Multilevel Regulation of Peroxisomal Proteome by Post-Translational Modifications




Abstract
:1. Introduction to Peroxisomal Metabolism, Function, and Dynamics
2. Post-Translational Modification of Peroxisomal Proteins
2.1. H2O2-Dependent Post-Translational Modifications
2.1.1. Protein Sulfenylation
Sulfenylated Peroxisomal Proteins
2.1.2. Protein Carbonylation
Carbonylated Peroxisomal Proteins
2.2. NO-Dependent PTMs
Peroxisomal Targets of NO-Dependent PTMs
2.3. H2S-Dependent Post-Translational Modifications: Protein Persulfidation
Persulfidated Peroxisomal Proteins
2.4. Phosphorylation
Phosporylated Peroxisomal Proteins
2.5. Other post-Translational Protein Modifications
3. Crosstalk between PTMs in the Regulation of Peroxisomal Metabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sandalio, L.M.; Romero-Puertas, M.C. Peroxisomes sense and respond to environmental cues by regulating ROS and RNS signalling networks. Annals Bot. 2015, 116, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, Y.T.; Gonzalez, K.L.; Bartel, B. Peroxisome function, biogenesis, and dynamics in plants. Plant Physiol. 2018, 176, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Reumann, S.; Bartel, B. Plant peroxisomes: Recent discoveries in functional complexity, organelle homeostasis, and morphological dynamics. Curr. Opin. Plant Biol. 2016, 34, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Galisteo, A.P.; Rodríguez-Serrano, M.; Pazmiño, D.M.; Gupta, D.K.; Sandalio, L.M.; and Romero-Puertas, M.C. S-Nitrosylated proteins in pea (Pisum sativum L.) leaf peroxisomes: Changes under abiotic stress. J. Exp. Bot. 2012, 63, 2089–2103. [Google Scholar] [CrossRef] [PubMed]
- Kataya, A.R.; Heidari, B.; Hagen, L.; Kommedal, R.; Slupphaug, G.; Lillo, C. Protein phosphatase 2A holoenzyme is targeted to peroxisomes by piggybacking and positively affects peroxisomal β-oxidation. Plant Physiol. 2015, 167, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Hu, J. Molecular basis of peroxisome division and proliferation in plants. Int. Rev. Cell Mol. Biol. 2010, 279, 79–99. [Google Scholar] [PubMed]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Sanz-Fernández, M.; Hu, J.; Sandalio, L.M. Peroxisomes extend peroxules in a fast response to stress via a reactive oxygen species-mediated induction of the peroxin PEX11a. Plant Physiol. 2016, 171, 1665–1674. [Google Scholar] [CrossRef] [PubMed]
- Aung, K.; Hu, J. Differential roles of Arabidopsis dynamin-related proteins DRP3A, DRP3B, and DRP5B in organelle division. J. Integr. Plant Biol. 2012, 54, 921–931. [Google Scholar]
- Goepfert, S.; Poirier, Y. β-Oxidation in fatty acid degradation and beyond. Cur. Opin. Plant Biol. 2007, 10, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Liu, J.; Hu, J. Peroxisomes in plant reproduction and seed-related development. J. Int. Plant Biol 2019, 61, 784–802. [Google Scholar] [CrossRef]
- Cornah, J.E.; Smith, S.M. Synthesis and function of glyoxylate cycle enzymes. In Plant Peroxisomes; Baker, A., Graham, L.A., Eds.; Elsevier Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 57–101. [Google Scholar]
- Widhalm, J.R.; Dudareva, N. A familiar ring to it: Biosynthesis of plant benzoic acids. Mol. Plant 2015, 8, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Adebesin, F.; Widhalm, J.R.; Lynch, J.H.; McCoy, R.M.; Dudareva, N. A peroxisomal thioesterase plays auxiliary roles in plant β-oxidative benzoic acid metabolism. Plant J. 2018, 93, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Dellero, Y.; Jossier, M.; Schmitz, J.; Maurino, V.G.; Hodges, M. Photorespiratory glycolate–glyoxylate metabolism. J. Exp. Bot. 2016, 67, 3041–3052. [Google Scholar] [CrossRef] [PubMed]
- Farmer, L.M.; Rinaldi, M.A.; Young, P.G.; Danan, C.H.; Burkhart, S.E.; Bartel, B. Disrupting autophagy restores peroxisome function to an Arabidopsis lon2 mutant and reveals a role for the LON2 protease in peroxisomal matrix protein degradation. Plant Cell 2013, 25, 4085–4100. [Google Scholar] [CrossRef] [PubMed]
- Goto-Yamada, S.; Mano, S.; Nakamori, C.; Kondo, M.; Yamawaki, R.; Kato, A.; Nishimura, M. Chaperone and protease functions of LON protease 2 modulate the peroxisomal transition and degradation with autophagy. Plant Cell Physiol. 2014, 55, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, L.A.; Pastori, G.M.; Palma, J.M.; Sandalio, L.M.; Sevilla, F.; Corpas, F.J.; Jimenez, A.; Lopez-Huertas, E.; Hernandez, J.A. The activated oxygen role ofperoxisomes in senescence. Plant Physiol. 1998, 116, 1195–1200. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Sparkes, I.; Hawes, C.; del Río, L.A.; Sandalio, L.M. Peroxisome dynamics in Arabidopsis plants under oxidative stress induced by cadmium Free Radic. Biol. Med. 2009, 47, 1632–1639. [Google Scholar]
- Sinclair, A.M.; Trobacher, C.P.; Mathur, N.; Greenwood, J.S.; Mathur, J. Peroxule extension over ER-defined paths constitutes a rapid subcellular response to hydroxyl stress. Plant J. 2009, 59, 231–242. [Google Scholar] [CrossRef]
- Ma, C.; Hagstrom, D.; Polley, S.G.; Subramani, S. Redox-regulated cargo binding and release by the peroxiosomal targeting signal receptor, Pex5. J. Biol. Chem. 2013, 288, 27220–27231. [Google Scholar] [CrossRef]
- Apanasets, O.; Grou, C.P.; Van Veldhoven, P.P.; Brees, C.; Wang, B.; Nordgren, M.; Dodt, G.; Azevedo, J.E.; Fransen, M. PEX5, the shuttling import receptor for peroxisomal matrix proteins, is a redox-sensitive protein. Traffic 2014, 15, 94–103. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Shibata, M.; Kondo, M.; Oikawa, K.; Sato, M.; Toyooka, K.; Shirasu, K.; Nishimura, M.; Ohsumi, Y. Organ-specific quality control of plant peroxisomes is mediated by autophagy. J. Cell Sci. 2014, 127, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Calero-Muñoz, N.; Exposito-Rodriguez, M.; Collado-Arenal, A.M.; Rodríguez-Serrano, M.; Laureano-Marín, A.M.; Santamaría, M.E.; Gotor, C.; Díaz, I.; Mullineaux, P.M.; Romero-Puertas, M.C.; et al. Cadmium induces reactive oxygen species dependent pexophagy in Arabidopsis leaves. Plant Cell Environ. 2019. [Google Scholar] [CrossRef] [PubMed]
- Voitsekhovskaja, O.V.; Schiermeyer, A.; Reumann, S. Plant peroxisomes are degraded by starvation-induced and constitutive autophagy in tobacco BY-2 suspension-cultured cells. Front. Plant Sci. 2014, 5, 629. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, A.; Komatsu, S. Impact of post-translational modifications of crop proteins under abiotic stress. Proteomes 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Walther, D. The Roles of Post-translational Modifications in the Context of Protein Interaction Networks. PLoS Comput. Biol. 2015, 11, e1004049. [Google Scholar] [CrossRef] [PubMed]
- Plant PTM Viewer. Available online: http://www.psb.ugent.be/webtools/ptm-viewer/index.php (accessed on 11 June 2019).
