Advances Towards How Meiotic Recombination Is Initiated: A Comparative View and Perspectives for Plant Meiosis Research


Abstract
:1. Introduction
2. Conserved Spo11 and Non-Conserved Spo11 Accessary Proteins in Different Species
3. Defining Meiotic DSB Hotspots in Different Species
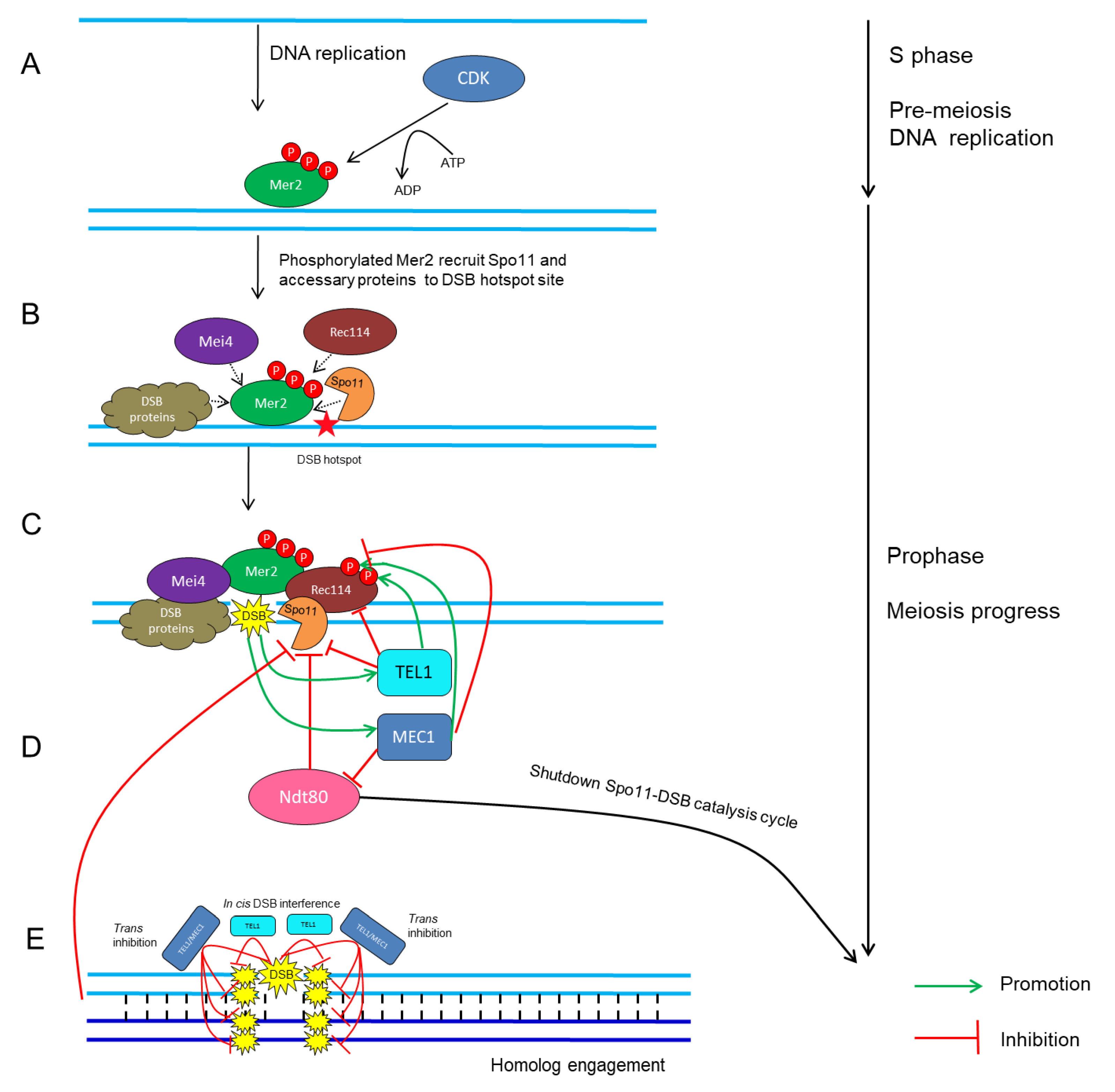
4. Control of Meiotic DSB Formation by Protein Phosphorylation
4.1. Cyclin-Dependent Kinases (CDKs)
4.2. Tel1/ATM and Mec1/ATR
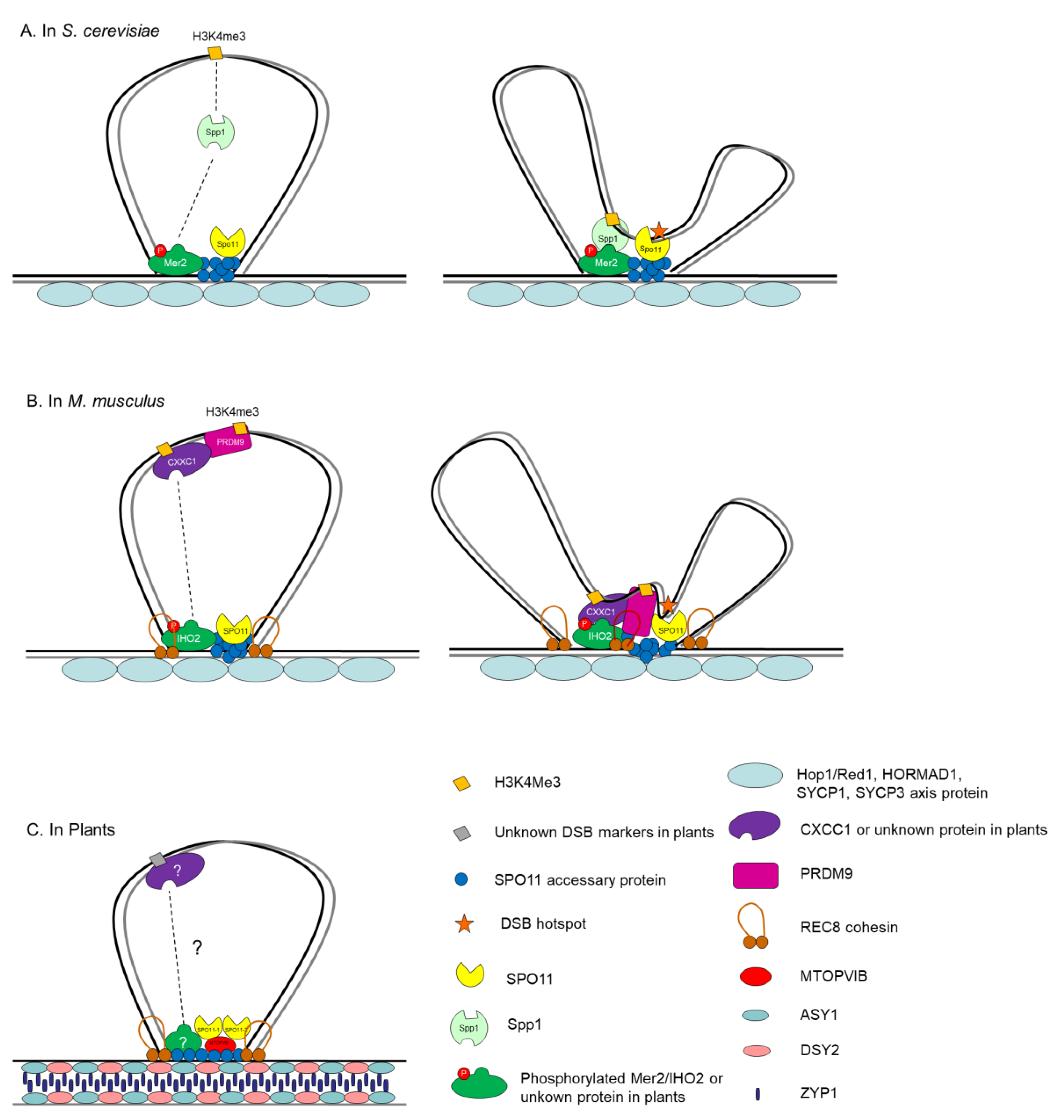
5. Control of Meiosis DSB Formation in the Context of ‘Tethered Loop-Axis Complex’
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feng, X.; Zilberman, D.; Dickinson, H. A conversation across generations: Soma-germ cell crosstalk in plants. Dev. Cell 2013, 24, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K. Be my baby: Patterning toward plant germ cells. Curr. Opin. Plant Biol. 2018, 41, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.T.; Colaiácovo, M.P. Regulation of Crossover Frequency and Distribution during Meiotic Recombination. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2018; Volume 82, pp. 223–234. [Google Scholar] [CrossRef]
- Lam, I.; Keeney, S. Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016634. [Google Scholar] [CrossRef] [PubMed]
- Mercier, R.; Mezard, C.; Jenczewski, E.; Macaisne, N.; Grelon, M. The molecular biology of meiosis in plants. Annu. Rev. Plant Biol. 2015, 66, 297–327. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S.; Lange, J.; Mohibullah, N. Self-organization of meiotic recombination initiation: General principles and molecular pathways. Annu. Rev. Genet. 2014, 48, 187–214. [Google Scholar] [CrossRef] [PubMed]
- Lambing, C.; Heckmann, S. Tackling Plant Meiosis: From Model Research to Crop Improvement. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Edlinger, B.; Schlogelhofer, P. Have a break: Determinants of meiotic DNA double strand break (DSB) formation and processing in plants. J. Exp. Bot. 2011, 62, 1545–1563. [Google Scholar] [CrossRef]
- Wang, Y.; Copenhaver, G.P. Meiotic Recombination: Mixing It Up in Plants. Annu. Rev. Plant Biol. 2018. [Google Scholar] [CrossRef]
- Zelkowski, M.; Olson, M.A.; Wang, M.; Pawlowski, W. Diversity and Determinants of Meiotic Recombination Landscapes. Trends Genet. TIG 2019. [Google Scholar] [CrossRef]
- Hartung, F.; Wurz-Wildersinn, R.; Fuchs, J.; Schubert, I.; Suer, S.; Puchta, H. The catalytically active tyrosine residues of both SPO11-1 and SPO11-2 are required for meiotic double-strand break induction in Arabidopsis. Plant Cell 2007, 19, 3090–3099. [Google Scholar] [CrossRef]
- De Muyt, A.; Vezon, D.; Gendrot, G.; Gallois, J.L.; Stevens, R.; Grelon, M. AtPRD1 is required for meiotic double strand break formation in Arabidopsis thaliana. EMBO J. 2007, 26, 4126–4137. [Google Scholar] [CrossRef] [PubMed]
- Vrielynck, N.; Chambon, A.; Vezon, D.; Pereira, L.; Chelysheva, L.; De Muyt, A.; Mezard, C.; Mayer, C.; Grelon, M. A DNA topoisomerase VI-like complex initiates meiotic recombination. Science 2016, 351, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, W.M.; Altun, C.; Armstrong, S.J.; Roberts, N.; Dean, P.J.; Young, K.; Weil, C.F.; Bray, C.M.; West, C.E. NBS1 is involved in DNA repair and plays a synergistic role with ATM in mediating meiotic homologous recombination in plants. Plant J. 2007, 52, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Bleuyard, J.Y.; Gallego, M.E.; White, C.I. Meiotic defects in the Arabidopsis rad50 mutant point to conservation of the MRX complex function in early stages of meiotic recombination. Chromosoma 2004, 113, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Puizina, J.; Siroky, J.; Mokros, P.; Schweizer, D.; Riha, K. Mre11 deficiency in Arabidopsis is associated with chromosomal instability in somatic cells and Spo11-dependent genome fragmentation during meiosis. Plant Cell 2004, 16, 1968–1978. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Tang, D.; Wang, K.; Wang, M.; Che, L.; Li, M.; Cheng, Z. The role of OsCOM1 in homologous chromosome synapsis and recombination in rice meiosis. Plant J. 2012, 72, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, L.; Zhang, T.; Jing, J.; He, Y. ZmCom1 Is Required for Both Mitotic and Meiotic Recombination in Maize. Front. Plant Sci. 2018, 9, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uanschou, C.; Siwiec, T.; Pedrosa-Harand, A.; Kerzendorfer, C.; Sanchez-Moran, E.; Novatchkova, M.; Akimcheva, S.; Woglar, A.; Klein, F.; Schlogelhofer, P. A novel plant gene essential for meiosis is related to the human CtIP and the yeast COM1/SAE2 gene. EMBO J. 2007, 26, 5061–5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Treco, D.; Szostak, J.W. Extensive 3′-overhanging, single-stranded DNA associated with the meiosis-specific double-strand breaks at the ARG4 recombination initiation site. Cell 1991, 64, 1155–1161. [Google Scholar] [CrossRef]
- Zakharyevich, K.; Ma, Y.; Tang, S.; Hwang, P.Y.; Boiteux, S.; Hunter, N. Temporally and biochemically distinct activities of Exo1 during meiosis: Double-strand break resection and resolution of double Holliday junctions. Mol. Cell 2010, 40, 1001–1015. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Alani, E.; Kleckner, N. A pathway for generation and processing of double-strand breaks during meiotic recombination in S. cerevisiae. Cell 1990, 61, 1089–1101. [Google Scholar] [CrossRef]
