MicroRNA Assisted Gene Regulation in Colorectal Cancer




Abstract
:1. Introduction
2. Results
2.1. Identification of Candidate MicroRNA and Target Genes
2.2. MicroRNA Target Genes Associated with CRC and Their MFE (miRTarBase)
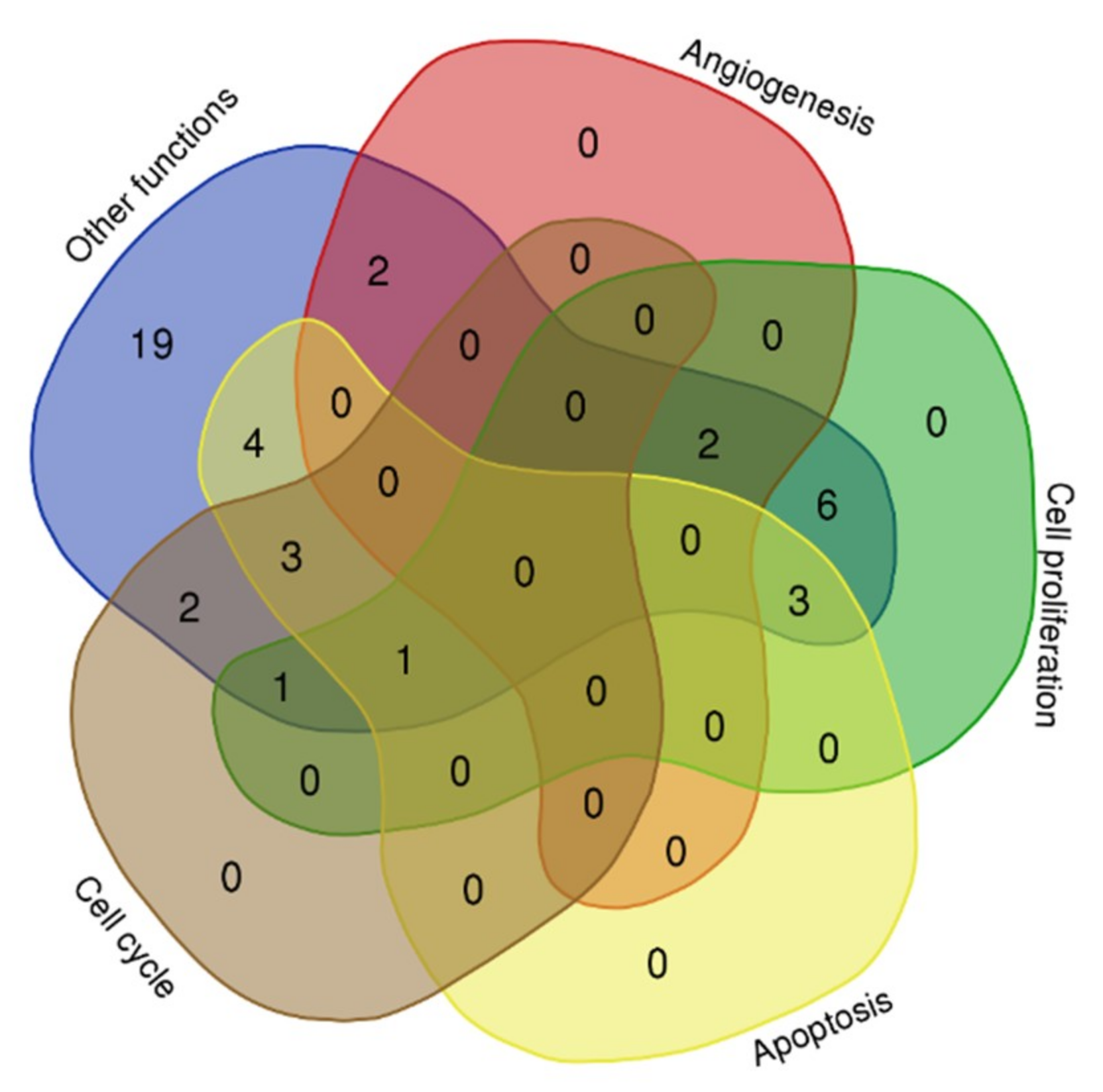
2.3. Biological Processes of the MicroRNA Target Genes
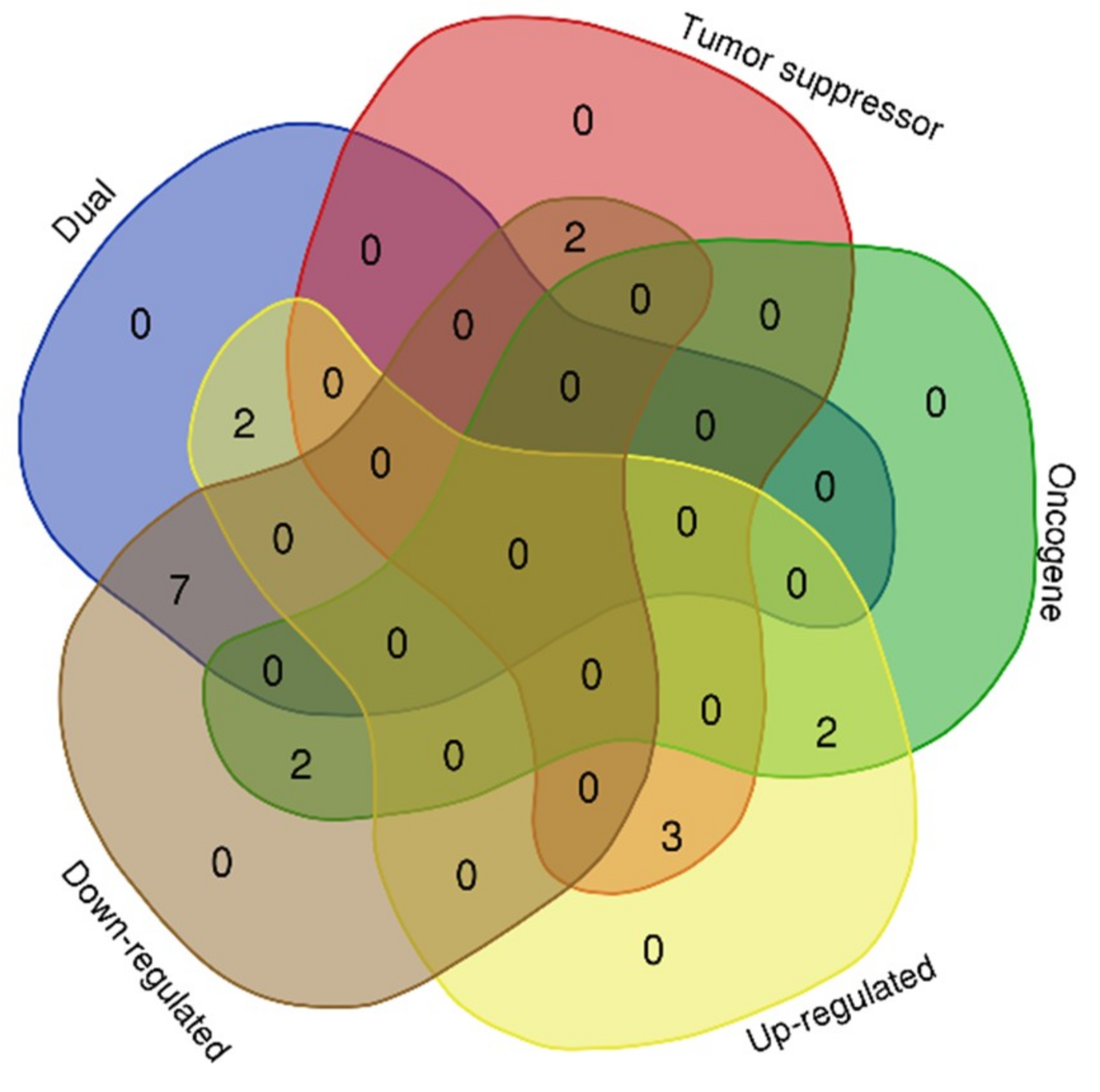
2.4. Gene Enrichment in Cancer and Their Biological Functions
2.5. Expressions MicroRNA Target Genes Involved in Several Cancer Subtypes
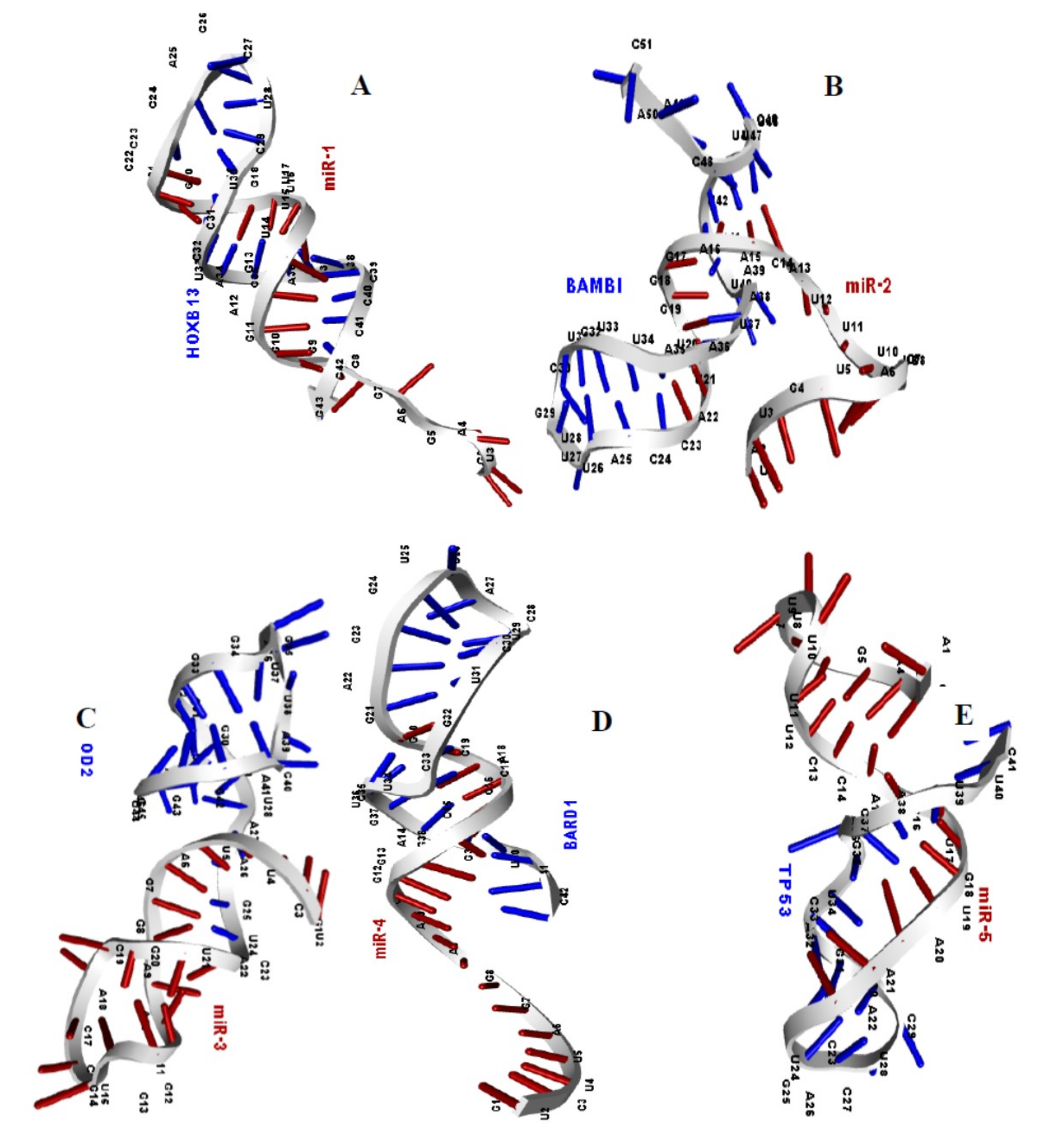
2.6. Binding Affinity and Structural Determination of MicroRNA and Duplex
2.7. Structural Model of MicroRNA-mRNA Duplexes
2.8. Extraction and Preparation of AGO Protein Structure
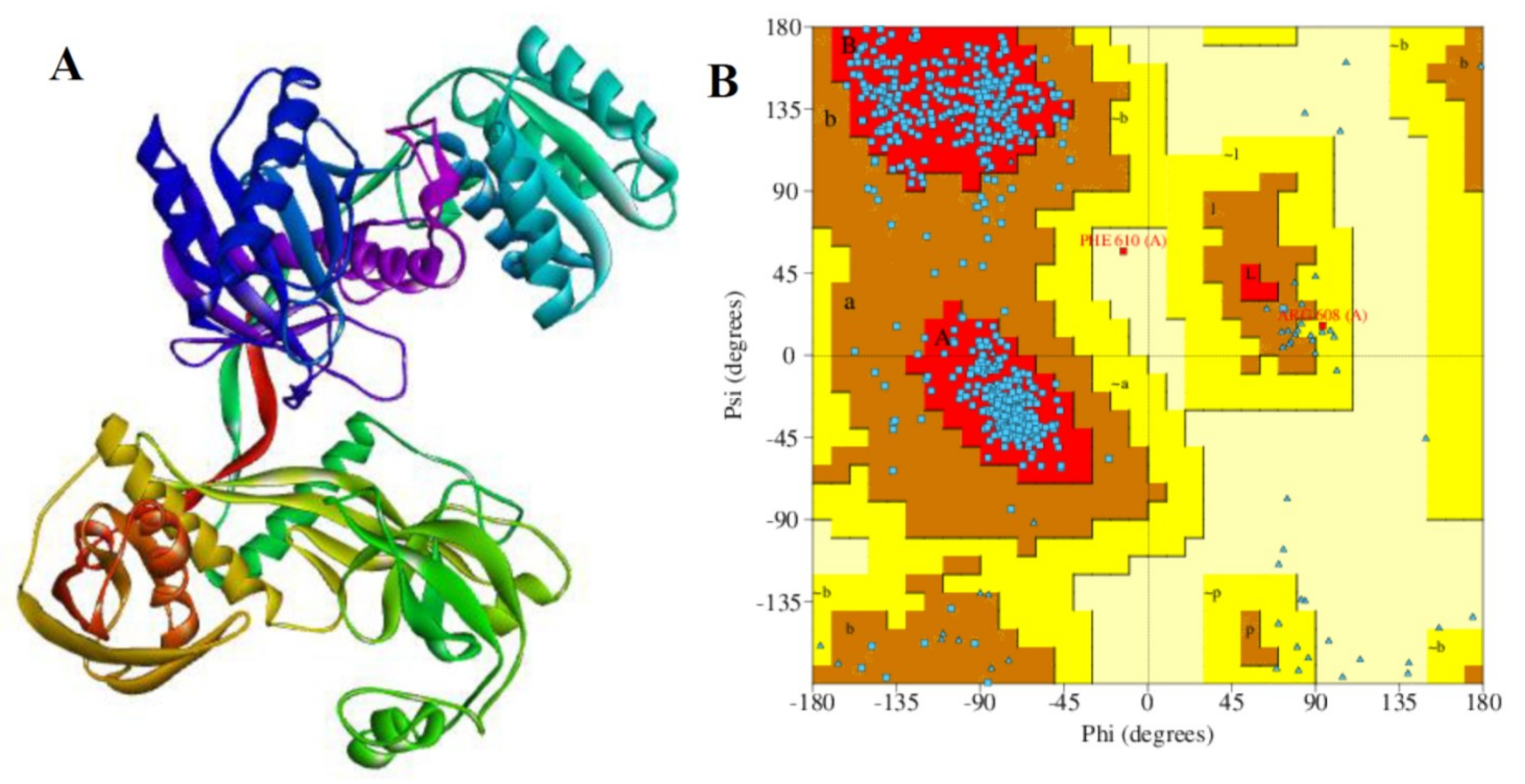
2.9. Validation of Chain A of Argonaute Protein
2.10. Docking Analysis Between Receptor Protein and MicroRNA
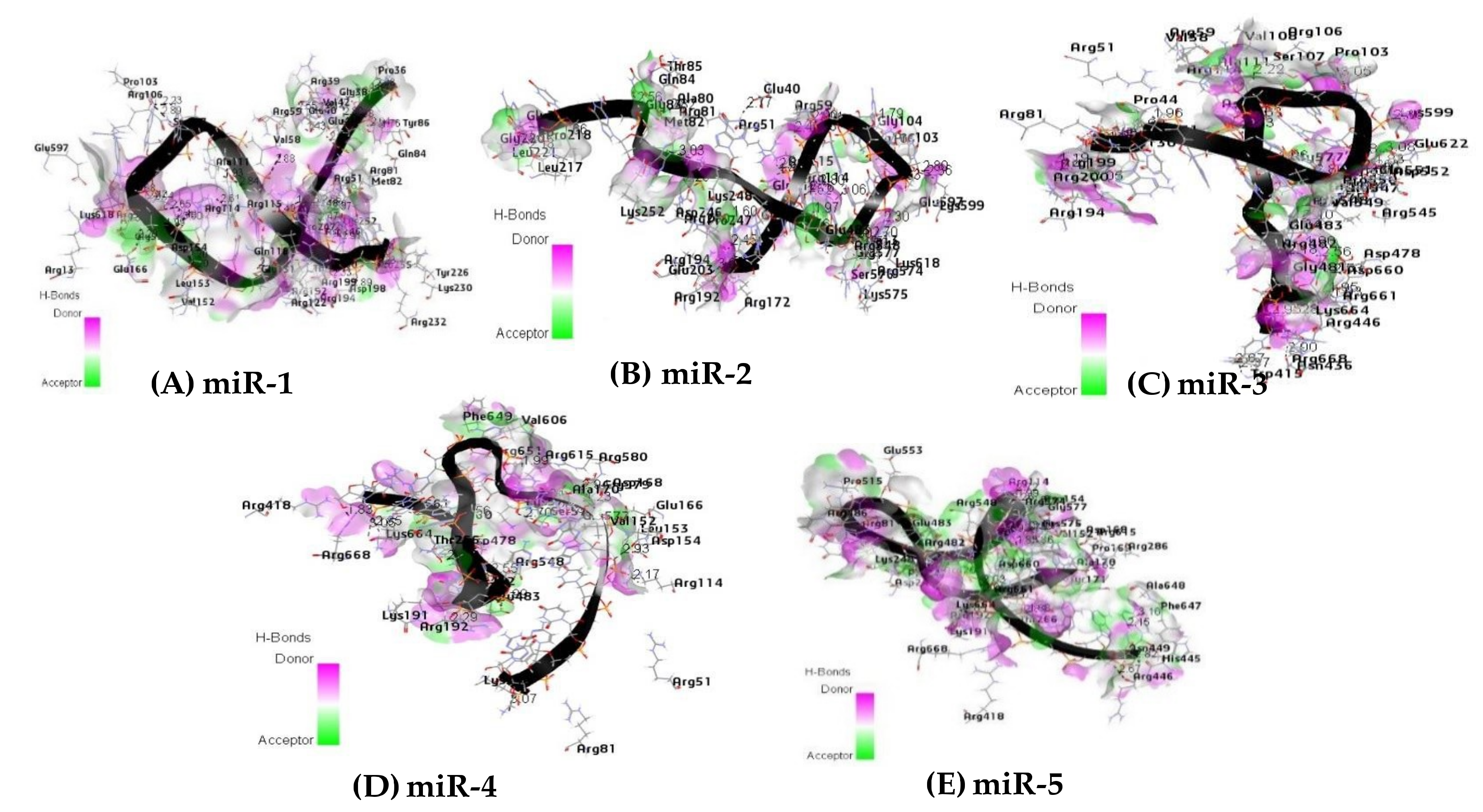
2.11. Hydrogen Bond Interaction
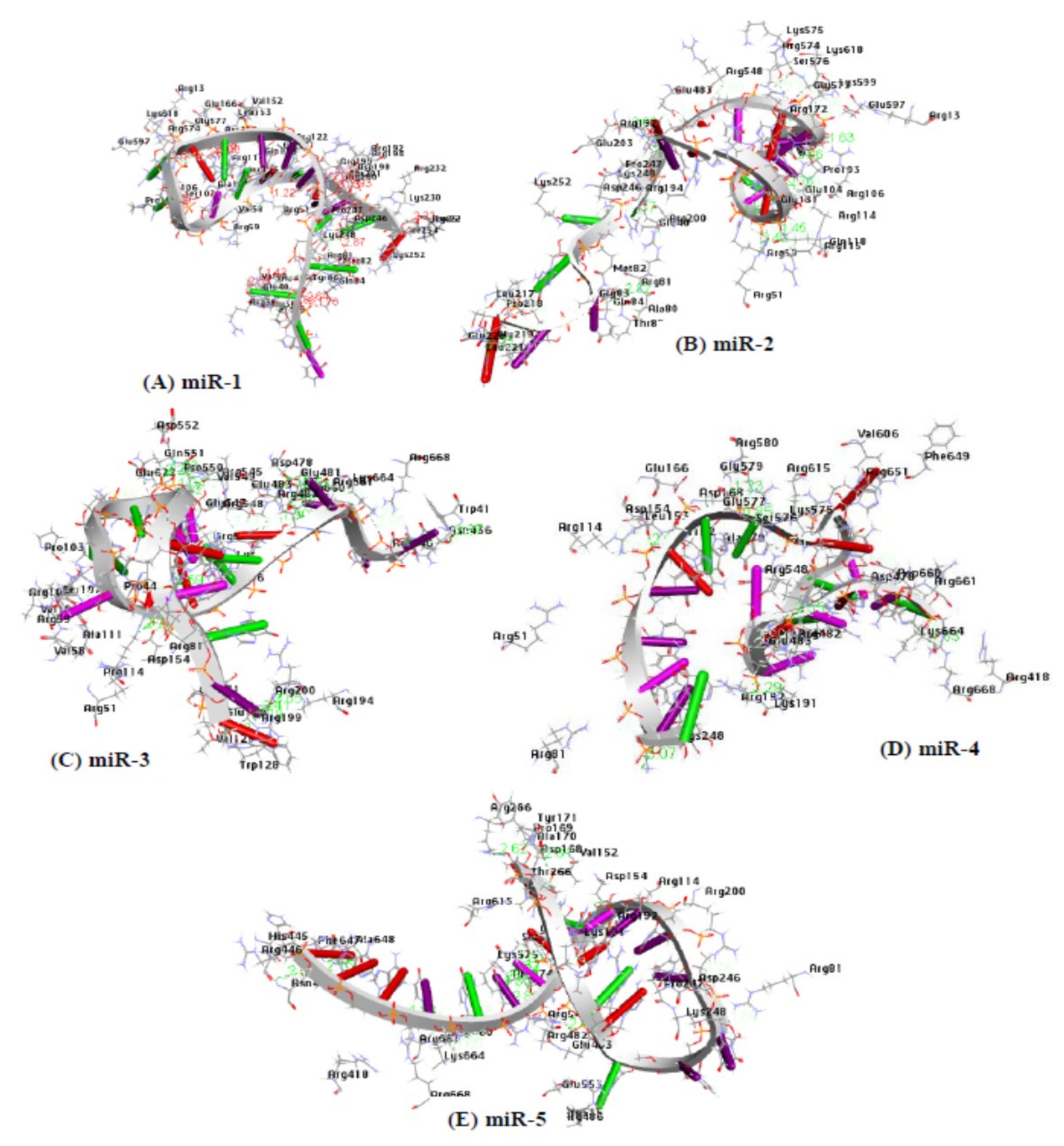
2.12. Docking Analysis between Argonaute Protein and MicroRNA-mRNA Complex
3. Discussion
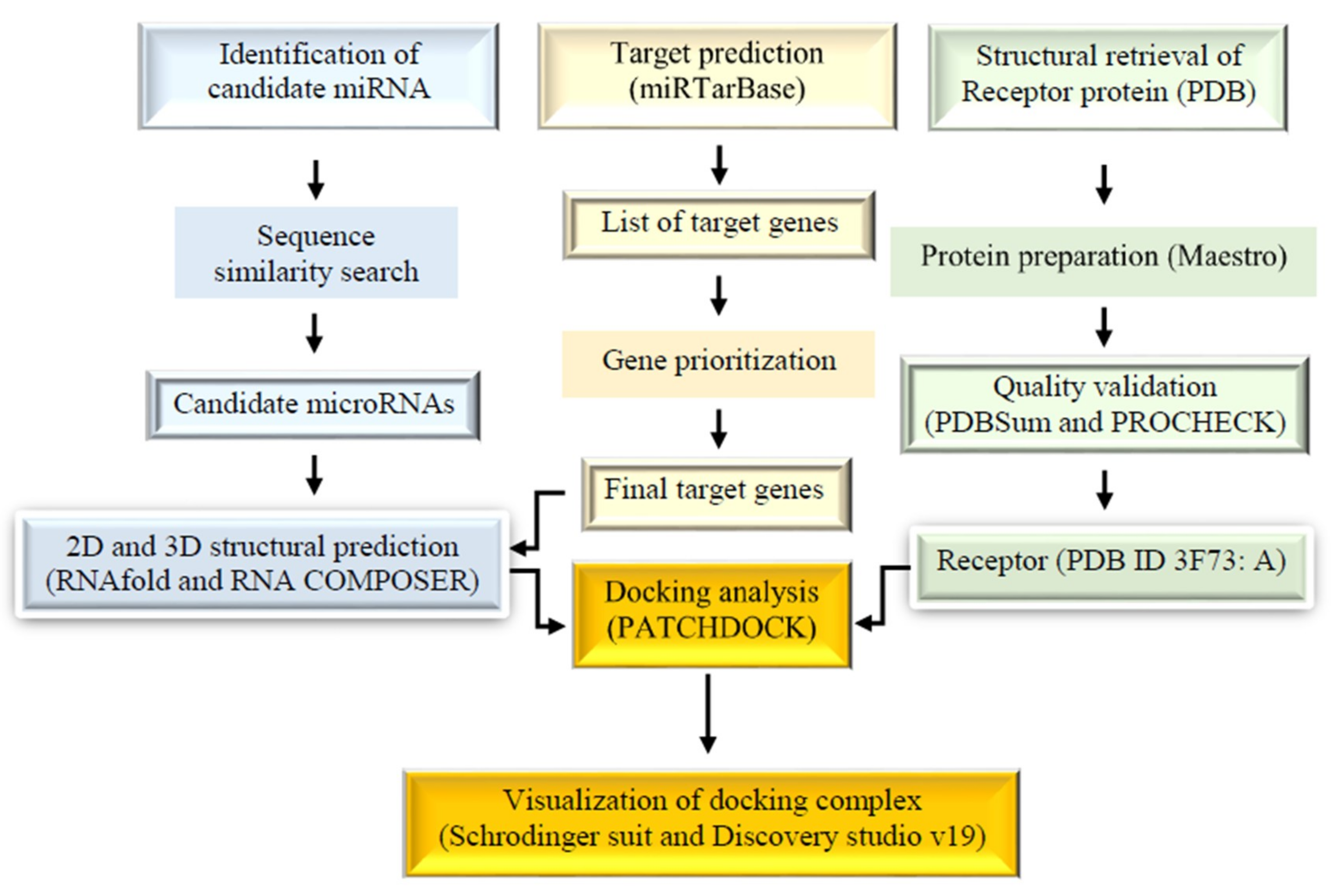
4. Materials and Methods
4.1. MicroRNA Identification
4.2. Target Prediction and Correlation to CRC
4.3. Structural Prediction of Candidate MicroRNA and Target Complexes
4.4. Protein Selection and Preparation
4.5. Molecular Docking
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
Abbreviations
| AGO | Argonaute (Receptor) |
| miRNA | microRNA |
| mRNA | Target genes |
| AA | Amino acid |
| H-bond | Hydrogen bond |
| PDB | protein data bank |
| RISC | RNA induced silencing complex |
| NA | Nucleic acid |
| BE | Binding energy |
| MFE | Minimum folding energy |
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA: A Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkenkamp-Demtroder, K.; Christensen, L.L.; Olesen, S.H.; Frederiksen, C.M.; Laiho, P.; Aaltonen, L.A.; Laurberg, S.; Sørensen, F.B.; Hagemann, R.; Ørntoft, T.F. Gene expression in colorectal cancer. Cancer Res. 2002, 62, 4352–4363. [Google Scholar] [PubMed]
- O’Keefe, S. Diet, microorganisms and their metabolites and colon cancer. Nature Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Kheirelseid, E.A.; Miller, N.; Chang, K.H.; Nugent, M.; Kerin, M.J. Clinical applications of gene expression in colorectal cancer. J. Gastrointest. Oncol. 2013, 4, 144. [Google Scholar] [PubMed]
- Fadaka, A.O.; Pretorius, A.; Klein, A. Biomarkers for Stratification in Colorectal Cancer: MicroRNAs. Cancer Control. 2019, 26, 1073274819862784. [Google Scholar] [CrossRef] [PubMed]
- Tuszynska, I.; Magnus, M.; Jonak, K.; Dawson, W.; Bujnicki, J.M. NPDock: A web server for protein–nucleic acid docking. Nucleic Acids Res. 2015, 43, W425–W430. [Google Scholar] [CrossRef]
- Hudson, W.H.; Ortlund, E.A. The structure, function and evolution of proteins that bind DNA and RNA. Nat. Rev. Mol. Cell Biol. 2014, 15, 749–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, T.A.; Wan, L.; Dreyfuss, G. RNA and Disease. Cell 2009, 136, 777–793. [Google Scholar] [CrossRef] [Green Version]
- Höck, J.; Meister, G. The Argonaute protein family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef]
- Schirle, N.T.; Kinberger, G.A.; Murray, H.F.; Lima, W.F.; Prakash, T.P.; MacRae, I.J. Structural analysis of human Argonaute-2 bound to a modified siRNA guide. J. Am. Chem. Soc. 2016, 138, 8694–8697. [Google Scholar] [CrossRef]
- Elkayam, E.; Kuhn, C.-D.; Tocilj, A.; Haase, A.D.; Greene, E.M.; Hannon, G.J.; Joshua-Tor, L. The structure of human argonaute-2 in complex with miR-20a. Cell 2012, 150, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Podder, S.; Ghosh, T.C. Insights into the miRNA regulations in human disease genes. BMC Genom. 2014, 15, 1010. [Google Scholar] [CrossRef] [PubMed]
- Silva Rodrigues, D.V.; Silva Monteiro, V.V.; Navegantes-Lima, K.C.; de Brito Oliveira, A.L.; de França Gaspar, S.L.; Gonçalves Quadros, L.B.; Monteiro, M.C. MicroRNAs in cell cycle progression and proliferation: Molecular mechanisms and pathways. Non-Coding RNA Investig. 2018. [Google Scholar] [CrossRef]