- The Arabidopsis Information Resource. Available online: https://www.arabidopsis.org (accessed on 11 June 2019).
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/protein (accessed on 11 June 2019).
- Pan, R.; Hu, J. Proteome of plant peroxisome. In Proteomics of peroxisomes: Identifying Novel Functions and Regulatory Networks; Del Rñío, L.A., Schrader, M., Eds.; Springer: Singapore, 2018; pp. 3–46. [Google Scholar]
- Venny Tool. Available online: http://bioinfogp.cnb.csic.es/tools/venny/ (accessed on 25 August 2019).
- Young, D.; Pedre, B.; Ezerinxa, D.; De Smet, B.; Lewandowska, A.; Tossounian, M.A.; Bodra, N.; Huang, J.; Astolfi-Rosado, L.; Van Breusegem, F.; et al. Protein Promiscuity in H2O2. Antioxid. Redox Sign. 2019, 30, 1285–1324. [Google Scholar] [CrossRef]
- Huang, J.; Willemsd, P.; Van Breusegemd, F.; Messens, J. Pathways crossing mammalian and plant sulfenomic landscapes. Free Radic. Biol. Med. 2018, 122, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Pye, V.E.; Christensen, C.E.; Dyer, J.H.; Arent, S.; Henriksen, A. Peroxisomal plant 3-ketoacyl-coa thiolase structure and activity are regulated by a sensitive redox switch. J. Biol. Chem. 2010, 285, 24078–24088. [Google Scholar] [CrossRef]
- Waszczak, C.; Akter, S.; Eeckhout, D.; Persiau, G.; Wahni, K.; Bodra, N.; Van Molle, I.; De Smet, B.; Vertommen, D.; Gevaert, K.; et al. Sulfenome mining in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2014, 111, 11545–11550. [Google Scholar] [CrossRef]
- Akter, S.; Carpentierg, S.; Van Breusegem, F.; Messens, J. Identification of dimedone-trapped sulfenylated proteins in plants under stress. Biochem. Biophys. Rep. 2017, 9, 106–113. [Google Scholar] [CrossRef]
- Poole, L.B.; Karplus, P.A.; Claiborne, A. Protein sulfenic acids in redox signaling. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 325–347. [Google Scholar] [CrossRef]
- Schmitz, J.; Rossoni, A.W.; Maurino, V.G. Dissecting the physiological function of plant glyoxalase I and glyoxalase I-like proteins. Front. Plant. Sci. 2018, 9, 1618. [Google Scholar] [CrossRef]
- Rao, R.S.; Møller, I.M. Pattern of occurrence and occupancy of carbonylation sites in proteins. Proteomics 2011, 11, 4166–4173. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Wong, C.M.; Marcocci, L.; Liu, L.; Suzuki, Y.J. Cell signaling by protein carbonylation and decarbonylation. Antioxid Redox Signal. 2010, 12, 393–404. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Palma, J.M.; Gómez, M.; Del Río, L.A.; Sandalio, L.M. Cadmium causes the oxidative modification of proteins in pea plants. Plant Cell Environ. 2002, 25, 677–686. [Google Scholar] [CrossRef]
- Nyström, T. Role of oxidative carbonylation in protein quality control and senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef]
- Lounifi, I.; Arc, E.; Molassiotis, A.; Job, D.; Rajjou, L.; Tanou, G. Interplay between protein carbonylation and nitrosylation in plants. Proteomics 2013, 13, 568–578. [Google Scholar] [CrossRef]
- Polge, C.; Jaquinod, M.; Holzer, F.; Bourguignon, J.; Walling, L.; Brouquisse, R. Evidence for the existence in Arabidopsis thaliana of the proteasome proteolytic pathway: Activation in response to cadmium. J. Biol Chem. 2009, 284, 35412–35424. [Google Scholar] [CrossRef]
- Xiong, Y.; Contento, A.L.; Nguyen, P.Q.; Bassham, D.C. Degradation of oxidized proteins by autophagy during oxidative stress in Arabidopsis. Plant Physiol. 2007, 143, 291–299. [Google Scholar] [CrossRef]
- Wehr, N.B.; Levine, R.L. Quantification of protein carbonylation. In Cell Senescence: Methods and Protocols, Methods in Molecular Biology; Galluzzi, L., Vitale, I., Kepp, O., Kroemer, G., Eds.; Humana: Louisville, KY, USA, 2013; pp. 265–281. [Google Scholar]
- Mano, J.; Nagata, M.; Okamura, S.; Shiraya, T.; Mitsui, T. Identification of oxidatively modified proteins in salt-stressed arabidopsis: A carbonyl-targeted proteomics approach. Plant Cell Physiol. 2014, 55, 1233–1244. [Google Scholar] [CrossRef]
- Weber, D.; Davies, M.J.; Grune, T. Determination of protein carbonyls in plasma, cell extracts, tissue homogenates, isolated proteins: Focus on sample preparation and derivatization conditions. Redox Biol. 2015, 5, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.; Donaldson, R.P. Metal-catalyzed oxidation induces carbonylation of peroxisomal proteins and loss of enzymatic activities. Arch. Biochem. Biophys. 2005, 439, 25–31. [Google Scholar] [CrossRef]