- Singh, G.; Da Ines, O.; Gallego, M.E.; White, C.I. Analysis of the impact of the absence of RAD51 strand exchange activity in Arabidopsis meiosis. PLoS ONE 2017, 12, e0183006. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Cheng, Z.; Huang, J.; Lin, J.; Copenhaver, G.P.; Ma, H.; Wang, Y. Arabidopsis RAD51, RAD51C and XRCC3 proteins form a complex and facilitate RAD51 localization on chromosomes for meiotic recombination. PLoS Genet. 2017, 13, e1006827. [Google Scholar] [CrossRef]
- Bleuyard, J.Y.; Gallego, M.E.; Savigny, F.; White, C.I. Differing requirements for the Arabidopsis Rad51 paralogs in meiosis and DNA repair. Plant J. 2005, 41, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Da Ines, O.; Degroote, F.; Goubely, C.; Amiard, S.; Gallego, M.E.; White, C.I. Meiotic recombination in Arabidopsis is catalysed by DMC1, with RAD51 playing a supporting role. PLoS Genet. 2013, 9, e1003787. [Google Scholar] [CrossRef] [PubMed]
- Kurzbauer, M.T.; Uanschou, C.; Chen, D.; Schlogelhofer, P. The recombinases DMC1 and RAD51 are functionally and spatially separated during meiosis in Arabidopsis. Plant Cell 2012, 24, 2058–2070. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Harper, L.C.; Golubovskaya, I.; Wang, C.R.; Weber, D.; Meeley, R.B.; McElver, J.; Bowen, B.; Cande, W.Z.; Schnable, P.S. Functional analysis of maize RAD51 in meiosis and double-strand break repair. Genetics 2007, 176, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, Q. OsDMC1 Is Not Required for Homologous Pairing in Rice Meiosis. Plant Physiol. 2016, 171, 230–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, D.; Miao, C.; Li, Y.; Wang, H.; Liu, X.; Yu, H.; Cheng, Z. OsRAD51C is essential for double-strand break repair in rice meiosis. Front. Plant Sci. 2014, 5, 167. [Google Scholar] [CrossRef] [Green Version]
- Szurman-Zubrzycka, M.; Baran, B.; Stolarek-Januszkiewicz, M.; Kwasniewska, J.; Szarejko, I.; Gruszka, D. The dmc1 Mutant Allows an Insight into the DNA Double-Strand Break Repair during Meiosis in Barley (Hordeum vulgare L.). Front. Plant Sci. 2019, 10, 761. [Google Scholar] [CrossRef]
- Kobayashi, W.; Liu, E.; Ishii, H.; Matsunaga, S.; Schlogelhofer, P.; Kurumizaka, H. Homologous pairing activities of Arabidopsis thaliana RAD51 and DMC1. J. Biochem. 2019, 165, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Pradillo, M.; Varas, J.; Oliver, C.; Santos, J.L. On the role of AtDMC1, AtRAD51 and its paralogs during Arabidopsis meiosis. Front. Plant Sci. 2014, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- San Filippo, J.; Sung, P.; Klein, H. Mechanism of eukaryotic homologous recombination. Annu. Rev. Biochem. 2008, 77, 229–257. [Google Scholar] [CrossRef] [PubMed]
- Lambing, C.; Franklin, F.C.H.; Wang, C.-J.R. Understanding and Manipulating Meiotic Recombination in Plants. Plant Physiol. 2017, 173, 1530–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Choi, K. Signaling-mediated meiotic recombination in plants. Curr. Opin. Plant Biol. 2019, 51, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Drouaud, J.; Khademian, H.; Giraut, L.; Zanni, V.; Bellalou, S.; Henderson, I.R.; Falque, M.; Mezard, C. Contrasted patterns of crossover and non-crossover at Arabidopsis thaliana meiotic recombination hotspots. PLoS Genet. 2013, 9, e1003922. [Google Scholar] [CrossRef] [PubMed]
- Dluzewska, J.; Szymanska, M.; Ziolkowski, P.A. Where to Cross Over? Defining Crossover Sites in Plants. Front. Genet. 2018, 9, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyt, A.; Mercier, R.; Mezard, C.; Grelon, M. Meiotic recombination and crossovers in plants. Genome Dyn. 2009, 5, 14–25. [Google Scholar] [CrossRef]
- Mezard, C.; Vignard, J.; Drouaud, J.; Mercier, R. The road to crossovers: Plants have their say. Trends Genet. TIG 2007, 23, 91–99. [Google Scholar] [CrossRef]
- Cifuentes, M.; Rivard, M.; Pereira, L.; Chelysheva, L.; Mercier, R. Haploid meiosis in Arabidopsis: Double-strand breaks are formed and repaired but without synapsis and crossovers. PLoS ONE 2013, 8, e72431. [Google Scholar] [CrossRef]
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef]
- Kumar, R.; Bourbon, H.M.; de Massy, B. Functional conservation of Mei4 for meiotic DNA double-strand break formation from yeasts to mice. Genes Dev. 2010, 24, 1266–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleki, S.; Neale, M.J.; Arora, C.; Henderson, K.A.; Keeney, S. Interactions between Mei4, Rec114, and other proteins required for meiotic DNA double-strand break formation in Saccharomyces cerevisiae. Chromosoma 2007, 116, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hooker, G.W.; Roeder, G.S. Saccharomyces cerevisiae Mer2, Mei4 and Rec114 Form a Complex Required for Meiotic Double-Strand Break Formation. Genetics 2006, 173, 1969. [Google Scholar] [CrossRef] [PubMed]
- Menees, T.M.; Roeder, G.S. MEI4, a yeast gene required for meiotic recombination. Genetics 1989, 123, 675. [Google Scholar] [PubMed]
- Arora, C.; Kee, K.; Maleki, S.; Keeney, S. Antiviral Protein Ski8 Is a Direct Partner of Spo11 in Meiotic DNA Break Formation, Independent of Its Cytoplasmic Role in RNA Metabolism. Mol. Cell 2004, 13, 549–559. [Google Scholar] [CrossRef]
- Salem, L.; Walter, N.; Malone, R. Suppressor Analysis of the Saccharomyces cerevisiae Gene REC104 Reveals a Genetic Interaction With REC102. Genetics 1999, 151, 1261. [Google Scholar]