- Chivukula, R.R.; Shi, G.; Acharya, A.; Mills, E.W.; Zeitels, L.R.; Anandam, J.L.; Abdelnaby, A.A.; Balch, G.C.; Mansour, J.C.; Yopp, A.C. An essential mesenchymal function for miR-143/145 in intestinal epithelial regeneration. Cell 2014, 157, 1104–1116. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Olson, E.N.; Bassel-Duby, R. Mending broken hearts: Cardiac development as a basis for adult heart regeneration and repair. Nat. Rev. Mol. Cell Biol. 2013, 14, 529. [Google Scholar] [CrossRef] [PubMed]
- Hasuwa, H.; Ueda, J.; Ikawa, M.; Okabe, M. miR-200b and miR-429 function in mouse ovulation and are essential for female fertility. Science 2013, 341, 71–73. [Google Scholar] [CrossRef]
- Giza, D.E.; Vasilescu, C.; Calin, G.A. Key principles of miRNA involvement in human diseases. Discoveries 2014, 2, e34. [Google Scholar] [CrossRef]
- Moridikia, A.; Mirzaei, H.; Sahebkar, A.; Salimian, J. MicroRNAs: Potential candidates for diagnosis and treatment of colorectal cancer. J. Cell. Physiol. 2018, 233, 901–913. [Google Scholar] [CrossRef]
- Nishida, N.; Nagahara, M.; Sato, T.; Mimori, K.; Sudo, T.; Tanaka, F.; Shibata, K.; Ishii, H.; Sugihara, K.; Doki, Y. Microarray analysis of colorectal cancer stromal tissue reveals upregulation of two oncogenic miRNA clusters. Clin. Cancer Res. 2012, 18, 3054–3070. [Google Scholar] [CrossRef]
- Hirata, H.; Ueno, K.; Shahryari, V.; Tanaka, Y.; Tabatabai, Z.L.; Hinoda, Y.; Dahiya, R. Oncogenic miRNA-182–5p targets Smad4 and RECK in human bladder cancer. PLoS ONE 2012, 7, e51056. [Google Scholar] [CrossRef]
- Fadaka, A.O.; Ojo, B.A.; Adewale, O.B.; Esho, T.; Pretorius, A. Effect of dietary components on miRNA and colorectal carcinogenesis. Cancer Cell Int. 2018, 18, 130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yu, C.; Gao, H.; Li, Y. Argonaute proteins: Potential biomarkers for human colon cancer. BMC Cancer 2010, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, S.M.; Ardekani, G.S.; Ghaffari, M.; Martinka, M.; Li, G. Sox4-mediated Dicer expression is critical for suppression of melanoma cell invasion. Oncogene 2013, 32, 2131. [Google Scholar] [CrossRef] [PubMed]
- Völler, D.; Meister, G.; Bosserhoff, A. Strong reduction of Ago2 expression in melanoma and cellular consequences. Exp. Dermatol. 2014, 23. [Google Scholar] [CrossRef] [PubMed]
- Carmell, M.A.; Xuan, Z.; Zhang, M.Q.; Hannon, G.J. The Argonaute family: Tentacles that reach into RNAi, developmental control, stem cell maintenance, and tumorigenesis. Genes Dev. 2002, 16, 2733–2742. [Google Scholar] [CrossRef]
- Sun, R.; Gao, C.-L.; Li, D.-H.; Li, B.-J.; Ding, Y.-H. Expression status of PIWIL1 as a prognostic marker of colorectal cancer. Dis. Markers 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Völler, D.; Linck, L.; Bruckmann, A.; Hauptmann, J.; Deutzmann, R.; Meister, G.; Bosserhoff, A.K. Argonaute family protein expression in normal tissue and cancer entities. PLoS ONE 2016, 11, e0161165. [Google Scholar] [CrossRef]
- Bajorath, J. Integration of virtual and high-throughput screening. Nat. Rev. Drug Discov. 2002, 1, 882. [Google Scholar] [CrossRef]
- Demchenko, A.P. Recognition between flexible protein molecules: Induced and assisted folding. J. Mol. Recognit. 2001, 14, 42–61. [Google Scholar] [CrossRef]
- Du, X.; Li, Y.; Xia, Y.-L.; Ai, S.-M.; Liang, J.; Sang, P.; Ji, X.-L.; Liu, S.-Q. Insights into protein–ligand interactions: Mechanisms, models, and methods. Int. J. Mol. Sci. 2016, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ye, Q.; Du, Z.; Huang, M.; Zhang, M.; Tan, H. MiR-148b-3p inhibits renal carcinoma cell growth and pro-angiogenic phenotype of endothelial cell potentially by modulating FGF2. Biomed. Pharmacother. 2018, 107, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, L.; Liu, W.; Li, F. MicroRNA-497-5p inhibits proliferation and invasion of non-small cell lung cancer by regulating FGF2. Oncol. Lett. 2019, 17, 3425–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, X.; Zhang, J.; Liu, S.; Lou, X.; Liao, D.J. The other side of the coin: The tumor-suppressive aspect of oncogenes and the oncogenic aspect of tumor-suppressive genes, such as those along the CCND–CDK4/6–RB axis. Cell Cycle 2014, 13, 1677–1693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hunter, T. Roles of Chk1 in cell biology and cancer therapy. Int. J. Cancer 2014, 134, 1013–1023. [Google Scholar] [CrossRef]