- Shibata, M.; Oikawa, K.; Yoshimoto, K.; Kondo, M.; Mano, S.; Yamada, K.; Hayashi, M.; Sakamoto, W.; Ohsumi, Y.; Nishimura, M. Highly oxidized peroxisomes are selectively degraded via autophagy in Arabidopsis. Plant Cell 2013, 25, 4967–4983. [Google Scholar] [CrossRef]
- Sanz, L.; Albertos, P.; Mateos, I.; Sánchez-Vicente, I.; Lechón, T.; Fernández Marcos, M.; Lorenzo, O. Nitric oxide (NO) and phytohormones crosstalk during early plant development. J. Exp. Bot. 2015, 66, 2857–2868. [Google Scholar] [CrossRef]
- Castillo, M.C.; Coego, A.; Costa-Broseta, Á.; León, J. Nitric oxide responses in Arabidopsis hypocotyls are mediated by diverse phytohormone pathways. J. Exp. Bot. 2018, 69, 5265–5278. [Google Scholar] [CrossRef] [Green Version]
- Trapet, P.; Kulik, A.; Lamotte, O.; Jeandroz, S.; Bourque, S.; Nicolas-Francès, V.; Rosnoblet, C.; Besson-Bard, A.; Wendehenne, D. NO signaling in plant immunity: A tale of messengers. Phytochemistry 2015, 112, 72–79. [Google Scholar] [CrossRef]
- Gupta, K.J.; Igamberdiev, A.U. Reactive nitrogen species in mitochondria and their implications in plant energy status and hypoxic stress tolerance. Front. Plant Sci. 2016, 7, 1–6. [Google Scholar] [CrossRef]
- Terrón-Camero, L.C.; Peláez-Vico, M.A.; Del Val, C.; Sandalio, L.M.; Romero-Puertas, M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versus endogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef]
- Martínez-Ruiz, A.; Lamas, S. S-Nitrosylation: A potential new paradigm in signal transduction. Cardiovasc. Res. 2004, 62, 43–52. [Google Scholar] [CrossRef]
- Janssen-Heininger, Y.M.; Mossman, B.T.; Heintz, N.H.; Forman, H.J.; Kalyanaraman, B.; Finkel, T.; Stamler, J.S.; Rhee, S.G.; van der Vliet, A. Redox-based regulation of signal transduction: Principles, pitfalls, and promises. Free Radic. Biol. Med. 2008, 45, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ruiz, A.; Araújo, I.M.; Izquierdo-Álvarez, A.; Hernansanz-Agustín, P.; Lamas, S.; Serrador, J.M. Specificity in S-Nitrosylation: A Short-Range Mechanism for NO Signaling? Antioxid. Redox Signal. 2013, 19, 1220–1235. [Google Scholar] [CrossRef] [Green Version]
- Romero-Puertas, M.C.; Rodríguez-Serrano, M.; Sandalio, L.M. Protein S-nitrosylation in plants under abiotic stress: An overview. Front. Plant Sci. 2013, 4, 373. [Google Scholar] [CrossRef]
- Spoel, S.H.; Tada, Y.; Loake, G.J. Post-translational protein modification as a tool for transcription reprogramming. New Phytol. 2010, 186, 333–339. [Google Scholar] [CrossRef]
- Cui, J.L.; Wang, Y.N.; Jiao, J.; Gong, Y.; Wang, J.H.; Wang, M.L. Fungal endophyte-induced salidroside and tyrosol biosynthesis combined with signal cross-talk and the mechanism of enzyme gene expression in Rhodiola crenulata. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Mengel, A.; Ageeva, A.; Georgii, E.; Bernhardt, J.; Wu, K.; Durner, J.; Lindermayr, C. Nitric oxide modulates histone acetylation at stress genes by inhibition of histone deacetylases. Plant Physiol. 2016, 173, 1434–1452. [Google Scholar] [CrossRef]
- Simontacchi, M.; García-Mata, C.; Bartoli, C.G.; Santa-María, G.E.; Lamattina, L. Nitric oxide as a key component in hormone-regulated processes. Plant Cell Rep. 2013, 32, 853–866. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Md Isa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marín-de la Rosa, N.; Conde, J.V.; Correia, C.S.; Pearce, S.P.; et al. Nitric oxide sensing in plants is mediated by proteolytic control of group vii erf transcription factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef]
- Albertos, P.; Romero-Puertas, M.C.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Nambara, E.; Lorenzo, O. S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth. Nat. Commun. 2015, 6, 8669. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Sandalio, L.M. Role of NO-dependent posttranslational modifications in switching metabolic pathways. Adv. Bot Res. 2016, 77, 123–144. [Google Scholar]
- Serrato, A.J.; Romero-Puertas, M.C.; Lázaro-Payo, A.; Sahrawy, M. Regulation by S-nitrosylation of the Calvin-Benson cycle fructose-1,6-bisphosphatase in Pisum sativum. Redox Biol. 2018, 14, 409–416. [Google Scholar] [CrossRef]