- Kee, K.; Protacio, R.U.; Arora, C.; Keeney, S. Spatial organization and dynamics of the association of Rec102 and Rec104 with meiotic chromosomes. EMBO J. 2004, 23, 1815–1824. [Google Scholar] [CrossRef]
- Borde, V.; Lin, W.; Novikov, E.; Petrini, J.H.; Lichten, M.; Nicolas, A. Association of Mre11p with Double-Strand Break Sites during Yeast Meiosis. Mol. Cell 2004, 13, 389–401. [Google Scholar] [CrossRef]
- Johzuka, K.; Ogawa, H. Interaction of Mre11 and Rad50: Two proteins required for DNA repair and meiosis-specific double-strand break formation in Saccharomyces cerevisiae. Genetics 1995, 139, 1521–1532. [Google Scholar]
- Ogawa, H.; Johzuka, K.; Nakagawa, T.; Leem, S.-H.; Hagihara, A.H. Functions of the yeast meiotic recombination genes, MRE11 and MRE2. Adv. Biophys. 1995, 31, 67–76. [Google Scholar] [CrossRef]
- Steiner, S.; Kohli, J.; Ludin, K. Functional interactions among members of the meiotic initiation complex in fission yeast. Curr. Genet. 2010, 56, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Smith, G.R. Transient, meiosis-induced expression of the rec6 and rec12 genes of Schizosaccharomyces pombe. Genetics 1994, 136, 769–779. [Google Scholar] [PubMed]
- Miyoshi, T.; Ito, M.; Kugou, K.; Yamada, T.; Yamada, S.; Furuichi, M.; Oda, A.; Hirota, K.; Masai, H.; Ohta, K. A Central Coupler for Recombination Initiation Linking Chromosome Architecture to S Phase Checkpoint. Mol. Cell 2012, 47, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfils, S.; Rozalen, A.E.; Smith, G.R.; Moreno, S.; Martin-Castellanos, C. Functional interactions of Rec24, the fission yeast ortholog of mouse Mei4, with the meiotic recombination-initiation complex. J. Cell Sci. 2011, 124, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Martin-Castellanos, C.; Blanco, M.; Rozalen, A.E.; Perez-Hidalgo, L.; Garcia, A.I.; Conde, F.; Mata, J.; Ellermeier, C.; Davis, L.; San-Segundo, P.; et al. A large-scale screen in S. pombe identifies seven novel genes required for critical meiotic events. Curr. Biol. 2005, 15, 2056–2062. [Google Scholar] [CrossRef] [PubMed]
- Gregan, J.; Rabitsch, P.K.; Sakem, B.; Csutak, O.; Latypov, V.; Lehmann, E.; Kohli, J.; Nasmyth, K. Novel genes required for meiotic chromosome segregation are identified by a high-throughput knockout screen in fission yeast. Curr. Biol. 2005, 15, 1663–1669. [Google Scholar] [CrossRef]
- Lorenz, A.; Wells, J.L.; Pryce, D.W.; Novatchkova, M.; Eisenhaber, F.; McFarlane, R.J.; Loidl, J. S. pombe meiotic linear elements contain proteins related to synaptonemal complex components. J. Cell Sci. 2004, 117, 3343–3351. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, A.; Estreicher, A.; Kohli, J.; Loidl, J. Meiotic recombination proteins localize to linear elements in Schizosaccharomyces pombe. Chromosoma 2006, 115, 330–340. [Google Scholar] [CrossRef]
- Young, J.A.; Hyppa, R.W.; Smith, G.R. Conserved and nonconserved proteins for meiotic DNA breakage and repair in yeasts. Genetics 2004, 167, 593–605. [Google Scholar] [CrossRef]
- Robert, T.; Vrielynck, N.; Mezard, C.; de Massy, B.; Grelon, M. A new light on the meiotic DSB catalytic complex. Semin. Cell Dev. Biol. 2016, 54, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Stanzione, M.; Baumann, M.; Papanikos, F.; Dereli, I.; Lange, J.; Ramlal, A.; Tränkner, D.; Shibuya, H.; de Massy, B.; Watanabe, Y.; et al. Meiotic DNA break formation requires the unsynapsed chromosome axis-binding protein IHO1 (CCDC36) in mice. Nat. Cell Biol. 2016, 18, 1208–1220. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Ghyselinck, N.; Ishiguro, K.; Watanabe, Y.; Kouznetsova, A.; Hoog, C.; Strong, E.; Schimenti, J.; Daniel, K.; Toth, A.; et al. MEI4—A central player in the regulation of meiotic DNA double-strand break formation in the mouse. J. Cell Sci. 2015, 128, 1800–1811. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Oliver, C.; Brun, C.; Juarez-Martinez, A.B.; Tarabay, Y.; Kadlec, J.; de Massy, B. Mouse REC114 is essential for meiotic DNA double-strand break formation and forms a complex with MEI4. Life Sci. Alliance 2018, 1, e201800259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panizza, S.; Mendoza, M.A.; Berlinger, M.; Huang, L.; Nicolas, A.; Shirahige, K.; Klein, F. Spo11-Accessory Proteins Link Double-Strand Break Sites to the Chromosome Axis in Early Meiotic Recombination. Cell 2011, 146, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libby, B.J.; Reinholdt, L.G.; Schimenti, J.C. Positional cloning and characterization of Mei1, a vertebrate-specific gene required for normal meiotic chromosome synapsis in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 15706–15711. [Google Scholar] [CrossRef] [PubMed]
- Libby, B.J.; De La Fuente, R.; O’Brien, M.J.; Wigglesworth, K.; Cobb, J.; Inselman, A.; Eaker, S.; Handel, M.A.; Eppig, J.J.; Schimenti, J.C. The mouse meiotic mutation mei1 disrupts chromosome synapsis with sexually dimorphic consequences for meiotic progression. Dev. Biol. 2002, 242, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; De Massy, B. Initiation of meiotic recombination in mammals. Genes 2010, 1, 521–549. [Google Scholar] [CrossRef]
- Robert, T.; Nore, A.; Brun, C.; Maffre, C.; Crimi, B.; Bourbon, H.M.; de Massy, B. The TopoVIB-Like protein family is required for meiotic DNA double-strand break formation. Science 2016, 351, 943–949. [Google Scholar] [CrossRef]
- Stacey, N.J.; Kuromori, T.; Azumi, Y.; Roberts, G.; Breuer, C.; Wada, T.; Maxwell, A.; Roberts, K.; Sugimoto-Shirasu, K. Arabidopsis SPO11-2 functions with SPO11-1 in meiotic recombination. Plant J. 2006, 48, 206–216. [Google Scholar] [CrossRef]