- Gali-Muhtasib, H.; Ocker, M.; Kuester, D.; Krueger, S.; El-Hajj, Z.; Diestel, A.; Evert, M.; El-Najjar, N.; Peters, B.; Jurjus, A. Thymoquinone reduces mouse colon tumor cell invasion and inhibits tumor growth in murine colon cancer models. J. Cell. Mol. Med. 2008, 12, 330–342. [Google Scholar] [CrossRef]
- Wallin, Å.; Francis, P.; Nilbert, M.; Svanvik, J.; Sun, X.-F. Gene expression profile of colon cancer cell lines treated with SN-38. Chemotherapy 2010, 56, 17–25. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, H.; Hu, W. The regulation of MDM2 oncogene and its impact on human cancers. Acta Biochim Biophys Sin. (Shanghai) 2014, 46, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Shen, Z.-L.; Gao, Z.-D.; Zhao, G.; Wang, C.-Y.; Yang, Y.; Zhang, J.-Z.; Yan, Y.-C.; Shen, C.; Jiang, K.-W. MiR-194, commonly repressed in colorectal cancer, suppresses tumor growth by regulating the MAP4K4/c-Jun/MDM2 signaling pathway. Cell Cycle 2015, 14, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, J.; Wang, X.; Wu, R.; Lin, M.; Laddha, S.V.; Yang, Q.; Chan, C.S.; Feng, Z. MicroRNA-339-5p inhibits colorectal tumorigenesis through regulation of the MDM2/p53 signaling. Oncotarget 2014, 5, 9106–9117. [Google Scholar] [CrossRef]
- Kaindl, U.; Morak, M.; Portsmouth, C.; Mecklenbräuker, A.; Kauer, M.; Zeginigg, M.; Attarbaschi, A.; Haas, O.; Panzer-Grümayer, R. Blocking ETV6/RUNX1-induced MDM2 overexpression by Nutlin-3 reactivates p53 signaling in childhood leukemia. Leukemia 2014, 28, 600. [Google Scholar] [CrossRef] [PubMed]
- Sporn, J.C.; Hothorn, T.; Jung, B. BARD1 expression predicts outcome in colon cancer. Clin. Cancer Res. 2011, 17, 5451–5462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Pilyugin, M.; Kuester, D.; Leoni, V.P.; Li, L.; Casula, G.; Zorcolo, L.; Schneider-Stock, R.; Atzori, L.; Irminger-Finger, I. Expression of oncogenic BARD1 isoforms affects colon cancer progression and correlates with clinical outcome. Br. J. Cancer 2012, 107, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irminger-Finger, I.; Pilyugin, M. Regulation of BARD1 expression by non-coding RNA. Google Patents US10273475B2, 2019. [Google Scholar]
- Bach, D.-H.; Park, H.J.; Lee, S.K. The Dual Role of Bone Morphogenetic Proteins in Cancer. Mol. Oncolytics 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Chai, H. Inhibition of BAMBI reduces the viability and motility of colon cancer via activating TGF-β/Smad pathway in vitro and in vivo. Oncol. Lett. 2017, 14, 4793–4799. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Gupta Vallur, P.; Phaëton, R.; Mythreye, K.; Hempel, N. Insights into the Dichotomous Regulation of SOD2 in Cancer. Antioxidants 2017, 6, 86. [Google Scholar] [CrossRef]
- Zhang, Z.; Lang, J.; Cao, Z.; Li, R.; Wang, X.; Wang, W. Radiation-induced SOD2 overexpression sensitizes colorectal cancer to radiation while protecting normal tissue. Oncotarget 2017, 8, 7791. [Google Scholar] [CrossRef] [PubMed]
- Trimmer, C.; Sotgia, F.; Whitaker-Menezes, D.; Balliet, R.M.; Eaton, G.; Martinez-Outschoorn, U.E.; Pavlides, S.; Howell, A.; Iozzo, R.V.; Pestell, R.G.; et al. Caveolin-1 and mitochondrial SOD2 (MnSOD) function as tumor suppressors in the stromal microenvironment: A new genetically tractable model for human cancer associated fibroblasts. Cancer Biol. Ther. 2011, 11, 383–394. [Google Scholar] [CrossRef]
- Feng, H.; Gu, Z.-Y.; Li, Q.; Liu, Q.-H.; Yang, X.-Y.; Zhang, J.-J. Identification of significant genes with poor prognosis in ovarian cancer via bioinformatical analysis. J. Ovarian Res. 2019, 12, 35–35. [Google Scholar] [CrossRef]
