality. Rice Grain Qual. 2018, 1892, 311–336. [Google Scholar]
- Tuncel, A.; Corbin, K.R.; Ahn-Jarvis, J.; Harris, S.; Hawkins, E.; Smedley, M.A.; Harwood, W.; Warren, F.J.; Patron, N.J.; Smith, A.M. Cas9-mediated mutagenesis of potato starch-branching enzymes generates a range of tuber starch phenotypes. Plant Biotechnol. J. 2019. [Google Scholar] [CrossRef]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An Effective Strategy for Reliably Isolating Heritable and Cas9-Free Arabidopsis Mutants Generated by CRISPR/Cas9-Mediated Genome Editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef]
- He, Y.; Zhu, M.; Wang, L.; Wu, J.; Wang, Q.; Wang, R.; Zhao, Y. Programmed self-elimination of the CRISPR/Cas9 construct greatly accelerates the isolation of edited and transgene-free rice plants. Mol. Plant 2018, 11, 2010–2013. [Google Scholar] [CrossRef]
- Oke, M.O.; Workneh, T.S. A review on sweet potato postharvest processing and preservation technology. Int. J. Agric. Res. Rev. 2013, 1, 001–014. [Google Scholar]
- James, M.G.; Robertson, D.S.; Myers, A.M. Characterization of the maize gene sugary1, a determinant of starch composition in kernels. Plant Cell 1995, 7, 417–429. [Google Scholar]
- Johnson, P.E.; Patron, N.J.; Bottrill, A.R.; Dinges, J.R.; Fahy, B.F.; Parker, M.L.; Waite, D.N.; Denyer, K. A low-starch barley mutant, Risø 16, lacking the cytosolic small subunit of ADP-glucose pyrophosphorylase, reveals the importance of the cytosolic isoform and the identity of the plastidial small subunit. Plant. Physiol. 2003, 131, 684–696. [Google Scholar] [CrossRef]
- Tetlow, I.J. Understanding storage starch biosynthesis in plants: A means to quality improvement. Can. J. Bot. 2006, 84, 1167–1185. [Google Scholar] [CrossRef]
- James, M.G.; Denyer, K.; Myers, A.M. Starch synthesis in the cereal endosperm. Curr. Opin. Plant Biol. 2003, 6, 215–222. [Google Scholar] [CrossRef]
- Kimura, T.; Otani, M.; Noda, T.; Ideta, O.; Shimada., T.; Saito, A. Absence of amylose in sweet potato [Ipomoea batatas (L.) Lam.] following the introduction of granules-bound starch synthase I cDNA. Plant Cell Rep. 2001, 20, 663–666. [Google Scholar] [CrossRef]
- Otani, M.; Hamada, T.; Katayama, K.; Kitahara, K.; Kim, S.H.; Takahata, Y.; Suganum, T. Inhibition of the gene expression for granule bound starch synthase I by RNA interference in sweet potato plants. Plant Cell Rep. 2007, 26, 1801–1807. [Google Scholar] [CrossRef]
- Kitahara, K.; Hamasuna, K.; Nozuma, K.; Otani, M.; Hamada, T.; Shimada, T.; Fujita, K.; Suganuma, T. Physicochemical properties of amylose-free and high-amylose starches from transgenic sweet potato modified by RNA interference. Carbohyd. Polym. 2007, 69, 233–240. [Google Scholar] [CrossRef]
- Shimada, T.; Otani, M.; Hamada, T.; Kim, S.H. Increase of amylose content of sweet potato starch by RNA interference of the starch branching enzyme II gene (IbSBEII). Plant Biotechnol. 2006, 23, 85–90. [Google Scholar] [CrossRef]
- Rydberg, U.; Andersson, L.; Andersson, R.; Aman, P.; Larsson, H. Comparison of starch branching enzyme I and II from potato. Eur. J. Biochem. 2001, 268, 6140–6145. [Google Scholar] [CrossRef]
- Yang, J.; Bi, H.P.; Fan, W.J.; Zhang, M.; Wang, H.X.; Zhang, P. Efficient embryogenic suspension culturing and rapid transformation of a range of elite genotypes of sweet potato (Ipomoea batatas [L.] Lam.). Plant. Sci. 2011, 181, 701–711. [Google Scholar] [CrossRef]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef]