- Grelon, M.; Vezon, D.; Gendrot, G.; Pelletier, G. AtSPO11-1 is necessary for efficient meiotic recombination in plants. EMBO J. 2001, 20, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shingu, Y.; Tokai, T.; Agawa, Y.; Toyota, K.; Ahamed, S.; Kawagishi-Kobayashi, M.; Komatsu, A.; Mikawa, T.; Yamamoto, M.T.; Wakasa, K.; et al. The double-stranded break-forming activity of plant SPO11s and a novel rice SPO11 revealed by a Drosophila bioassay. BMC Mol. Biol. 2012, 13, 1. [Google Scholar] [CrossRef]
- Hartung, F.; Puchta, H. Molecular characterisation of two paralogous SPO11 homologues in Arabidopsis thaliana. Nucleic Acids Res. 2000, 28, 1548–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprink, T.; Hartung, F. The splicing fate of plant SPO11 genes. Front. Plant Sci. 2014, 5, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartung, F.; Angelis, K.J.; Meister, A.; Schubert, I.; Melzer, M.; Puchta, H. An Archaebacterial Topoisomerase Homolog Not Present in Other Eukaryotes Is Indispensable for Cell Proliferation of Plants. Curr. Biol. 2002, 12, 1787–1791. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Yin, Z.; Zeng, Y.; Zhang, Q.; Chen, L.; He, Y.; Lu, P.; Ye, D.; Zhang, X. MTOPVIB interacts with AtPRD1 and plays important roles in formation of meiotic DNA double-strand breaks in Arabidopsis. Sci. Rep. 2017, 7, 10007. [Google Scholar] [CrossRef] [PubMed]
- De Muyt, A.; Pereira, L.; Vezon, D.; Chelysheva, L.; Gendrot, G.; Chambon, A.; Laine-Choinard, S.; Pelletier, G.; Mercier, R.; Nogue, F.; et al. A high throughput genetic screen identifies new early meiotic recombination functions in Arabidopsis thaliana. PLoS Genet. 2009, 5, e1000654. [Google Scholar] [CrossRef] [PubMed]
- Capilla-Perez, L.; Solier, V.; Portemer, V.; Chambon, A.; Hurel, A.; Guillebaux, A.; Vezon, D.; Cromer, L.; Grelon, M.; Mercier, R. The HEM Lines: A New Library of Homozygous Arabidopsis thaliana EMS Mutants and its Potential to Detect Meiotic Phenotypes. Front. Plant Sci. 2018, 9, 1339. [Google Scholar] [CrossRef] [PubMed]
- Cole, F.; Keeney, S.; Jasin, M. Evolutionary conservation of meiotic DSB proteins: More than just Spo11. Genes Dev. 2010, 24, 1201–1207. [Google Scholar] [CrossRef]
- Tesse, S.; Bourbon, H.M.; Debuchy, R.; Budin, K.; Dubois, E.; Liangran, Z.; Antoine, R.; Piolot, T.; Kleckner, N.; Zickler, D.; et al. Asy2/Mer2: An evolutionarily conserved mediator of meiotic recombination, pairing, and global chromosome compaction. Genes Dev. 2017, 31, 1880–1893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Song, Y.; Cheng, Z.H.; Wang, Y.X.; Zhu, J.; Ma, H.; Xu, L.; Yang, Z.N. The Arabidopsis thaliana DSB formation (AtDFO) gene is required for meiotic double-strand break formation. Plant J. 2012, 72, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Li, Y.; Zhang, L.; Shi, W.; Zhang, C.; Feng, M.; Zhang, F.; Tang, D.; Yu, H.; Gu, M.; et al. OsMTOPVIB Promotes Meiotic DNA Double-Strand Break Formation in Rice. Mol. Plant 2016, 9, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- An, X.J.; Deng, Z.Y.; Wang, T. OsSpo11-4, a rice homologue of the archaeal TopVIA protein, mediates double-strand DNA cleavage and interacts with OsTopVIB. PLoS ONE 2011, 6, e20327. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, M.; Tang, D.; Wang, K.; Chen, F.; Gong, Z.; Gu, M.; Cheng, Z. OsSPO11-1 is essential for both homologous chromosome pairing and crossover formation in rice. Chromosoma 2010, 119, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Wang, C.; Xue, F.; Higgins, J.; Chen, M.; Zhang, D.; Liang, W. The DNA Topoisomerase VI-B Subunit OsMTOPVIB Is Essential for Meiotic Recombination Initiation in Rice. Mol. Plant 2016, 9, 1539–1541. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ji, J.; Tang, D.; Wang, H.; Shen, Y.; Shi, W.; Li, Y.; Tan, X.; Cheng, Z.; Luo, Q. OsSDS is essential for DSB formation in rice meiosis. Front. Plant Sci. 2015, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Tang, D.; Zhang, H.; Wang, M.; Li, Y.; Tang, S.; Yu, H.; Gu, M.; Cheng, Z. Central region component1, a novel synaptonemal complex component, is essential for meiotic recombination initiation in rice. Plant Cell 2013, 25, 2998–3009. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, B.; Tomchick, D.R.; Machius, M.; Rizo, J.; Yu, H.; Luo, X. p31comet blocks Mad2 activation through structural mimicry. Cell 2007, 131, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Han, J.; Chen, Y.; Wang, Y.; Liu, Y.G. Bivalent Formation 1, a plant-conserved gene, encodes an OmpH/coiled-coil motif-containing protein required for meiotic recombination in rice. J. Exp. Bot. 2017, 68, 2163–2174. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Tang, D.; Shen, Y.; Xue, Z.; Wang, H.; Shi, W.; Zhang, C.; Du, G.; Li, Y.; Cheng, Z. P31comet, a member of the synaptonemal complex, participates in meiotic DSB formation in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 10577–10582. [Google Scholar] [CrossRef]