- Hu, J.; Huang, X.; Chen, L.; Sun, X.; Lu, C.; Zhang, L.; Wang, Y.; Zuo, J. Site-Specific nitrosoproteomic identification of endogenously s -nitrosylated proteins in Arabidopsis. Plant Physiol. 2015, 167, 1731–1746. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, I.; Lindermayr, C. Nitric oxide-based protein modification: Formation and site-specificity of protein S-nitrosylation. Front. Plant. Sci. 2013, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Kulik, A.; Koen, E.; Besson-Bard, A.; Bourque, S.; Jeandroz, S.; Lamotte, O.; Wendehenne, D. Protein S-nitrosylation: What’s going on in plants? Free Radic. Biol. Med. 2012, 53, 1101–1110. [Google Scholar] [CrossRef]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J. A physiological perspective on targets of nitration in NO-based signalling networks in plants. J. Exp. Bot. 2019, 70, 4379–4389. [Google Scholar] [CrossRef] [PubMed]
- Chaki, M.; Álvarez de Morales, P.; Ruiz, C.; Begara-Morales, J.C.; Barroso, J.B.; Corpas, F.J.; Palma, J.M. Ripening of pepper (Capsicum annuum) fruit is characterized by an enhancement of protein tyrosine nitration. Ann. Bot. 2015, 116, 637–647. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; Colom-Moreno, R.; León, J. In vivo protein tyrosine nitration in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 3501–3517. [Google Scholar] [CrossRef] [PubMed]
- Vandelle, E.; Delledonne, M. Peroxynitrite formation and function in plants. Plant Sci. 2011, 181, 534–539. [Google Scholar] [CrossRef]
- Martínez-Ruiz, A.; Lamas, S. Two decades of new concepts in nitric oxide signaling: From the discovery of a gas messenger to the mediation of nonenzymatic posttranslational modifications. IUBMB Life 2009, 61, 91–98. [Google Scholar] [CrossRef]
- Perazzolli, M.; Dominici, P.; Romero-Puertas, M.C.; Zago, E.; Zeier, J.; Sonoda, M.; Lamb, C.; Delledonne, M. Nonsymbiotic hemoglobin AHb1 modulates nitric oxide bioactivity in Arabidopsis thaliana. Plant Cell 2004, 16, 2785–2794. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Bykova, N.V.; Hill, R.D. Nitric oxide scavenging by barley hemoglobin is facilitated by a monodehydroascorbate reductase-mediated ascorbate reduction of methemoglobin. Planta 2006, 223, 1033–1040. [Google Scholar] [CrossRef]
- Smagghe, B.J.; Trent, I.I.I.J.T.; Hargrove, M.S. NO dioxygenase activity in hemoglobins is ubiquitous in vitro, but limited by reduction in vivo. PLoS ONE 2008, 3, e2039. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Begueria, L.; Cuypers, B.; Van Doorslaer, S.; Abbruzzetti, S.; Bruno, S.; Berghmans, H.; Dewilde, S.; Ramos, J.; Viappiani, C.; Becana, M. Characterization of the heme pocket structure and ligand binding kinetics of non-symbiotic hemoglobins from the model legume Lotus japonicus. Front. Plant. Sci. 2017, 8, 407. [Google Scholar] [CrossRef]
- Prado, A.M.; Porterfield, D.M.; Feijó, J.A. Nitric oxide is involved in pollen tube growth regulation and re-orientation of pollen tubes. Development 2004, 131, 2707–2714. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Barroso, J.B.; Carreras, A.; Quirós, M.; León, A.M.; Romero-Puertas, M.C.; Esteban, F.J.; Valderrama, R.; Palma, J.M.; Sandalio, L.M.; et al. Cellular and subcellular localization of endogenous nitric oxide in young and senescent pea plants. Plant Physiol. 2004, 136, 2722–2733. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A. Peroxisomes as a cellular source of reactive nitrogen species signal molecules. Arch. Biochem. Biophys. 2011, 506, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2018, 69, 3401–3411. [Google Scholar] [CrossRef]
- Godber, B.L.J.; Doel, J.J.; Durgan, J.; Eisenthal, R.; Harrison, R.A. A new route to peroxynitrite: A role for xanthine oxidoreductase A new route to peroxynitrite: A role for xanthine oxidoreductase. FEBS Lett. 2000, 475, 93–96. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Peroxynitrite (ONOO-) is endogenously produced in Arabidopsis peroxisomes and is overproduced under cadmium stress. Ann. Bot. 2014, 113, 87–96. [Google Scholar] [CrossRef]
- Fares, A.; Rossignol, M.; Peltier, J.-B. Proteomics investigation of endogenous S-nitrosylation inArabidopsis. Biochem. Biophys. Res. Comm. 2011, 416, 331–336. [Google Scholar] [CrossRef]
- Puyaubert, J.; Fares, A.; Rézé, N.; Peltier, J.-B.; Baudouin, E. Identification of endogenously S-nitrosylated proteins in Arabidopsis plantlets: Effect of cold stress on cysteine nitrosylation level. Plant Sci. 2014, 215, 150–156. [Google Scholar] [CrossRef]