- Wenzel, E.S.; Singh, A.T. Cell-cycle checkpoints and aneuploidy on the path to cancer. In Vivo 2018, 32, 1–5. [Google Scholar]
- Arthurs, C.; Murtaza, B.N.; Thomson, C.; Dickens, K.; Henrique, R.; Patel, H.R.; Beltran, M.; Millar, M.; Thrasivoulou, C.; Ahmed, A. Expression of ribosomal proteins in normal and cancerous human prostate tissue. PLoS ONE 2017, 12, e0186047. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Lu, Y.; Qin, H.; Zhou, Y.; Gu, Y.; Zhou, J.; Wang, X.; Fan, D. High Ran level is correlated with poor prognosis in patients with colorectal cancer. Int. J. Clin. Oncol. 2013, 18, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Flossbach, L.; Holzmann, K.; Mattfeldt, T.; Buck, M.; Lanz, K.; Held, M.; Möller, P.; Barth, T.F. High-resolution genomic profiling reveals clonal evolution and competition in gastrointestinal marginal zone B-cell lymphoma and its large cell variant. Int. J. Cancer 2013, 132, E116–E127. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Han, C.; Feng, J. Relationship of the expression levels of XIAP and p53 genes in hepatocellular carcinoma and the prognosis of patients. Oncol. Lett. 2017, 14, 4037–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, R.C.; Dou, Y. Hijacked in cancer: The KMT2 (MLL) family of methyltransferases. Nat. Rev. Cancer 2015, 15, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, D.; Hu, T.; Zhao, H.; Zhao, X.; Lou, Z.; He, Y.; Qin, W.; Xia, J.; Zhang, X. KMT2A histone methyltransferase contributes to colorectal cancer development by promoting cathepsin Z transcriptional activation. Cancer Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Badr, E.A.E.; Ali Assar, M.F.; Gohar, S.F.; Badr, M.H.; Hathout, R.M.; El-Kousy, S.M. The clinical impact of miRNA34a and P53 gene expression in colon cancer. Biochem. Biophys. Rep. 2018, 16, 88–95. [Google Scholar] [CrossRef]
- Okuda, H.; Toyota, M.; Ishida, W.; Furihata, M.; Tsuchiya, M.; Kamada, M.; Tokino, T.; Shuin, T. Epigenetic inactivation of the candidate tumor suppressor gene HOXB13 in human renal cell carcinoma. Oncogene 2006, 25, 1733–1742. [Google Scholar] [CrossRef]
- Maia, S.; Cardoso, M.; Pinto, P.; Pinheiro, M.; Santos, C.; Peixoto, A.; Bento, M.J.; Oliveira, J.; Henrique, R.; Jerónimo, C.; et al. Identification of Two Novel HOXB13 Germline Mutations in Portuguese Prostate Cancer Patients. PLoS ONE 2015, 10, e0132728. [Google Scholar] [CrossRef]
- Xue, X.; Taylor, M.; Anderson, E.; Hao, C.; Qu, A.; Greenson, J.K.; Zimmermann, E.M.; Gonzalez, F.J.; Shah, Y.M. Hypoxia-inducible factor-2α activation promotes colorectal cancer progression by dysregulating iron homeostasis. Cancer Res. 2012, 72, 2285–2293. [Google Scholar] [CrossRef]
- Heo, S.J.; Lee, C.-K.; Hahn, K.Y.; Kim, G.; Hur, H.; Choi, S.H.; Han, K.S.; Cho, A.; Jung, M. A Case of von Hippel-Lindau Disease with Colorectal Adenocarcinoma, Renal Cell Carcinoma and Hemangioblastomas. Cancer Res. Treat. 2016, 48, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Popenda, M.; Szachniuk, M.; Antczak, M.; Purzycka, K.J.; Lukasiak, P.; Bartol, N.; Blazewicz, J.; Adamiak, R.W. Automated 3D structure composition for large RNAs. Nucleic Acids Res. 2012, 40, e112–e112. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, G.; Sharma, P.; Anant, A.; Deshmukh, S.; Kaushik, H.; Gopal, K.; Srivastava, N.; Sharma, N.; Garg, L.C. Identification and modeling of a drug target for Clostridium perfringens SM101. Bioinformation 2010, 4, 278. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.; Singh, H.; Srivastava, H.K.; Singh, S.; Kishore, G.; Raghava, G.P. Benchmarking of different molecular docking methods for protein-peptide docking. BMC Bioinform. 2019, 19, 426. [Google Scholar] [CrossRef] [PubMed]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef] [PubMed]
- Wade, R.C.; Goodford, P.J. The role of hydrogen-bonds in drug binding. Prog. Clin. Biol. Res. 1989, 289, 433–444. [Google Scholar]