- Pan, J.; Sasaki, M.; Kniewel, R.; Murakami, H.; Blitzblau, H.G.; Tischfield, S.E.; Zhu, X.; Neale, M.J.; Jasin, M.; Socci, N.D.; et al. A hierarchical combination of factors shapes the genome-wide topography of yeast meiotic recombination initiation. Cell 2011, 144, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, A.; Treco, D.; Schultes, N.P.; Szostak, J.W. An initiation site for meiotic gene conversion in the yeast Saccharomyces cerevisiae. Nature 1989, 338, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Baudat, F.; Nicolas, A. Clustering of meiotic double-strand breaks on yeast chromosome III. Proc. Natl. Acad. Sci. USA 1997, 94, 5213. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.; Gray, S.; Allison, R.M.; Cooper, T.J.; Neale, M.J. Tel1(ATM)-mediated interference suppresses clustered meiotic double-strand-break formation. Nature 2015, 520, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Richardson, L.L.; Jasin, M.; Handel, M.A.; Arnheim, N. Mouse strains with an active H2-Ea meiotic recombination hot spot exhibit increased levels of H2-Ea-specific DNA breaks in testicular germ cells. Mol. Cell. Biol. 2004, 24, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- De Massy, B.; Nicolas, A. The control in cis of the position and the amount of the ARG4 meiotic double-strand break of Saccharomyces cerevisiae. EMBO J. 1993, 12, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Henderson, I.R. Meiotic recombination hotspots—A comparative view. Plant J. 2015, 83, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Zhao, X.; Tock, A.J.; Lambing, C.; Underwood, C.J.; Hardcastle, T.J.; Serra, H.; Kim, J.; Cho, H.S.; Kim, J.; et al. Nucleosomes and DNA methylation shape meiotic DSB frequency in Arabidopsis thaliana transposons and gene regulatory regions. Genome Res. 2018, 28, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Lange, J.; Yamada, S.; Tischfield, S.E.; Pan, J.; Kim, S.; Zhu, X.; Socci, N.D.; Jasin, M.; Keeney, S. The Landscape of Mouse Meiotic Double-Strand Break Formation, Processing, and Repair. Cell 2016, 167, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Thacker, D.; Mohibullah, N.; Zhu, X.; Keeney, S. Homologue engagement controls meiotic DNA break number and distribution. Nature 2014, 510, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Underwood, C.J.; Choi, K.; Lambing, C.; Zhao, X.; Serra, H.; Borges, F.; Simorowski, J.; Ernst, E.; Jacob, Y.; Henderson, I.R.; et al. Epigenetic activation of meiotic recombination near Arabidopsis thaliana centromeres via loss of H3K9me2 and non-CG DNA methylation. Genome Res. 2018, 28, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Smagulova, F.; Gregoretti, I.V.; Brick, K.; Khil, P.; Camerini-Otero, R.D.; Petukhova, G.V. Genome-wide analysis reveals novel molecular features of mouse recombination hotspots. Nature 2011, 472, 375–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brick, K.; Smagulova, F.; Khil, P.; Camerini-Otero, R.D.; Petukhova, G.V. Genetic recombination is directed away from functional genomic elements in mice. Nature 2012, 485, 642–645. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, M.; Dukowic-Schulze, S.; Zhou, A.; Tiang, C.L.; Shilo, S.; Sidhu, G.K.; Eichten, S.; Bradbury, P.; Springer, N.M.; et al. Genomic features shaping the landscape of meiotic double-strand-break hotspots in maize. Proc. Natl. Acad. Sci. USA 2017, 114, 12231–12236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tock, A.J.; Henderson, I.R. Hotspots for Initiation of Meiotic Recombination. Front. Genet. 2018, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Kim, S.; Tischfield, S.E.; Jasin, M.; Lange, J.; Keeney, S. Genomic and chromatin features shaping meiotic double-strand break formation and repair in mice. Cell Cycle 2017, 16, 1870–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vader, G.; Blitzblau, H.G.; Tame, M.A.; Falk, J.E.; Curtin, L.; Hochwagen, A. Protection of repetitive DNA borders from self-induced meiotic instability. Nature 2011, 477, 115–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, M.; Tischfield, S.E.; van Overbeek, M.; Keeney, S. Meiotic Recombination Initiation in and around Retrotransposable Elements in Saccharomyces cerevisiae. PLoS Genet. 2013, 9, e1003732. [Google Scholar] [CrossRef]
- Zhu, X.; Keeney, S. High-Resolution Global Analysis of the Influences of Bas1 and Ino4 Transcription Factors on Meiotic DNA Break Distributions in Saccharomyces cerevisiae. Genetics 2015, 201, 525–542. [Google Scholar] [CrossRef]
- Mohibullah, N.; Keeney, S. Numerical and spatial patterning of yeast meiotic DNA breaks by Tel1. Genome Res. 2017, 27, 278–288. [Google Scholar] [CrossRef]
- Lam, I.; Keeney, S. Nonparadoxical evolutionary stability of the recombination initiation landscape in yeast. Science 2015, 350, 932–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, K.R.; Sasaki, M.; Milman, N.; Keeney, S.; Smith, G.R. Evolutionarily diverse determinants of meiotic DNA break and recombination landscapes across the genome. Genome Res. 2014, 24, 1650–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante-Jaramillo, L.F.; Ramos, C.; Alonso, L.; Sesmero, A.; Segurado, M.; Martin-Castellanos, C. CDK contribution to DSB formation and recombination in fission yeast meiosis. PLoS Genet. 2019, 15, e1007876. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Niu, H.; Futcher, B.; Zhang, C.; Shokat, K.M.; Boulton, S.J.; Hollingsworth, N.M. Cdc28-Clb5 (CDK-S) and Cdc7-Dbf4 (DDK) collaborate to initiate meiotic recombination in yeast. Genes Dev. 2008, 22, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, K.R. Control of landmark events in meiosis by the CDK Cdc28 and the meiosis-specific kinase Ime2. Genes Dev. 2003, 17, 1524–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasmyth, K. Control of the yeast cell cycle by the Cdc28 protein kinase. Curr. Opin. Cell Biol. 1993, 5, 166–179. [Google Scholar] [CrossRef]