- Morisse, S.; Zaffagnini, M.; Gao, X.-H.; Lemaire, S.D.; Marchand, C.H. Insight into protein S-nitrosylation in Chlamydomonas reinhardtii. Antioxid. Redox Signal. 2014, 21, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Holzmeister, C.; Gaupels, F.; Geerlof, A.; Sarioglu, H.; Sattler, M.; Durner, J.; Lindermayr, C. Differential inhibition of Arabidopsis superoxide dismutases by peroxynitrite-mediated tyrosine nitration. J. Exp. Bot. 2015, 66, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Begara-Morales, J.C.; Sánchez-Calvo, B.; Chaki, M.; Mata-Pérez, C.; Valderrama, R.; Padilla, M.N.; López-Jaramillo, J.; Luque, F.; Corpas, F.J.; Barroso, J.B. Differential molecular response of monodehydroascorbate reductase and glutathione reductase by nitration and S-nitrosylation. J. Exp. Bot. 2015, 66, 5983–5996. [Google Scholar] [CrossRef] [PubMed]
- Gotor, C.; Laureano-Marín, A.M.; Arenas-Alfonseca, L.; Moreno, I.; Aroca, A.; García, I.; Romero, L.C. Advances in Plant Sulfur Metabolism and Signaling. In Progress in Botany. Cánovas; Lüttge, U., Matyssek, R., Eds.; Springer: Cham, Switzerland, 2017; pp. 45–66. [Google Scholar]
- Hancock, J.T. Hydrogen sulfide and environmental stresses. Environ. Exp. Bot. 2019, 161, 50–56. [Google Scholar] [CrossRef]
- Li, Z.-G.; Min, X.; Zhou, Z.-H. Hydrogen Sulfide: A signal molecule in plant cross-adaptation. Front. Plant. Sci. 2016, 7, 1621. [Google Scholar] [CrossRef] [PubMed]
- Gotor, C.; Garcia, I.; Aroca, A.; Laureano-Marin, A.M.; Arenas-Alfonseca, L.; Jurado-Flores, A.; Moreno, I.; Romero, L.C. Signaling by hydrogen sulfide and cyanide through posttranslational modification. J. Exp. Bot. 2019, 70, 4251–4265. [Google Scholar] [CrossRef]
- Chen, J.; Wu, F.H.; Wang, W.H.; Zheng, C.J.; Lin, G.H.; Dong, X.J.; He, J.X.; Pei, Z.M.; Zheng, H.L. Hydrogen sulphide enhances photosynthesis through promoting chloroplast biogenesis, photosynthetic enzyme expression, and thiol redox modification in Spinacia oleracea seedlings. J. Exp. Bot. 2011, 62, 4481–4493. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, C.; Lai, D.; Sun, Y.; Samma, M.K.; Zhang, J.; Shen, W. Hydrogen sulfide delays GA-triggered programmed cell death in wheat aleurone layers by the modulation of glutathione homeostasis and heme oxygenase-1 expression. J. Plant. Physiol. 2014, 171, 53–62. [Google Scholar] [CrossRef]
- Gotor, C.; Garcia, I.; Crespo, J.L.; Romero, L.C. Sulfide as a signaling molecule in autophagy. Autophagy 2013, 9, 609–611. [Google Scholar] [CrossRef] [Green Version]
- Laureano-Marin, A.M.; Moreno, I.; Aroca, Á.; García, I.; Romero, L.C.; Gotor, C. Regulation of Autophagy by Hydrogen Sulfide. In Gasotransmitters in Plants: The Rise of a New Paradigm in Cell Signaling; Lamattina, L., García-Mata, C., Eds.; Springer: Cham, Switzerland, 2016; pp. 53–75. [Google Scholar]
- Jin, Z.; Xue, S.; Luo, Y.; Tian, B.; Fang, H.; Li, H.; Pei, Y. Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis. Plant Physiol. Biochem. 2013, 62, 41–46. [Google Scholar] [CrossRef]
- Scuffi, D.; Álvarez, C.; Laspina, N.; Gotor, C.; Lamattina, L.; García-Mata, C. Hydrogen sulfide generated by l-cysteine desulfhydrase acts upstream of nitric oxide to modulate abscisic acid-dependent stomatal closure. Plant Physiol. 2014, 166, 2065–2076. [Google Scholar] [CrossRef] [PubMed]
- Papanatsiou, M.; Scuffi, D.; Blatt, M.R.; García-Mata, C. Hydrogen sulfide regulates inward-rectifying k+ channels in conjunction with stomatal closure. Plant Physiol. 2015, 168, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wan, R.; Shi, Y.; Xue, S. Hydrogen sulfide activates s-type anion channel via ost1 and Ca(2+) modules. Mol. Plant 2016, 9, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Scuffi, D.; Nietzel, T.; Di Fino, L.M.; Meyer, A.J.; Lamattina, L.; Schwarzländer, M.; Laxalt, A.M.; García-Mata, C. Hydrogen sulfide increases production of NADPH oxidase-dependent hydrogen peroxide and phospholipase d-derived phosphatidic acid in guard cell signaling. Plant Physiol. 2018, 176, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical biology of H2S signaling through persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant. Sci. 2018, 9, 1369. [Google Scholar] [CrossRef] [Green Version]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [Green Version]
- Aroca, Á.; Serna, A.; Gotor, C.; Romero, L.C. S-Sulfhydration: A cysteine posttranslational modification in plant systems. Plant Physiol. 2015, 168, 334–342. [Google Scholar] [CrossRef]
- Aroca, A.; Schneider, M.; Scheibe, R.; Gotor, C.; Romero, L.C. Hydrogen sulfide regulates the cytosolic/nuclear partitioning of glyceraldehyde-3-phosphate dehydrogenase by enhancing its nuclear localization. Plant Cell Physiol. 2017, 58, 983–992. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Gonzalez-Gordo, S.; Munoz-Vargas, M.A.; Palma, J.M. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. J. Integr. Plant Biol. 2019, 61, 871–883. [Google Scholar] [CrossRef]