- Schiebel, J.; Gaspari, R.; Wulsdorf, T.; Ngo, K.; Sohn, C.; Schrader, T.E.; Cavalli, A.; Ostermann, A.; Heine, A.; Klebe, G. Intriguing role of water in protein-ligand binding studied by neutron crystallography on trypsin complexes. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Tüfekci, K.U.; Öner, M.G.; Meuwissen, R.L.J.; Genç, Ş. The role of microRNAs in human diseases. In miRNomics: MicroRNA Biology and Computational Analysis; Springer: Berlin, Germany, 2014; pp. 33–50. [Google Scholar]
- Ha, T.-Y. MicroRNAs in human diseases: From cancer to cardiovascular disease. Immune Netw. 2011, 11, 135–154. [Google Scholar] [CrossRef]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med Biotechnol. 2010, 2, 161–179. [Google Scholar]
- Zhou, S.-S.; Jin, J.-P.; Wang, J.-Q.; Zhang, Z.-G.; Freedman, J.H.; Zheng, Y.; Cai, L. miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta Pharmacol. Sin. 2018, 39, 1073. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.C.P.D.; Carneiro, F.D.; Almeida, K.C.D.; Fernandes-Santos, C. Role of miRNAs on the Pathophysiology of Cardiovascular Diseases. Arq. Bras. De Cardiol. 2018, 111, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Liu, H.; Zhang, L.; Lv, W.; Hu, X. Altered microRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 2015, 6, 37043. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Cho, S.K.; Thulstrup, P.W.; Bjerrum, M.J.; Lee, P.H.; Kang, J.-H.; Bhang, Y.-J.; Yang, S.W. MicroRNA biomarkers in neurodegenerative diseases and emerging nanosensors technology. J. Mov. Disord. 2017, 10, 18. [Google Scholar] [CrossRef]
- Rajgor, D. Macro roles for microRNAs in neurodegenerative diseases. Non-Coding RNA Res. 2018, 3, 154–159. [Google Scholar] [CrossRef] [PubMed]
- James, A.B.; Fadaka, A.O.; Magbagbeola, O.A.; Oturu, A.; KOLAWOLE, O.O.; Ogunjimi, A.; Oshodi, T.; Habeebu, M.; Onawoga, F.O.; Ajogbeje, E.O. Haematological and miRNAs (let-7g, miR-21, miR-141) expression modulation profile in serum samples of Human prostate cancer. FASEB J. 2017, 31, 712–757. [Google Scholar]
- Ryan, B.M.; Robles, A.I.; Harris, C.C. Genetic variation in microRNA networks: The implications for cancer research. Nat. Rev. Cancer 2010, 10, 389. [Google Scholar] [CrossRef]
- Luo, X.; Burwinkel, B.; Tao, S.; Brenner, H. MicroRNA signatures: Novel biomarker for colorectal cancer? Cancer Epidemiol. Prev. Biomark. 2011, 20, 1272–1286. [Google Scholar] [CrossRef]
- Madhavan, D.; Cuk, K.; Burwinkel, B.; Yang, R. Cancer diagnosis and prognosis decoded by blood-based circulating microRNA signatures. Front. Genet. 2013, 4, 116. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Feng, Y.; Coukos, G.; Zhang, L. Therapeutic microRNA strategies in human cancer. AAPS J. 2009, 11, 747. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-S.; Xiao, H.-S. MicroRNAs as potential biomarkers for gastric cancer. World J. Gastroenterol. 2014, 20, 12007–12017. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Leslie, C.S. Learning to predict miRNA-mRNA interactions from AGO CLIP sequencing and CLASH data. PLoS Comput. Biol. 2016, 12, e1005026. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Hamada, J.-I.; Takada, M.; Asano, T.; Murakawa, K.; Takahashi, Y.; Murai, T.; Tada, M.; Miyamoto, M.; Kondo, S. Aberrant expressions of HOX genes in colorectal and hepatocellular carcinomas. Oncol. Rep. 2010, 23, 843–851. [Google Scholar]
- Zacchetti, G.; Duboule, D.; Zakany, J. Hox gene function in vertebrate gut morphogenesis: The case of the caecum. Development 2007, 134, 3967–3973. [Google Scholar] [CrossRef] [PubMed]
- Tatangelo, F.; Di Mauro, A.; Scognamiglio, G.; Aquino, G.; Lettiero, A.; Delrio, P.; Avallone, A.; Cantile, M.; Botti, G. Posterior HOX genes and HOTAIR expression in the proximal and distal colon cancer pathogenesis. J. Transl. Med. 2018, 16, 350. [Google Scholar] [CrossRef]