- Wu, S.; Lau, K.H.; Cao, Q.; Hamilton, J.P.; Sun, H.; Zhou, C.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.; et al. Genome sequences of two diploid wild relatives of cultivated sweet potato reveal targets for genetic improvement. Nat. Commun. 2018, 9, 4580. [Google Scholar] [CrossRef]
- Ukoskit, K.; Thompson, P.G. Autopolyploidy versus allopolyoloidy and low-density randomly amplified polymorphic DNA linkage maps of sweet potato. J. Am. Soc. Hortic. Sci. 1997, 122, 822–828. [Google Scholar] [CrossRef]
- Sanchez-Leon, S.; Gil-Humanes, J.; Ozuna, C.V.; Gimenez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant. Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Zhang, Y.; Liu, J.; Yin, K.; Qiu, J.L.; Gao, C. Genome editing of bread wheat using biolistic delivery of CRISPR/Cas9 in vitro transcripts or ribonucleoproteins. Nat. Protoc. 2018, 13, 413–430. [Google Scholar] [CrossRef]
- Li, C.; Unver, T.; Zhang, B. A high-efficiency CRISPR/Cas9 system for targeted mutagenesis in Cotton (Gossypium hirsutum L.). Sci. Rep. 2017, 7, 43902. [Google Scholar] [CrossRef]
- Janga, M.R.; Campbell, L.M.; Rathore, K.S. CRISPR/Cas9-mediated targeted mutagenesis in upland cotton (Gossypium hirsutum L.). Plant. Mol. Biol. 2017, 94, 349–360. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Nicolia, A.; Falt, A.S.; Samuelsson, M.; Hofvander, P. Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas9 expression in protoplasts. Plant. Cell Rep. 2017, 36, 117–128. [Google Scholar] [CrossRef]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant. Physiol. Biochem. 2018, 131, 63–69. [Google Scholar] [CrossRef]
- Sander, J.D.; Joung, J.K. CRISPR-Cas systems for genome editing, regulation and targeting. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef]
- He, Y.; Wang, R.; Dai, X.; Zhao, Y. On Improving CRISPR for Editing Plant Genes: Ribozyme-Mediated Guide RNA Production and Fluorescence-Based Technology for Isolating Transgene-Free Mutants Generated by CRISPR. Prog. Mol. Biol. Transl. Sci. 2017, 149, 151–166. [Google Scholar]
- Zhu, F.; Wang, S. Physicochemical properties, molecular structure, and uses of sweet potato starch. Trends Food sci. Tech. 2014, 36, 68–78. [Google Scholar] [CrossRef]
- Hebelstrup, K.H.; Nielsen, M.M.; Carciofi, M.; Andrzejczak, O.; Shaik, S.S.; Andreas, B.; Palcic, M.M. Waxy and non-waxy barley cultivars exhibit differences in the targeting and catalytic activity of GBSS1a. J. Exp. Bot. 2017, 68, 931–941. [Google Scholar] [CrossRef]
- Wang, S.; Li, C.; Copeland, L.; Niu, Q.; Wang, S. Starch Retrogradation: A Comprehensive Review. Food Sci. Food Saf. 2015, 14, 568–585. [Google Scholar] [CrossRef]
- Zhao, S.S.; Dufour, D.; Sanchez, T.; Ceballos, H.; Zhang, P. Development of Waxy Cassava With Different Biological and Physico-Chemical Characteristics of Starches for Industrial Applications. Biotechnol. Bioeng. 2011, 108, 1925–1935. [Google Scholar] [CrossRef]
- Rai, S.; Kaur, A.; Chopra, C.S. Gluten-Free Products for Celiac Susceptible People. Front. Nutr. 2018, 5, 116. [Google Scholar] [CrossRef]
- Raigond, P.; Ezekiel, R. Raigond B Resistant starch in food: A review. J. Sci. Food Agr. 2015, 95, 1968–7198. [Google Scholar] [CrossRef]
- Bindels, L.B.; Segura Munoz, R.R.; Gomes-Neto, J.C.; Mutemberezi, V.; Martínez, I.; Salazar, N.; Cody, E.A.; Quintero-Villegas, M.I.; Kittana, H.; de Los Reyes-Gavilán, C.G.; et al. Resistant starch can improve insulin sensitivity independently of the gut microbiota. Microbiota 2017, 5, 12. [Google Scholar] [CrossRef]
