Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H2O2-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury



Abstract
:1. Introduction
2. Results and Discussion
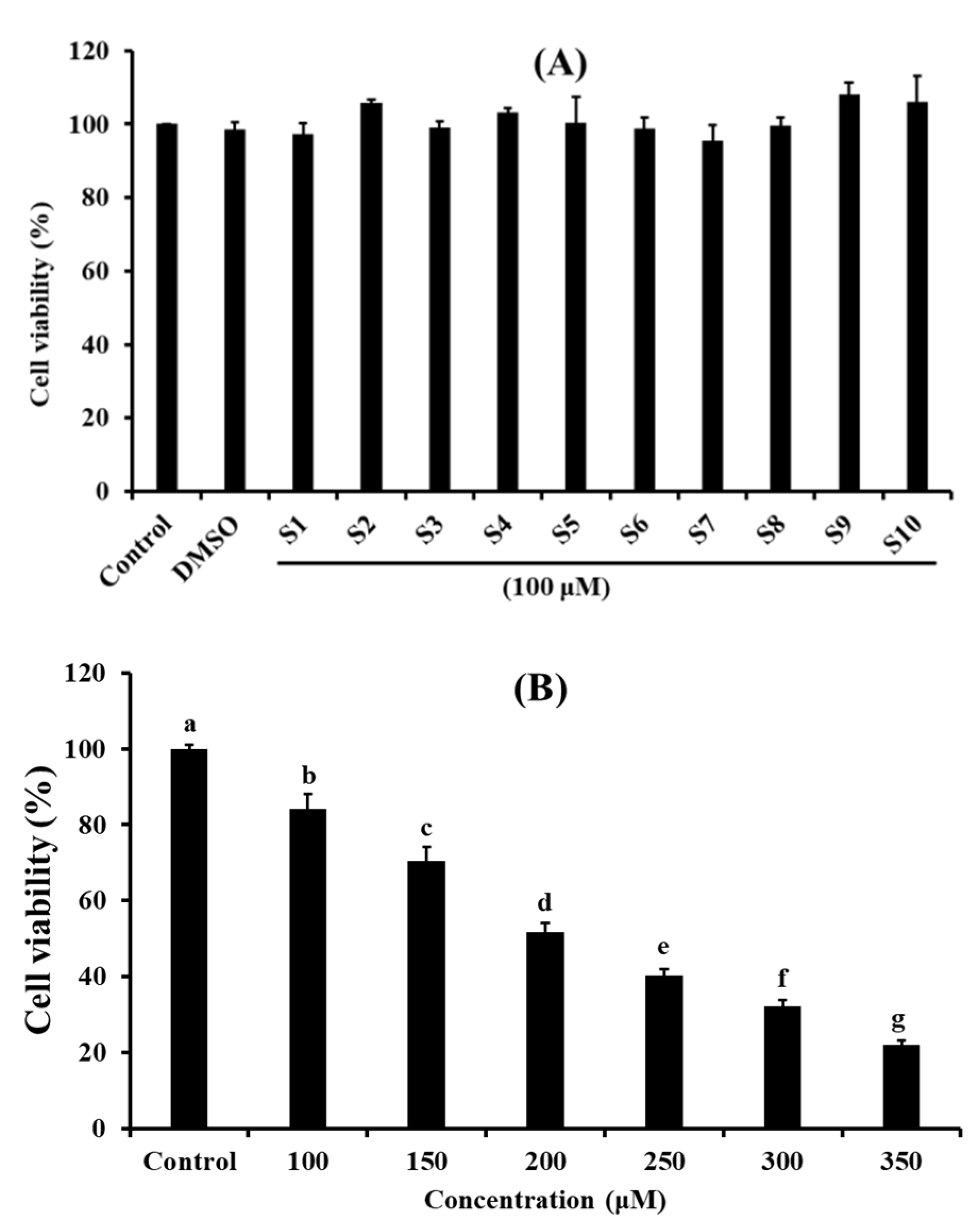