- Sekiya, T.; Adachi, S.; Kohu, K.; Yamada, T.; Higuchi, O.; Furukawa, Y.; Nakamura, Y.; Nakamura, T.; Tashiro, K.; Kuhara, S. Identification of BMP and activin membrane-bound inhibitor (BAMBI), an inhibitor of transforming growth factor-β signaling, as a target of the β-catenin pathway in colorectal tumor cells. J. Biol. Chem. 2004, 279, 6840–6846. [Google Scholar] [CrossRef]
- Togo, N.; Ohwada, S.; Sakurai, S.; Toya, H.; Sakamoto, I.; Yamada, T.; Nakano, T.; Muroya, K.; Takeyoshi, I.; Nakajima, T. Prognostic significance of BMP and activin membrane-bound inhibitor in colorectal cancer. World J. Gastroenterol. 2008, 14, 4880. [Google Scholar] [CrossRef]
- Tang, J.; Gifford, C.; Samarakoon, R.; Higgins, P. Deregulation of negative controls on TGF-β1 signaling in tumor progression. Cancers 2018, 10, 159. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Joshi, S.; Brasky, J.T.; DeLaCruz, M.; Gibson, T.P.; Kunte, D.; Subramanian, P.; Wali, R.K.; Roy, H.K. Superoxide Dismutase 2 (Sod2) is Overexpressed at an Early Stage During Colorectal Carcinogenesis: A Putative Target for Celecoxib Chemoprevention. Gastroenterology 2011, 140, 343. [Google Scholar] [CrossRef]
- Irminger-Finger, I.; Ratajska, M.; Pilyugin, M. New concepts on BARD1: Regulator of BRCA pathways and beyond. Int. J. Biochem. Cell Biol. 2016, 72, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.J.; Li, H.-G.; Tang, E.-J.; Wu, W.; Chen, Y.; Jiang, H.-H.; Lin, M.-B.; Yin, L. PIK3CA and TP53 mutations predict overall survival of stage II/III colorectal cancer patients. World J. Gastroenterol. 2018, 24, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.; Craik, D.; Martin, J. Functional group contributions to drug-receptor interactions. J. Med. Chem. 1984, 27, 1648–1657. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.N.; Das, D.; Konkimalla, V.B.; Pradhan, S.K. In silico study of miRNA based gene regulation, involved in solid cancer, by the assistance of argonaute protein. Genom. Inform. 2016, 14, 112. [Google Scholar] [CrossRef] [PubMed]
- Alex, A.; Millan, D.S.; Perez, M.; Wakenhut, F.; Whitlock, G.A. Intramolecular hydrogen bonding to improve membrane permeability and absorption in beyond rule of five chemical space. MedChemComm 2011, 2, 669–674. [Google Scholar] [CrossRef]
- Kuhn, B.; Mohr, P.; Stahl, M. Intramolecular hydrogen bonding in medicinal chemistry. J. Med. Chem. 2010, 53, 2601–2611. [Google Scholar] [CrossRef]
- Sahu, V.; Khan, A.; Singh, R.; Singh, P. Hydrophobic, polar and hydrogen bonding based drug-receptor interaction of tetrahydroimidazobenzodiazepinones. Am. J. Immunol 2008, 4, 33. [Google Scholar] [CrossRef]
- Hsu, S.-D.; Lin, F.-M.; Wu, W.-Y.; Liang, C.; Huang, W.-C.; Chan, W.-L.; Tsai, W.-T.; Chen, G.-Z.; Lee, C.-J.; Chiu, C.-M. miRTarBase: A database curates experimentally validated microRNA–target interactions. Nucleic Acids Res. 2010, 39, D163–D169. [Google Scholar] [CrossRef]
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2017, 46, D296–D302. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2008, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candidate miRNA | Validated microRNA | Fasta Sequences |
|---|---|---|
| miR-1 | hsa-miR-193a-5p | >hsa-miR-193a-5p MIMAT0004614 UGGGUCUUUGCGGGCGAGAUGA |
| miR-2 | hsa-miR-450b-3p | >hsa-miR-450b-3p MIMAT0004910 UUGGGAUCAUUUUGCAUCCAUA |
| miR-3 | hsa-miR-501-3p | >hsa-miR-501-3p MIMAT0004774 AAUGCACCCGGGCAAGGAUUCU |
| miR-4 | hsa-miR-501-3p | >hsa-miR-501-3p MIMAT0004774 AAUGCACCCGGGCAAGGAUUCU |
| miR-5 | hsa-miR-513a-3p | >hsa-miR-513a-3p MIMAT0004777 UAAAUUUCACCUUUCUGAGAAGG |
| miR-1 | MFE | miR-2 | MFE | miR-3 | MFE | miR-4 | MFE | miR-5 | MFE |
|---|---|---|---|---|---|---|---|---|---|
| A1CF | −19.10 | BAMBI | −9.80 | SOD2 | −16.20 | BARD1 | −21.30 | PDCD4 | −11.80 |
| PAQR3 | −13.80 | XIAP | −8.70 | PAQR3 | −10.80 | SLC1A5 | −17.30 | VMP1 | −12.50 |
| STMN1 | −19.90 | BMP2 | −8.70 | SLC7A11 | −20.60 | WT1 | −14.80 | CDK4 | −10.70 |
| MACC1 | −18.00 | ZNF703 | −13.80 | MDM2 | −11.90 | CLMN | −16.40 | TP53 | −10.70 |
| FGB | −12.90 | PPM1D | −16.90 | RAN | −14.70 | REL | −19.80 | CHEK1 | −8.70 |
| HOXB13 | −23.90 | BUB1 | −8.00 | LAMB1 | −11.52 | HDGF | −21.70 | H2AFZ | −9.60 |