- Munoz-Vargas, M.A.; Gonzalez-Gordo, S.; Palma, J.M.; Corpas, F.J. Inhibition of NADP-malic enzyme activity by H2S and NO in sweet pepper (Capsicum annuum L.) fruits. Physiol. Plant 2019. [Google Scholar] [CrossRef]
- Munoz-Vargas, M.A.; Gonzalez-Gordo, S.; Canas, A.; Lopez-Jaramillo, J.; Palma, J.M.; Corpas, F.J. Endogenous hydrogen sulfide (H2S) is up-regulated during sweet pepper (Capsicum annuum L.) fruit ripening. In vitro analysis shows that NADP-dependent isocitrate dehydrogenase (ICDH) activity is inhibited by H2S and NO. Nitric. Oxide 2018, 81, 36–45. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente van Bentem, S.; Anrather, D.; Roitinger, E.; Djamei, A.; Hufnag, T.; Barta, A.; Csaszar, E.; Dohnal, I.; Lecourieux, D.; Hirt, H. Phosphoproteomics reveals extensive in vivo phosphorylation of Arabidopsis proteins involved in RNA metabolism. Nucleic. Acids Res. 2006, 34, 3267–3278. [Google Scholar] [CrossRef] [PubMed]
- Lillo, C.; Kataya, A.R.; Heidari, B.; Creighton, M.T.; Nemie-Feyissa, D.; Ginbot, Z.; Jonassen, E.M. Protein phosphatases PP2A, PP4 and PP6: Mediators and regulators in development and responses to environmental cues. Plant Cell Environ. 2014, 37, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.C.; Li, Z.; Graeber, T.G.; Graham, N.A.; Drake, J.M. Phosphopeptide enrichment coupled with label-free quantitative mass spectrometry to investigate the phosphoproteome in prostate cancer. J. Vis. Exp. 2018, 138, e57996. [Google Scholar] [CrossRef] [PubMed]
- Kataya, A.R.A.; Muenchen, D.G.; Moorhead, G.B. A framework to investigate peroxisomal protein phosphorylation in Arabidopsis. Trends Plant Sci. 2019, 24, 366–381. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Reumann, S. Improved prediction of peroxisomal PTS1 proteins from genome sequences based on experimental subcellular targeting analyses as exemplified for protein kinases from Arabidopsis. J. Exp. Bot. 2008, 59, 3767–3779. [Google Scholar] [CrossRef] [Green Version]
- Fukao, Y.; Hayashi, M.; Hara-Nishimura, I.; Nishimura, M. Novel glyoxysomal protein kinase, GPK1, identified by proteomic analysis of glyoxysomes in etiolated cotyledons of Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 1002–1012. [Google Scholar] [CrossRef]
- Dammann, C.; y Ichida, A.; Hong, B.; Romanowsky, S.M.; Hrabak, E.M.; Harmon, A.C.; Pickard, B.G.; Harper, J.H. Subcellular targeting of nine calcium dependent protein kinase isoforms from Arabidopsis. Plant Physiol. 2003, 132, 1840–1848. [Google Scholar] [CrossRef]
- Lee, H.Y.; Yoon, G.M. Kinase assay for CONSTITUTIVE TRIPLE RESPONSE 1 (CTR1) in Arabidopsis thaliana. Meth. Mol. Biol. 2017, 1573, 133–140. [Google Scholar]
- Hodges, M.; Jossier, M.; Boex-Fontvieille, E.; Tcherkez, G. Protein phosphorylation and photorespiration. Plant Biol. (Stuttg) 2013, 15, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Pazmiño, D.M. Contribución de las Especies de Oxígeno y Nitrógeno Reactivo, y de los Peroxisomas a la Toxicidad del 2,4-d en Plantas. Ph.D. Thesis, University of Granada, Granada, Spain, 2009. [Google Scholar]
- Zou, J.J.; Li, X.D.; Ratnasekera, D.; Wang, C.; Liu, W.Y.; Song, L.F.; Zhang, W.Z.; Wu, W.H. Arabidopsis CALCIUM-DEPENDENT PROTEIN KINASE8 and CATALASE3 Function in Abscisic Acid-Mediated Signaling and H2O2 Homeostasis in Stomatal Guard Cells under Drought Stress. Plant Cell 2015, 27, 1445–1460. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Batelli, G.; Grillo, S.; Agius, F.; Kim, Y.S.; Zhu, J.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.K. Interaction of SOS2 with Nucleoside Diphosphate Kinase 2 and Catalases Reveals a Point of Connection between Salt Stress and H2O2 Signaling in Arabidopsis thaliana. Mol. Cell Biol. 2007, 27, 7771–7780. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Yang, K.S.; Kang, S.W.; Woo, H.A.; Chan, T.S. Controlled elimination of intracellular H2O2: Regulation of peroxiredoxin, catalase, and glutathione peroxidase via post-translational modification. Antioxid. Redox Signal. 2013, 7, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Joshi, D.; Yadav, P.K.; Gupta, A.K.; Bhatt, T.K. Role of ubiquitin-mediated degradation system in plant biology. Front. Plant. Sci. 2016, 7, 806. [Google Scholar] [CrossRef]
- Platta, H.W.; El Magraoui, F.; Bäumer, B.E.; Schlee, D.; Girzalsky, W.; Erdmann, R. Pex2 and pex12 function as protein-ubiquitin ligases in peroxisomal protein import. Mol. Cell Biol. 2009, 29, 5505–5516. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, S.E.; Kao, Y.T.; Bartel, B. Peroxisomal ubiquitin-protein ligases peroxin2 and peroxin10 have distinct but synergistic roles in matrix protein import and peroxin5 retrotranslocation in Arabidopsis. Plant Physiol. 2014, 166, 1329–1344. [Google Scholar] [CrossRef]