2.1. Effects of Antioxidant Pentapeptides (S1–S10) on the Viability of HUVEC
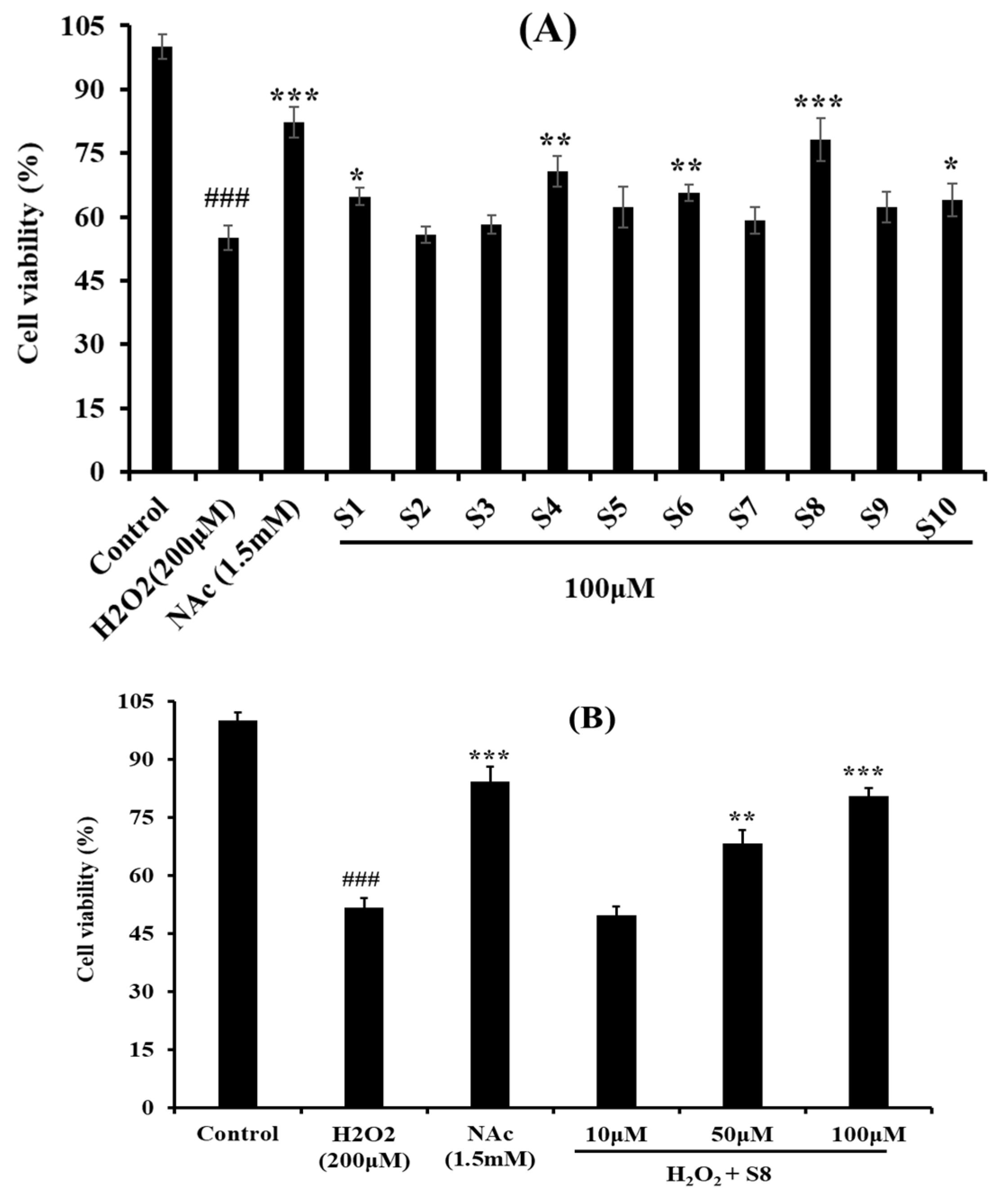
2.2. Protective Effect of Antioxidant Pentapeptides (S1–S10) on the Oxidative Injury HUVEC by H2O2