| ALDOA | −19.20 | LYN | −12.90 | ORAI2 | −19.50 | RNF138 | −18.20 | ||
| CHAC1 | −20.10 | KLF8 | −11.02 | VAV3 | −17.80 | SLC7A5 | −12.50 | ||
| GSTK1 | −18.10 | FGF2 | −14.60 | ||||||
| RPS19 | −19.10 | KMT2A | −17.02 | ||||||
| CRKL | −15.40 | ||||||||
| VHL | −19.90 |
| Gene | Function | miRNA | MFE |
|---|---|---|---|
| TP53 | Cell cycle, Apoptosis, Cell proliferation, others | miR-5 | −10.70 |
| FGF2 | Angiogenesis, Cell proliferation, others | miR-2 | −14.60 |
| CHEK1 | Cell cycle, Apoptosis, other functions | miR-5 | −8.70 |
| WT1 | Apoptosis, Cell proliferation, others | miR-4 | −14.80 |
| MDM2 | Cell cycle, Cell proliferation, others | miR-3 | −11.90 |
| BARD1 | Cell cycle, Apoptosis, others | miR-4 | −21.30 |
| BUB1 | Cell cycle, others | miR-2 | −8.00 |
| XIAP | Apoptosis, others | miR-2 | −8.70 |
| BMP2 | Cell proliferation, others | miR-2 | −8.70 |
| CDK4 | Cell cycle, others | miR-5 | −10.70 |
| HOXB13 | Angiogenesis, others | miR-1 | −23.90 |
| KMT2A | Apoptosis, others | miR-2 | −17.02 |
| VHL | Angiogenesis, others | miR-1 | −19.90 |
| BAMBI | Other functions | miR-2 | −9.80 |
| RAN | Other functions | miR-3 | −14.70 |
| REL | Other functions | miR-4 | −19.80 |
| RPS19 | Other functions | miR-1 | −19.10 |
| SOD2 | Other functions | miR-3 | −16.20 |
| S/N | miRNAs | Target Gene |
|---|---|---|
| 1 | miR-1 | HOXB13 |
| 2 | miR-2 | BAMBI |
| 3 | miR-3 | SOD2 |
| 4 | miR-4 | BARD1 |
| 5 | miR-5 | TP53 |
| Gene | miRNA | Dot-Bracket Notation | 2° Structure of Duplex | BE | MFE |
|---|---|---|---|---|---|
| HOXB13 | miR-1 | .......((((..((((((((........))))))))..)))) |  | −13.3 | −23.9 |
| BAMBI | miR-2 | ............(((((.((((.((.....)).))))..)))))....... |  | −2.3 | −9.6 |
| SOD2 | miR-3 | ...((((..(((......))).)))).((.(((......))).))... |  | −8.5 | −16.2 |
| BARD1 | miR-4 | .............((((((((((....)))))...))))).. |  | −12.8 | −21.3 |
| TP53 | miR-5 | .(((((...)))))..(((...(((...)))....)))... |  | −4.0 | −10.7 |
| miRNA-mRNA and AGO | Score | Area | ACE |
|---|---|---|---|
| miR-1 -AGO | 19544 | 3390.80 | −258.22 |
| miR-2-AGO | 18618 | 2832.70 | −22.43 |
| miR-3-AGO | 18420 | 2814.10 | −151.43 |
| miR-4-AGO | 18024 | 2344.20 | −131.18 |
| miR-5-AGO | 20.372 | 2913.20 | −488.07 |
| miRNA | Hydrophobic AA | Aromatic AA | H-Bond |
|---|---|---|---|
| (21 a), LEU45 d, ALA47 d, VAL58 d, VAL108 d, ALA111 d, LEU112 d, VAL129 d, LEU132 e, ALA133 e, LEU217 d, ALA245 d, ILE254 d, VAL264 d, LEU596 d | (7b), TYR43 d, TYR135 d, TRP156 e, TRP202 d | (25 c) ARG114 d, ARG574 d, GLY577 d, LYS248 d, ASP246 d, ASP154 d, ARG200 d, GLY131 d, PRO103 d, LEU153 d | |
| miR-2 | (20 a), ALA47 d, VAL58 d, VAL108 d, ALA111 d, LEU112 d, LEU132 e, ALA133 e, VAL152 d, LEU153 d, LEU217 d, ALA245 d, ILE254 d, VAL264 d, VAL549d d, LEU596 d, VAL620 d | (3 b), TYR43 d, TRP156 e, TRP202 d | (21 c) ARG114 d, ARG574 d, GLY577 d, LYS248 d, ASP246 d, ARG548 d, GLU483 d, SER576 d, ARG192 d, LYS599 d, ARG81 d |
| miR-3 | (27 a), ALA47 d, VAL58 d, LEU64 d, VAL108 d, ALA111 d, LEU112 d, VAL129 d, LEU132 e, ALA133 e, VAL152 d, LEU153 d, ALA450 d, ALA479 d, VAL549 d, VAL620 d, LEU652 d, VAL663 d | (6 b), TYR43 d, TRP156 e, TRP447 d | (26 c) ARG114, ARG574, GLY577, ASP154, ARG548 GLU483, LYS664 ARG661, ARG200 GLY131 PRO103, LYS599 ARG81, ASP660, |
| miR-4 | (22 a), ALA47 d, LEU132 e, ALA133 e, ALA151 d, VAL152 d, LEU153 d, ALA170 d, ILE173 d, VAL264 d, LEU265 d, LEU267 d, LEU279 d, ALA479 d, VAL573 d, ALA648 d, LEU652 d, LEU662 d, VAL663 d | (4 b), TYR135 d, TRP156e, PHE649 d | (15 c) ARG114, LYS248, ARG548 GLU483, SER576 ARG192, LYS664 ARG661, LEU153, THR266 LYS575 ARG482 |
| miR-5 | (27 a), LEU132 e, ALA133 e, ALA151 d, VAL152d, LEU153 d, ALA170 d, ILE173 d, VAL264 d, LEU265 d, LEU267 d, LEU279 d, ALA450 d, ALA479 d, VAL549 d, VAL573 d, ALA648 d, LEU652 d, LEU662 d, VAL663 d | (7 b) TYR135 d, TRP156 e, TRP447 d, PHE649 d | (17 c) ARG574 d, ASP246 d, ASP154 d, SER576 d, ARG192 d, LYS664 d, ARG661 d, ASP660 d, THR266 d, LYS575 d, ARG482 d |