- Stuart, D.; Wittenberg, C. CLB5 and CLB6 are required for premeiotic DNA replication and activation of the meiotic S/M checkpoint. Genes Dev. 1998, 12, 2698–2710. [Google Scholar] [CrossRef]
- Smith, K.N.; Penkner, A.; Ohta, K.; Klein, F.; Nicolas, A. B-type cyclins CLB5 and CLB6 control the initiation of recombination and synaptonemal complex formation in yeast meiosis. Curr. Biol. 2001, 11, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Sasanuma, H.; Hirota, K.; Fukuda, T.; Kakusho, N.; Kugou, K.; Kawasaki, Y.; Shibata, T.; Masai, H.; Ohta, K. Cdc7-dependent phosphorylation of Mer2 facilitates initiation of yeast meiotic recombination. Genes Dev. 2008, 22, 398–410. [Google Scholar] [CrossRef] [Green Version]
- Keeney, S. Mechanism and control of meiotic recombination initiation. Curr. Top. Dev. Biol. 2001, 52, 1–53. [Google Scholar] [CrossRef]
- Boudolf, V.; Barrôco, R.; de Almeida Engler, J.; Verkest, A.; Beeckman, T.; Naudts, M.; Inzé, D.; De Veylder, L. B1-Type Cyclin-Dependent Kinases Are Essential for the Formation of Stomatal Complexes in Arabidopsis thaliana. Plant Cell 2004, 16, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Boniotti, M.B.; Gutierrez, C. A cell-cycle-regulated kinase activity phosphorylates plant retinoblastoma protein and contains, in Arabidopsis, a CDKA/cyclin D complex. Plant J. 2001, 28, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Wijnker, E.; Harashima, H.; Müller, K.; Parra-Nuñez, P.; de Snoo, C.B.; van de Belt, J.; Dissmeyer, N.; Bayer, M.; Pradillo, M.; Schnittger, A. The Cdk1/Cdk2 homolog CDKA;1 controls the recombination landscape in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 12534–12539. [Google Scholar] [CrossRef] [PubMed]
- Corellou, F.; Camasses, A.; Ligat, L.; Peaucellier, G.; Bouget, F.Y. Atypical regulation of a green lineage-specific B-type cyclin-dependent kinase. Plant Physiol. 2005, 138, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Uchimiya, H.; Umeda, M. Expression of B2-type cyclin-dependent kinase is controlled by protein degradation in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 1683–1686. [Google Scholar] [CrossRef]
- Bulankova, P.; Akimcheva, S.; Fellner, N.; Riha, K. Identification of Arabidopsis Meiotic Cyclins Reveals Functional Diversification among Plant Cyclin Genes. PLoS Genet. 2013, 9, e1003508. [Google Scholar] [CrossRef] [PubMed]
- Potuschak, T.; Doerner, P. Cell cycle controls: Genome-wide analysis in Arabidopsis. Curr. Opin. Plant Biol. 2001, 4, 501–506. [Google Scholar] [CrossRef]
- Nowack, M.K.; Harashima, H.; Dissmeyer, N.; Zhao, X.A.; Bouyer, D.; Weimer, A.K.; De Winter, F.; Yang, F.; Schnittger, A. Genetic Framework of Cyclin-Dependent Kinase Function in Arabidopsis. Dev. Cell 2012, 22, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Richard, C.; Granier, C.; Inze, D.; De Veylder, L. Analysis of cell division parameters and cell cycle gene expression during the cultivation of Arabidopsis thaliana cell suspensions. J. Exp. Bot. 2001, 52, 1625–1633. [Google Scholar] [CrossRef]
- Wijnker, E.; Schnittger, A. Control of the meiotic cell division program in plants. Plant Reprod. 2013, 26, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Nibau, C.; Phillips, D.W.; Jenkins, G.; Armstrong, S.J.; Doonan, J.H. CDKG1 protein kinase is essential for synapsis and male meiosis at high ambient temperature in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2014, 111, 2182–2187. [Google Scholar] [CrossRef] [PubMed]
- Azumi, Y.; Liu, D.; Zhao, D.; Li, W.; Wang, G.; Hu, Y.; Ma, H. Homolog interaction during meiotic prophase I in Arabidopsis requires the SOLO DANCERS gene encoding a novel cyclin-like protein. EMBO J. 2002, 21, 3081–3095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Erfurth, I.; Cromer, L.; Jolivet, S.; Girard, C.; Horlow, C.; Sun, Y.; To, J.P.; Berchowitz, L.E.; Copenhaver, G.P.; Mercier, R. The cyclin-A CYCA1;2/TAM is required for the meiosis I to meiosis II transition and cooperates with OSD1 for the prophase to first meiotic division transition. PLoS Genet. 2010, 6, e1000989. [Google Scholar] [CrossRef] [PubMed]
- Bulankova, P.; Riehs-Kearnan, N.; Nowack, M.K.; Schnittger, A.; Riha, K. Meiotic progression in Arabidopsis is governed by complex regulatory interactions between SMG7, TDM1, and the meiosis I-specific cyclin TAM. Plant Cell 2010, 22, 3791–3803. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewicz, A.; Lange, J.; Keeney, S.; Jasin, M. Control of meiotic double-strand-break formation by ATM: Local and global views. Cell Cycle 2018, 17, 1155–1172. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.J.; Wardell, K.; Garcia, V.; Neale, M.J. Homeostatic regulation of meiotic DSB formation by ATM/ATR. Exp. Cell Res. 2014, 329, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Penedos, A.; Johnson, A.L.; Strong, E.; Goldman, A.S.; Carballo, J.A.; Cha, R.S. Essential and Checkpoint Functions of Budding Yeast ATM and ATR during Meiotic Prophase Are Facilitated by Differential Phosphorylation of a Meiotic Adaptor Protein, Hop1. PLoS ONE 2015, 10, e0134297. [Google Scholar] [CrossRef]
- Shiloh, Y. The ATM-mediated DNA-damage response: Taking shape. Trends Biochem. Sci. 2006, 31, 402–410. [Google Scholar] [CrossRef]