- Kao, Y.T.; Fleming, W.A.; Ventura, M.J.; Bartel, B. Genetic interactions between peroxin12 and other peroxisome-associated ubiquitination components. Plant Physiol. 2016, 172, 1643–1656. [Google Scholar] [CrossRef]
- Song, G.; Walley, J.W. Dynamic Protein Acetylation in Plant–Pathogen Interactions. Front. Plant Sci. 2016, 7, 421. [Google Scholar] [CrossRef]
- Chen, H.; Li, S.; Li, L.; Wu, W.; Ke, X.; Zou, W.; Zhao, J. Na-Acetyltransferases 10 and 15 are required for the correct initiation of endosperm cellularization in Arabidopsis. Plant Cell Physiol. 2018, 59, 2113–2128. [Google Scholar] [CrossRef]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The world of protein acetylation. Biochim. Biophys. Acta 2016, 1864, 1372–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arent, S.; Rye, V.E.; Henriksen, A. Structure and function of plant acyl-CoA osidase. Plant Physiol. Biochem. 2008, 46, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.M.; Liu, W.C.; Lu, Y.T. CATALASE2 coordinates SA-mediated repression of both auxin accumulation and JA biosynthesis in plant defenses. Cell Host Microb. 2017, 21, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, Y.; Xie, Z.; Li, Y.; He, Z.H.; Peng, X.X. Association–dissociation of glycolate oxidase with catalase in rice: A potential switch to modulate intracellular H2O2 levels. Mol. Plant. 2016, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Hausladen, A.; Wang, Y.J.; Zhang, G.F.; Stomberski, C.; Brunengraber, H.; Hess, D.T.; Stamler, J.S. Identification of S-nitroso-CoA reductases that regulate protein S-nitrosylation. Proc. Natl. Acad. Sci. USA 2014, 111, 18572–18577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, D.P.; Skipsey, M.; Grundy, N.M.; Edwards, R. Stress-induced protein Sglutathionylation in Arabidopsis. Plant Physiol. 2005, 138, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Tanou, G.; Filippou, P.; Belghazi, M.; Job, D.; Diamantidis, G.; Fotopoulos, V.; Molassiotis, A. Oxidative and nitrosative-based signaling and associated post-translational modifications orchestrate the acclimation of citrus plants to salinity stress. Plant J. 2012, 72, 585–599. [Google Scholar] [CrossRef]
- Molassiotis, A.; Job, D.; Ziogas, V.; Tanou, G. Citrus plants: A model system for unlocking the secrets of no and ros-inspired priming against salinity and drought. Front. Plant Sci. 2016, 7, 229. [Google Scholar] [CrossRef] [PubMed]
- Baena, G.; Feria, A.B.; Echevarría, C.; Monreal, J.A.; García-Mauriño, S. Salinity promotes opposite patterns of carbonylation and nitrosylation of C4 phosphoenolpyruvate carboxylase in sorghum leaves. Planta 2017, 246, 1203–1214. [Google Scholar] [CrossRef]
- Gangwar, A.; Paul, S.; Ahmad, Y.; Bhargava, K. Competing trends of ROS and RNS-mediated protein modifications during hypoxia as an alternate mechanism of NO benefits. Biochimie 2018, 148, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.P. Signal transduction by protein tyrosine nitration: Competition or cooperation with tyrosine phosphorylation-dependent signaling events? Free Radic. Biol. Med. 2002, 33, 765–773. [Google Scholar] [CrossRef]
- Zhou, H.L.; Stomberski, C.T.; Stamler, J.S. Cross talk between S-nitrosylation and phosphorylation involving kinases and nitrosylases. Circ. Res. 2018, 122, 1485–1487. [Google Scholar] [CrossRef] [PubMed]
- Farré, J.C.; Mahalingam, S.S.; Proietto, M.; Subramani, S. Peroxisome biogenesis, membrane contact sites, and quality control. EMBO Rep. 2019, 20, e46864. [Google Scholar] [CrossRef] [PubMed]
- Bánréti, Á.; Sass, M.S.; Graba, Y. The emerging role of acetylation in the regulation of autophagy. Autophagy 2013, 9, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Oeljeklaus, S.; Schummer, A.; Mastalski, T.; Platta, H.W.; Warscheid, B. Regulation of peroxisome dynamics by phosphorylation. Biochim. Biophis. Acta 2016, 1863, 1227–1237. [Google Scholar]



{kind=link}
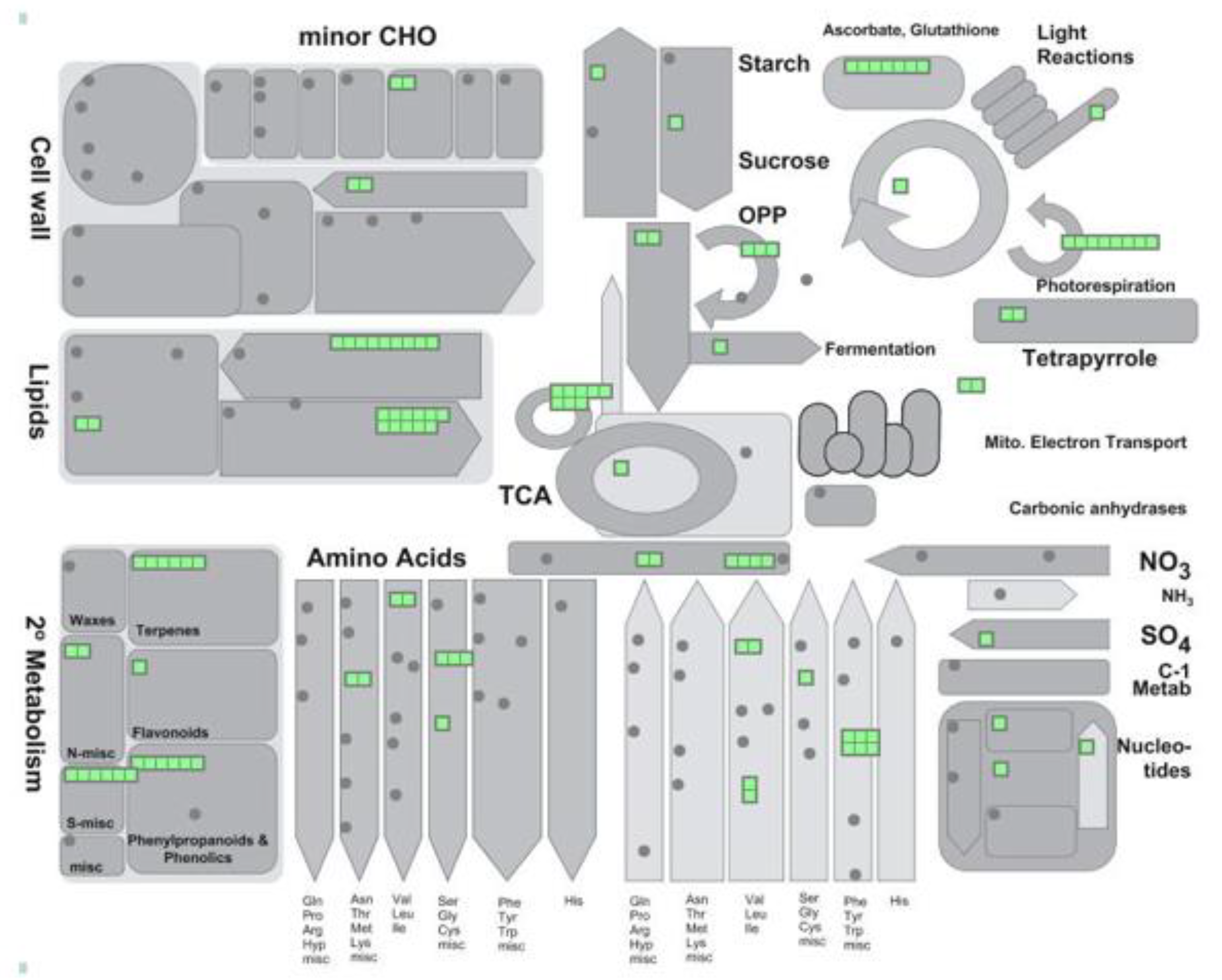{kind=link}
| Locus | Description | PTM_Types |
|---|---|---|
| REDOX/ASC-GLU CYCLE | ||
| AT1G19570 | dehydroascorbate reductase | ac; na; no; nt; ro |
| AT2G31570 | glutathione peroxidase 2 | ac; na; nt; ph |
| AT3G24170 | glutathione-disulfide reductase | ac; mo; na; no; nt; sf |
| AT3G27820 | monodehydroascorbate reductase 4 | ph; sf |