| microRNA | AA Residues | Atoms | Distance | NA Residues |
|---|---|---|---|---|
| miR-1 | GLN84 | HE21-OP1 | 1.8 | (G3) |
| ARG574 | HH11-O3’ | 1.7 | (G12) | |
| ALA111 | HA-O2’ | 1.9 | (G16) | |
| PRO36 | O-H4’ | 1.8 | (G2) | |
| ASP154 | OD1-H5’ | 2.0 | (A11) | |
| O-H4′ | 2.0 | (G2) | ||
| miR-2 | GLY104 | HA3-O6 | 1.8 | (G15) |
| ARG114 | HD3-O4’ | 1.6 | (G19) | |
| ARG574 | HD3-OP1 | 1.5 | (A10) | |
| GLU483 | OE1-H5 | 2.0 | (A8) | |
| ARG59 | O-H4’ | 1.9 | (A17) | |
| miR-3 | ARG548 | HH11-O2’ | 1.9 | (A17) |
| ARG574 | HH22-O4’ | 1.9 | (G) | |
| VAL129 | O-HO5’ | 1.8 | (A1) | |
| ASP154 | OD1-HO2’ | 1.6 | (A13) | |
| PRO44 | HA-O3’ | 2.0 | (C4) | |
| GLY577 | HA2-O2’ | 2.0 | (A14) | |
| ARG661 | HA-O2’ | 1.9 | (U19) | |
| GLU622 | OE1-H5’ | 1.8 | (G9) | |
| ASP660 | O-H2’ | 2.0 | (U19) | |
| miR-4 | ARG668 | HH12-O5’ | 1.8 | (G1) |
| ARG615 | HD2-OP2 | 2.0 | (A12) | |
| THR266 | OG1-H5’ | 2.0 | (G8) | |
| miR-5 | LYS575 | HZ1-O2 | 1.9 | (U6) |
| ARG661 | HE-O4’ | 1.9 | (U4) | |
| ARG574 | HD2-O4’ | 2.0 | (A8) | |
| SER576 | H-O2 | 2.0 | (C7) |
| miRNA-mRNA and AGO | Score | Area | ACE |
|---|---|---|---|
| miR-1-HOXB13-AGO | 24046 | 3962.90 | −851.20 |
| miR-2-BAMBI-AGO | 24380 | 5528.70 | −966.63 |
| miR-3-SOD2-AGO | 27570 | 3974.80 | −652.52 |
| miR-4-BARD1–AGO | 24816 | 3524.00 | −836.85 |
| miR-5-TP53-AGO | 23716 | 3402.30 | −547.97 |
| miRNA-mRNA | Residual Hydrophobic AA | Aromatic AA |
|---|---|---|
| miR1-HOXB13 | (34 a), VAL42 c, LEU45 d, LEU132 c, ALA133 c, VAL147 c, ALA151 c, VAL152 c, ALA170 c, ILE173 c, VAL264 c, LEU265 d, LEU267 d, LEU277 c, LEU279 c, LEU281 c, ALA331 c, ALA414 c, ILE434 c, ALA479 c, VAL549 c, VAL573 c, VAL606 c, LEU617 c, ALA644 c, LEU652 c, VAL663 c, VAL685 c | (8 b), TYR43 d, TYR135 c, TRP202 d, TRP415 c, TYR642 c, PHE647 c, PHE649 d |
| miR-2-BAMBI | (38 a), VAL42 c, LEU45 d, LEU46 c, ALA47 c, ALA50 c, VAL58 c, ALA111 c, LEU132 c, VAL264 c, LEU265 d, LEU267 d, LEU277 c, ALA331 c, ALA414 c, LEU435 c, LEU439 c, ALA479 c, VAL549 c, ALA648 c, VAL663 c. | (12 b), TYR43 d, TRP202 d, TRP415 c, TRP447 c, PHE487 c, PHE610 c, PHE647 c, PHE649 d |
| miR-3-SOD2 | (24 a), VAL42 c, LEU45 d, LEU46 c, ALA47 c, VAL58 c, ALA111 c, LEU132 c, ALA133 c, VAL152 c, ALA170 c, ILE173 c, LEU265 d, LEU267 d, LEU279 c, LEU281 c, VAL606 c, LEU617 c, ALA648 c, LEU652 c | (9 b), TYR43 d, TYR171 c, TRP202 d, TRP415 c, PHE487 c, PHE647 c, PHE649 d |
| miR-4-BARD1 | (23 a), VAL42 c, LEU45 d, VAL147 c, VAL152 c, ALA170 c, ILE173 c, VAL264 c, LEU265 d, LEU267 d, LEU277 c, ALA278 c, LEU279 c, LEU281 c, ILE434 c, LEU435 c, ALA450 c, VAL573 c, VAL606 c, ALA644 c, ALA648 c, LEU652 c, VAL685 c | (10 b), TYR43 d, TYR135 c, TYR171 c, TRP202 d, TRP447 c, PHE487 c, PHE610 c, TYR642 c, PHE649 d |
| miR-5-TP53 | (30 a), LEU45 d, ALA47 c, ALA50 c, VAL58 c, ALA111 c, LEU132 c, ALA133 c, VAL147 c, ALA151 c, VAL152 c, ALA170 c, ILE173 c, VAL264 c, LEU265 d, LEU267 d, LEU277 c, ALA278 c, LEU279 c, LEU281 c, ILE434 c, LEU435 c, LEU439 c, ALA450 c, VAL606 c, LEU617 c, ALA644 c, ALA648 c, LEU652 c, VAL685 c | (10 b), TYR43 d, TYR135 c, TYR171 c, TRP202 d, TRP447 c, PHE610 c, TYR642 c, PHE647 c, PHE649 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fadaka, A.O.; Pretorius, A.; Klein, A. MicroRNA Assisted Gene Regulation in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 4899. https://doi.org/10.3390/ijms20194899
Fadaka AO, Pretorius A, Klein A. MicroRNA Assisted Gene Regulation in Colorectal Cancer. International Journal of Molecular Sciences. 2019; 20(19):4899. https://doi.org/10.3390/ijms20194899
Chicago/Turabian StyleFadaka, Adewale O., Ashley Pretorius, and Ashwil Klein. 2019. "MicroRNA Assisted Gene Regulation in Colorectal Cancer" International Journal of Molecular Sciences 20, no. 19: 4899. https://doi.org/10.3390/ijms20194899
APA StyleFadaka, A. O., Pretorius, A., & Klein, A. (2019). MicroRNA Assisted Gene Regulation in Colorectal Cancer. International Journal of Molecular Sciences, 20(19), 4899. https://doi.org/10.3390/ijms20194899