- Carballo, J.A.; Panizza, S.; Serrentino, M.E.; Johnson, A.L.; Geymonat, M.; Borde, V.; Klein, F.; Cha, R.S. Budding yeast ATM/ATR control meiotic double-strand break (DSB) levels by down-regulating Rec114, an essential component of the DSB-machinery. PLoS Genet. 2013, 9, e1003545. [Google Scholar] [CrossRef]
- Zhang, L.; Kim, K.P.; Kleckner, N.E.; Storlazzi, A. Meiotic double-strand breaks occur once per pair of (sister) chromatids and, via Mec1/ATR and Tel1/ATM, once per quartet of chromatids. Proc. Natl. Acad. Sci. USA 2011, 108, 20036–20041. [Google Scholar] [CrossRef] [Green Version]
- Lange, J.; Pan, J.; Cole, F.; Thelen, M.P.; Jasin, M.; Keeney, S. ATM controls meiotic double-strand-break formation. Nature 2011, 479, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Kleckner, N. Sequence non-specific double-strand breaks and interhomolog interactions prior to double-strand break formation at a meiotic recombination hot spot in yeast. EMBO J. 1995, 14, 5115–5128. [Google Scholar] [CrossRef] [PubMed]
- Blitzblau, H.G.; Hochwagen, A. ATR/Mec1 prevents lethal meiotic recombination initiation on partially replicated chromosomes in budding yeast. eLife 2013, 2, e00844. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ajimura, M.; Padmore, R.; Klein, C.; Kleckner, N. NDT80, a meiosis-specific gene required for exit from pachytene in Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 6572. [Google Scholar] [CrossRef] [PubMed]
- Winter, E. The Sum1/Ndt80 Transcriptional Switch and Commitment to Meiosis in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2012, 76, 1. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.; Allison, R.M.; Garcia, V.; Goldman, A.S.; Neale, M.J. Positive regulation of meiotic DNA double-strand break formation by activation of the DNA damage checkpoint kinase Mec1(ATR). Open Biol. 2013, 3, 130019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argunhan, B.; Farmer, S.; Leung, W.K.; Terentyev, Y.; Humphryes, N.; Tsubouchi, T.; Toyoizumi, H.; Tsubouchi, H. Direct and indirect control of the initiation of meiotic recombination by DNA damage checkpoint mechanisms in budding yeast. PLoS ONE 2013, 8, e65875. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.J.; Garcia, V.; Neale, M.J. Meiotic DSB patterning: A multifaceted process. Cell Cycle 2016, 15, 13–21. [Google Scholar] [CrossRef]
- Kauppi, L.; Barchi, M.; Lange, J.; Baudat, F.; Jasin, M.; Keeney, S. Numerical constraints and feedback control of double-strand breaks in mouse meiosis. Genes Dev. 2013, 27, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Ryu, T.H.; Lee, S.S.; Lee, S.; Chung, B.Y. Ionizing radiation manifesting DNA damage response in plants: An overview of DNA damage signaling and repair mechanisms in plants. Plant Sci. 2019, 278, 44–53. [Google Scholar] [CrossRef]
- Culligan, K.M.; Britt, A.B. Both ATM and ATR promote the efficient and accurate processing of programmed meiotic double-strand breaks. Plant J. 2008, 55, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.; Bruchet, H.; Camescasse, D.; Granier, F.; Bouchez, D.; Tissier, A. AtATM is essential for meiosis and the somatic response to DNA damage in plants. Plant Cell 2003, 15, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. ATM to the rescue: Repairing DNA damage. Plant Cell 2003, 15, 1–3. [Google Scholar] [CrossRef]
- Roitinger, E.; Hofer, M.; Kocher, T.; Pichler, P.; Novatchkova, M.; Yang, J.; Schlogelhofer, P.; Mechtler, K. Quantitative phosphoproteomics of the ataxia telangiectasia-mutated (ATM) and ataxia telangiectasia-mutated and rad3-related (ATR) dependent DNA damage response in Arabidopsis thaliana. Mol. Cell. Proteom. MCP 2015, 14, 556–571. [Google Scholar] [CrossRef]
- Prugar, E.; Burnett, C.; Chen, X.; Hollingsworth, N.M. Coordination of Double Strand Break Repair and Meiotic Progression in Yeast by a Mek1-Ndt80 Negative Feedback Loop. Genetics 2017, 206, 497–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blat, Y.; Protacio, R.U.; Hunter, N.; Kleckner, N. Physical and Functional Interactions among Basic Chromosome Organizational Features Govern Early Steps of Meiotic Chiasma Formation. Cell 2002, 111, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Borde, V.; de Massy, B. Programmed induction of DNA double strand breaks during meiosis: Setting up communication between DNA and the chromosome structure. Curr. Opin. Genet. Dev. 2013, 23, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Keeney, S. Spo11 and the Formation of DNA Double-Strand Breaks in Meiosis. Genome Dyn. Stab. 2008, 2, 81–123. [Google Scholar] [CrossRef] [PubMed]
- Kleckner, N. Chiasma formation: Chromatin/axis interplay and the role(s) of the synaptonemal complex. Chromosoma 2006, 115, 175. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, L.; Szekvolgyi, L.; Dichtl, B.; Dichtl, B.S.; de La Roche Saint Andre, C.; Nicolas, A.; Geli, V. The COMPASS subunit Spp1 links histone methylation to initiation of meiotic recombination. Science 2013, 339, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Sommermeyer, V.; Beneut, C.; Chaplais, E.; Serrentino, M.E.; Borde, V. Spp1, a member of the Set1 Complex, promotes meiotic DSB formation in promoters by tethering histone H3K4 methylation sites to chromosome axes. Mol. Cell 2013, 49, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Yoshida, K.; Matsui, Y. A histone H3 methyltransferase controls epigenetic events required for meiotic prophase. Nature 2005, 438, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Billings, T.; Petkov, P.M. CXXC1 is not essential for normal DNA double-strand break formation and meiotic recombination in mouse. PLoS Genet. 