| AT3G52880 | monodehydroascorbate reductase 1 | ac; mo; ph; ps |
| AT4G35000 | ascorbate peroxidase 3 | ac; na; nt; ro; ub |
| AT4G35970 | ascorbate peroxidase 5 | na; nt; ro |
| AT1G20620 | catalase 3 | ac; na; no; nt; ph; ps; ntyr |
| AT1G20630 | catalase 1 | ac; no; nt; ph |
| AT4G35090 | catalase 2 | ac; na; no; nt; ph; ps; ca |
| AT5G18100 | copper/zinc superoxide dismutase 3 | ph |
| PHOTORESPIRATION | ||
| AT1G68010 | hydroxypyruvate reductase | ac; mo; na; no; nt; ph; ps |
| AT1G70580 | alanine-2-oxoglutarate aminotransferase 2 | ac; na; no; nt; ph; ro; ps |
| AT2G13360 | alanine:glyoxylate aminotransferase | ac; na; no; nt; ph; ps; ntyr |
| AT3G14130 | Aldolase-type TIM barrel family protein/ glycolate oxidase | ac; na |
| AT3G14150 | Aldolase-type TIM barrel family protein/ glycolate oxidase | na |
| AT3G14415 | Aldolase-type TIM barrel family protein/ glycolate oxidase | ac; na; ng; nt; ph; ntyr |
| AT3G14420 | Aldolase-type TIM barrel family protein/ glycolate oxidase | ac; na; ng; nt; ph; ntyr |
| AT4G18360 | Aldolase-type TIM barrel family protein/ glycolate oxidase 3 | ac; na; ph |
| AT4G39660 | alanine:glyoxylate aminotransferase 2 | ac |
| AT1G23310 | glutamate:glyoxylate aminotransferase | ac; na; no; nt; ph; ro; ub; ps |
| GLUCONEO/GLYOXYLATE CYCLE | ||
| AT1G54340 | isocitrate dehydrogenase | na; no; ro; ps |
| AT2G22780 | peroxisomal NAD-malate dehydrogenase 1 | no; nt; ps |
| AT2G42790 | citrate synthase 3 | ac; nt; ph; ub; ps |
| AT3G16910 | acyl-activating enzyme 7 | ac; nt; ps |
| AT3G21720 | isocitrate lyase | na; nt |
| AT3G58740 | citrate synthase 1 | ac; nt; ph |
| AT3G58750 | citrate synthase 2 | ac; no; nt; ph; ps |
| AT5G03860 | malate synthase | na; nt |
| AT5G09660 | peroxisomal NAD-malate dehydrogenase 2 | ac; mo; no; nt; ph; ro; ps; ntyr |
| LIPID METABOLISM-β OXIDATION | ||
| AT4G16760 | acyl-CoA oxidase 1 | ac; mo; na; nt; ps; sf |
| AT5G65110 | acyl-CoA oxidase 2 | no |
| AT1G06290 | acyl-CoA oxidase 3 | no; nt; ps |
| AT1G06310 | acyl-CoA oxidase 6 | no; ph; ps |
| AT1G20560 | acyl activating enzyme 1 | na; nt |
| AT1G66120 | AMP-dependent synthetase and ligase family protein | ub |
| AT1G77540 | Acyl-CoA N-acyltransferases (NAT) superfamily protein | na; nt; ph |
| AT1G76150 | enoyl-CoA hydratase 2 | na; nt; ro; ps |
| AT1G60550 | enoyl-CoA hydratase/isomerase D | ph; ps |
| AT2G30200 | EMBRYO DEFECTIVE 3147 | ac; mo; nt; ph |
| AT3G05970 | long-chain acyl-CoA synthetase 6 | nt; ps |
| AT3G06810 | acyl-CoA dehydrogenase-like protein | ac; na; ph; ps |
| AT3G06860 | multifunctional protein 2 | ac; nt; ph; ps |
| AT3G51840 | acyl-CoA oxidase 4 | na; nt; ph; ps |
| AT4G04320 | malonyl-CoA decarboxylase family protein | ph |
| AT4G16210 | enoyl-CoA hydratase/isomerase A | ac; mo; na; nt; ph; ps |
| AT4G27780 | acyl-CoA binding protein 2 | ph |
| AT4G29010 | Enoyl-CoA hydratase/isomerase family | ac; mo; na; nt; ps; ca |
| AT5G16370 | acyl activating enzyme 5 | na; nt |
| AT5G27600 | long-chain acyl-CoA synthetase 7 | nt |
| AT5G36880 | acetyl-CoA synthetase | ac; na; no; nt; ph |
| AT5G42890 | sterol carrier protein 2 | mo; na; nt |
| AT4G00520 | Acyl-CoA thioesterase family protein | ph |
| AT5G48880 | peroxisomal 3-keto-acyl-CoA thiolase 2 | ac; nt; ph; ps |
| AT2G33150 | peroxisomal 3-ketoacyl-CoA thiolase 3 | ac; mo; no; nt; ub; ps; sf |
| AT4G14440 | 3-hydroxyacyl-CoA dehydratase 1 | ph |
| AT3G26820 | Esterase/lipase/thioesterase family | ph |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandalio, L.M.; Gotor, C.; Romero, L.C.; Romero-Puertas, M.C. Multilevel Regulation of Peroxisomal Proteome by Post-Translational Modifications. Int. J. Mol. Sci. 2019, 20, 4881. https://doi.org/10.3390/ijms20194881
Sandalio LM, Gotor C, Romero LC, Romero-Puertas MC. Multilevel Regulation of Peroxisomal Proteome by Post-Translational Modifications. International Journal of Molecular Sciences. 2019; 20(19):4881. https://doi.org/10.3390/ijms20194881
Chicago/Turabian StyleSandalio, Luisa M., Cecilia Gotor, Luis C. Romero, and Maria C. Romero-Puertas. 2019. "Multilevel Regulation of Peroxisomal Proteome by Post-Translational Modifications" International Journal of Molecular Sciences 20, no. 19: 4881. https://doi.org/10.3390/ijms20194881
APA StyleSandalio, L. M., Gotor, C., Romero, L. C., & Romero-Puertas, M. C. (2019). Multilevel Regulation of Peroxisomal Proteome by Post-Translational Modifications. International Journal of Molecular Sciences, 20(19), 4881. https://doi.org/10.3390/ijms20194881