- Emilien, C.H.; Hsu, W.H.; Hollis, J.H. Effect of resistant wheat starch on subjective appetite and food intake in healthy adults. Nutrition 2017, 43, 69–74. [Google Scholar] [CrossRef]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; Martin, R.J.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol Ren. Physiol. 2016, 310, 857–871. [Google Scholar] [CrossRef]
- Jobling, S. Improving starch for food and industrial applications. Curr. Opin. Plant. Biol. 2004, 7, 210–218. [Google Scholar] [CrossRef]
- Wang, H.X.; Yang, J.; Zhang, M.; Fan, W.J.; Nurit, F.; Sitakanta, P.; Yuan, L.; Zhang, P. Altered phenylpropanoid metabolism in the Maize Lc-expressed sweet potato (Ipomoea batatas) affects storage root development. Sci. Rep. 2016, 6, 18645. [Google Scholar] [CrossRef]
- Zhou, W.Z.; Yang, J.; Hong, Y.; Liu, G.l.; Zheng, J.l.; Gu, Z.B.; Zhang, P. Impact of amylose content on starch physicochemical properties in transgenic sweet potato. Carbohydr. Polym. 2015, 122, 417–427. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, W.Z.; Qiao, D.; Peng, Z.; Zhao, S.; Zhang, L.; Xie, F. Changes in Nanoscale Chain Assembly in Sweet Potato Starch Lamellae by Downregulation of Biosynthesis Enzymes. J. Agric. Food Chem. 2019, 67, 6302–6312. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Target Gene | No. of Plants with Cas9 Gene | No. of Plants with Mutation | Mutation Rates of Transgenic Plants |
|---|---|---|---|---|
| Xushu22 | IbGBSSI | 25 | 23 | 92.0% |
| Xushu22 | IbSBEII | 47 | 40 | 85.1% |
| Taizhong6 | IbGBSSI | 24 | 15 | 62.5% |
| Taizhong6 | IbSBEII | 11 | 7 | 63.6% |
| Sample | 6 < DP < 12 (%) | 13 < DP < 24 (%) | 25 < DP < 36 (%) | 37 < DP < 90 (%) |
|---|---|---|---|---|
| Xushu22 | 29.2 d (0.42) | 42.7 c (0.752) | 14.7 c (0.40) | 13.4 c (0.35) |
| IbSBEII-sgRNA12-24 | 24.6 e (0.63) | 44.6 b (0.53) | 15.0 c (0.16) | 15.8 b (0.78) |
| IbSBEII-sgRNA12-26 | 16.4 g (0.03) | 45.6 ab (0.06) | 19.2 a (0.04) | 18.8 a (0.05) |
| IbSBEII-sgRNA2-1 | 19.1 f (0.25) | 46.3 a (0.34) | 16.3 b (0.04) | 18.3 a (0.62) |
| IbGBSSI-sgRNA2-2 | 33.7 b (0.07) | 41.7 cd (0.27) | 13.8 d (0.13) | 11.2 d (0.23) |
| IbGBSSI-sgRNA2-6 | 34.7 a (0.21) | 41.4 d (0.46) | 13.1 e (0.13) | 10.8 d (0.31) |
| IbGBSSI-sgRNA2-7 | 30.4 c (0.08) | 42.6 c (0.10) | 13.6 d (0.23) | 13.4 c (0.23) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wu, Y.; Zhang, Y.; Yang, J.; Fan, W.; Zhang, H.; Zhao, S.; Yuan, L.; Zhang, P. CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality. Int. J. Mol. Sci. 2019, 20, 4702. https://doi.org/10.3390/ijms20194702
Wang H, Wu Y, Zhang Y, Yang J, Fan W, Zhang H, Zhao S, Yuan L, Zhang P. CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality. International Journal of Molecular Sciences. 2019; 20(19):4702. https://doi.org/10.3390/ijms20194702
Chicago/Turabian StyleWang, Hongxia, Yinliang Wu, Yandi Zhang, Jun Yang, Weijuan Fan, Hui Zhang, Shanshan Zhao, Ling Yuan, and Peng Zhang. 2019. "CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality" International Journal of Molecular Sciences 20, no. 19: 4702. https://doi.org/10.3390/ijms20194702
APA StyleWang, H., Wu, Y., Zhang, Y., Yang, J., Fan, W., Zhang, H., Zhao, S., Yuan, L., & Zhang, P. (2019). CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality. International Journal of Molecular Sciences, 20(19), 4702. https://doi.org/10.3390/ijms20194702