2018, 14, e1007657. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Baudat, F.; Taillepierre, M.; Stanzione, M.; Toth, A.; de Massy, B. The PRDM9 KRAB domain is required for meiosis and involved in protein interactions. Chromosoma 2017, 126, 681–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvanov, E.D.; Tian, H.; Billings, T.; Saxl, R.L.; Spruce, C.; Aithal, R.; Krejci, L.; Paigen, K.; Petkov, P.M. PRDM9 interactions with other proteins provide a link between recombination hotspots and the chromosomal axis in meiosis. Mol. Biol. Cell 2017, 28, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Ronceret, A.; Golubovskaya, I.; Ku, J.-C.; Lee, D.H.; Timofejeva, L.; Angoa, A.K.G.; Kao, Y.-H.; Kremling, K.; Williams-Carrier, R.; Meeley, R.; et al. The dynamic association of SPO11-1 with conformational changes of meiotic axial elements in maize. bioRxiv 2018. [Google Scholar] [CrossRef]
- Ferdous, M.; Higgins, J.D.; Osman, K.; Lambing, C.; Roitinger, E.; Mechtler, K.; Armstrong, S.J.; Perry, R.; Pradillo, M.; Cunado, N.; et al. Inter-homolog crossing-over and synapsis in Arabidopsis meiosis are dependent on the chromosome axis protein AtASY3. PLoS Genet. 2012, 8, e1002507. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kao, Y.H.; Ku, J.C.; Lin, C.Y.; Meeley, R.; Jan, Y.S.; Wang, C.J. The Axial Element Protein DESYNAPTIC2 Mediates Meiotic Double-Strand Break Formation and Synaptonemal Complex Assembly in Maize. Plant Cell 2015, 27, 2516–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, G.; Phillips, D.; Mikhailova, E.I.; Timofejeva, L.; Jones, R.N. Meiotic genes and proteins in cereals. Cytogenet. Genome Res. 2008, 120, 291–301. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Arabidopsis Thaliana | Oryza Sativa | Saccharomyces Cerevisiae | Schizosaccharomyces Pombe | Mus Musculus |
|---|---|---|---|---|
| DFO | ||||
| PRD1 | MEI1 | |||
| PRD2 | Mei4 | Rec24 | MEI4 | |
| PRD3 | PAIR1 | Mer2/Rec107 | Rec15 | IHO1 |
| Spo11-1,2 | Spo11-1,4 | Spo11 | Rec12 | SPO11 |
| TopoVI B | TopoVI B | TopoVI B | ||
| P31comet/BVF1 | P31comet | |||
| SDS | SDS | |||
| - | CRC1 | |||
| Rec102 | ||||
| Rec104 | ||||
| Rec6 | ||||
| Rec114 | Rec7 | REC114 | ||
| Rec10 | ||||
| Ski8/Rec103 | Rec14 | WDR61 | ||
| Mde2 | ||||
| MRE11 | Mre11 | Rad32 | ||
| RAD50 | Rad50 | Rad50 | RAD50 | |
| NBS1 | Xrs2 | Nbs1 | NBS1 |
| Species | Genome Size | Chromosome No. | Number of DSBs | DSB Hotspot No. | Most Common DSB Location | Average Width (kb) | Predominantly DSB Formation Among Transposon | Method | Hotspot Detection | References |
|---|---|---|---|---|---|---|---|---|---|---|
| S. cerevisiae (SK1) | 12.1 Mb | 16 | ~175 | 3604–4099 | Gene promoters | 0.248–0.264 | Ty retrotransposons | SPO11-oligos | Enrichment threshold | [92,101,110,111] |
| S. cerevisiae (YPS128) | 12.1 Mb | 16 | ~175 | 4177 | Gene promoters | 0.265 | n/a | SPO11-oligos | Enrichment threshold | [112] |
| S. cerevisiae (UWOPS03-461.4) | 12.1 Mb | 16 | ~175 | 3881 | Gene promoters | 0.256 | n/a | SPO11-oligos | Enrichment threshold | [112] |
| S. pombe | 13.8 Mb | 3 | ~60 | 603 | All chromosome regions | 1.4 | n/a | Rec12-oligos | Enrichment threshold | [113] |
| M. musculus (9R×13R) | 2.8 Gb | 20 | ~250 | 9874– 15,677 | Intergenic | ~2.000–3.400 | LTR retrotransposons SINE | SSDS | Peak calling | [103,104] |
| M. musculus (9R) | 2.8 Gb | 20 | ~250 | 14,869 | Intergenic | ~2.000 | n/a | SSDS | Peak calling | [104] |
| M. musculus (13R) | 2.8 Gb | 20 | ~250 | 15,481 | Intergenic | ~2.000 | n/a | SSDS | Peak calling | [104] |
| M. musculus (B6) | 2.8 Gb | 20 | ~250 | 18,313 | Intergenic | ~2.000 | n/a | SSDS | Peak calling | [104] |
| M. musculus (B6) | 2.8 Gb | 20 | ~250 | 13,960 | Intergenic | ~0.281 | n/a | SPO11-oligos | Enrichment threshold | [100] |
| Arabidopsis thaliana | 135 Mb | 5 | ~250–300 | 5914 | Gene promoters and terminators | 0.823 | Helitron /Pogo/Tc1/Mariner DNA transposons | SPO11-1-oligos | Peak calling | [99] |
| Zea mays | 2.4 Gb | 10 | ~500 | 3126 | All chromosome regions | 1.2 | Gypsy retrotransposons | SSDS | Peak calling | [105] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, J.-L.; Zhang, T.; Wang, Y.-Z.; He, Y. Advances Towards How Meiotic Recombination Is Initiated: A Comparative View and Perspectives for Plant Meiosis Research. Int. J. Mol. Sci. 2019, 20, 4718. https://doi.org/10.3390/ijms20194718
Jing J-L, Zhang T, Wang Y-Z, He Y. Advances Towards How Meiotic Recombination Is Initiated: A Comparative View and Perspectives for Plant Meiosis Research. International Journal of Molecular Sciences. 2019; 20(19):4718. https://doi.org/10.3390/ijms20194718
Chicago/Turabian StyleJing, Ju-Li, Ting Zhang, Ya-Zhong Wang, and Yan He. 2019. "Advances Towards How Meiotic Recombination Is Initiated: A Comparative View and Perspectives for Plant Meiosis Research" International Journal of Molecular Sciences 20, no. 19: 4718. https://doi.org/10.3390/ijms20194718
APA StyleJing, J. -L., Zhang, T., Wang, Y. -Z., & He, Y. (2019). Advances Towards How Meiotic Recombination Is Initiated: A Comparative View and Perspectives for Plant Meiosis Research. International Journal of Molecular Sciences, 20(19), 4718. https://doi.org/10.3390/ijms20194718