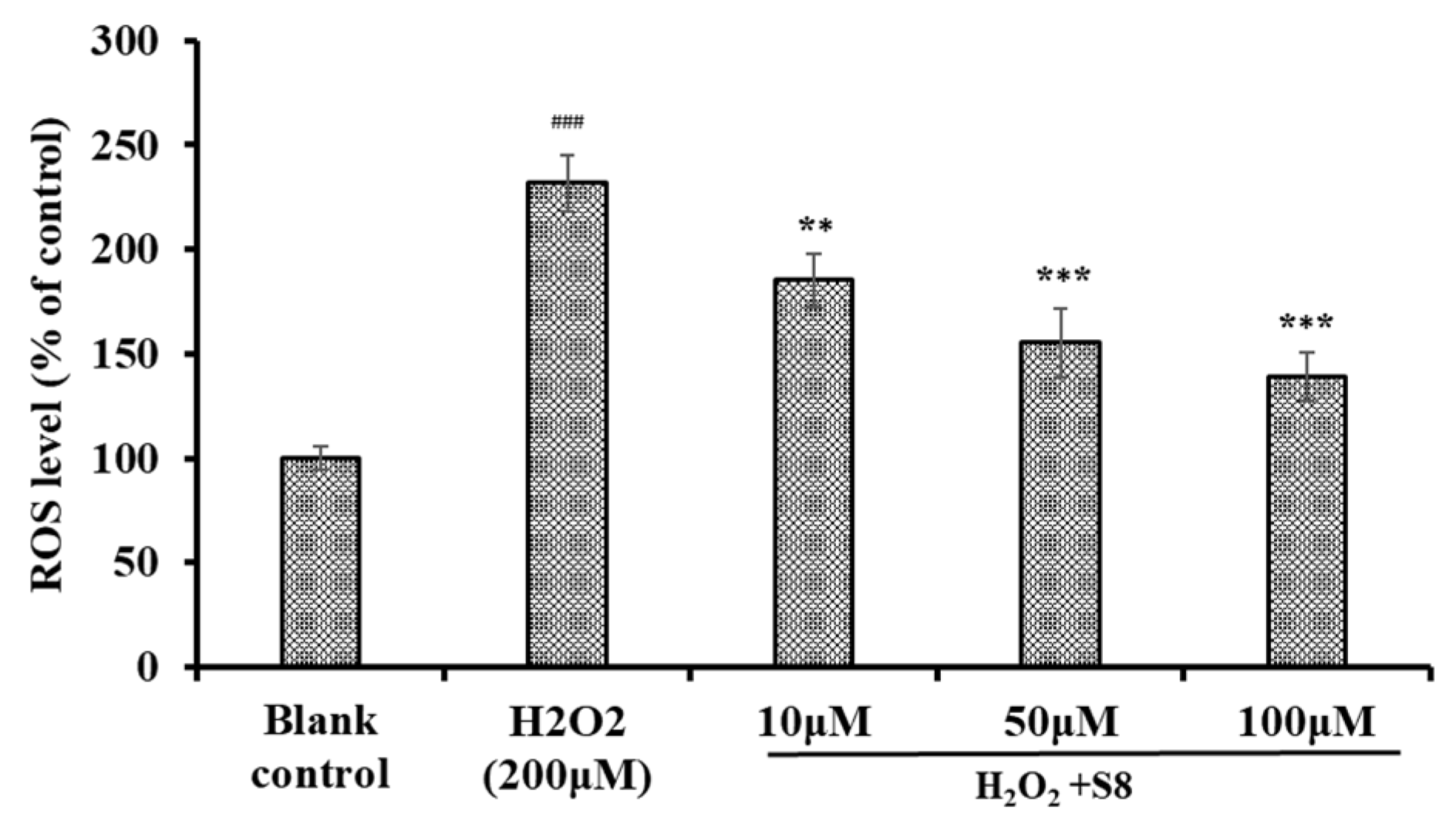
2.3. Effect of FPYLRH (S8) on the Levels of ROS in H2O2-Induced HUVEC Injury Model
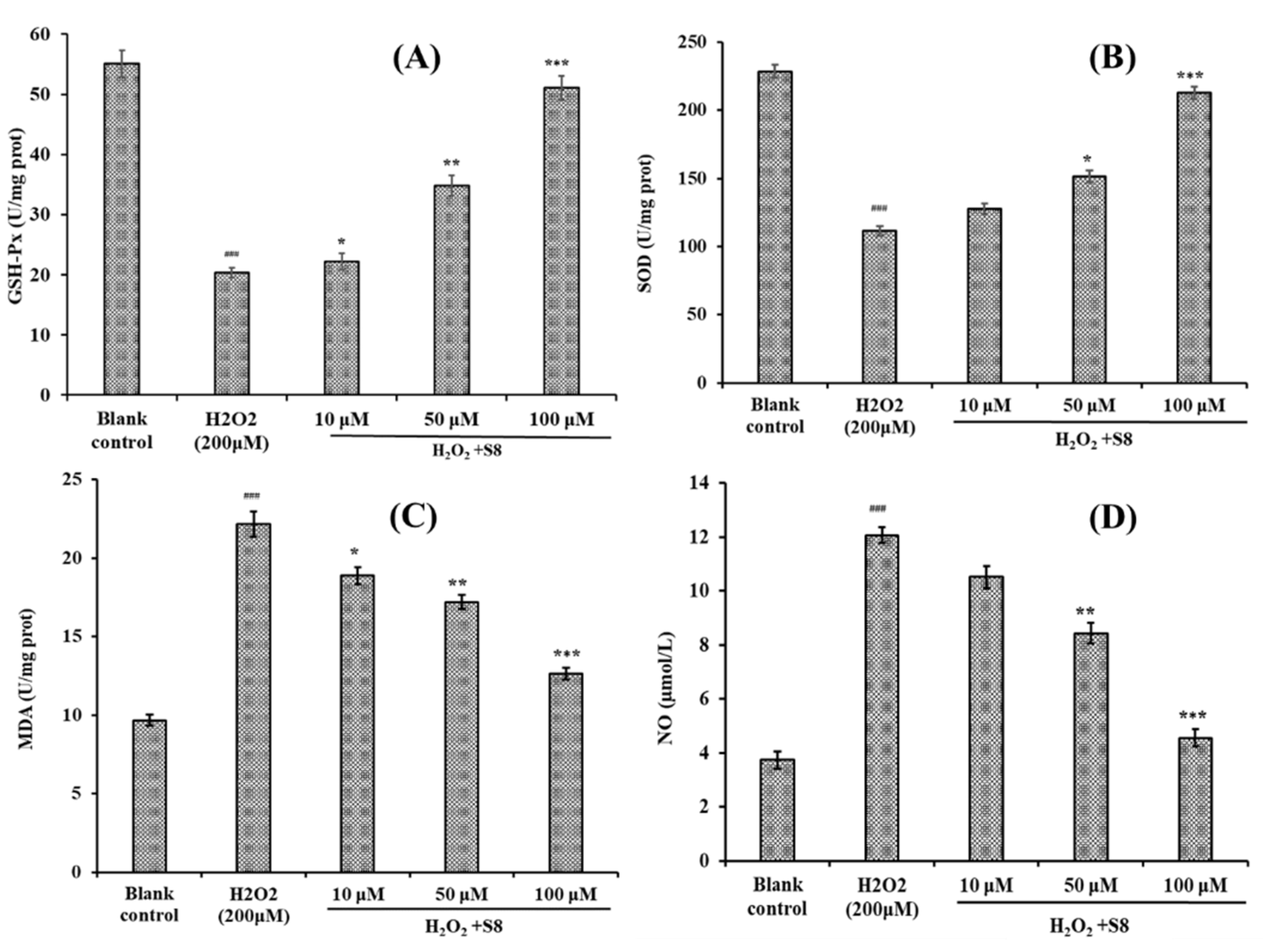
2.4. Effect of FPYLRH (S8) on the Levels of GSH-Px, SOD, Malondialdehyde (MDA) and Nitric Oxide (NO) in H2O2-Induced HUVEC Injury Model
2.5. Protective Activity of FPYLRH (S8) on Oxidative Damage DNA Induced by H2O2
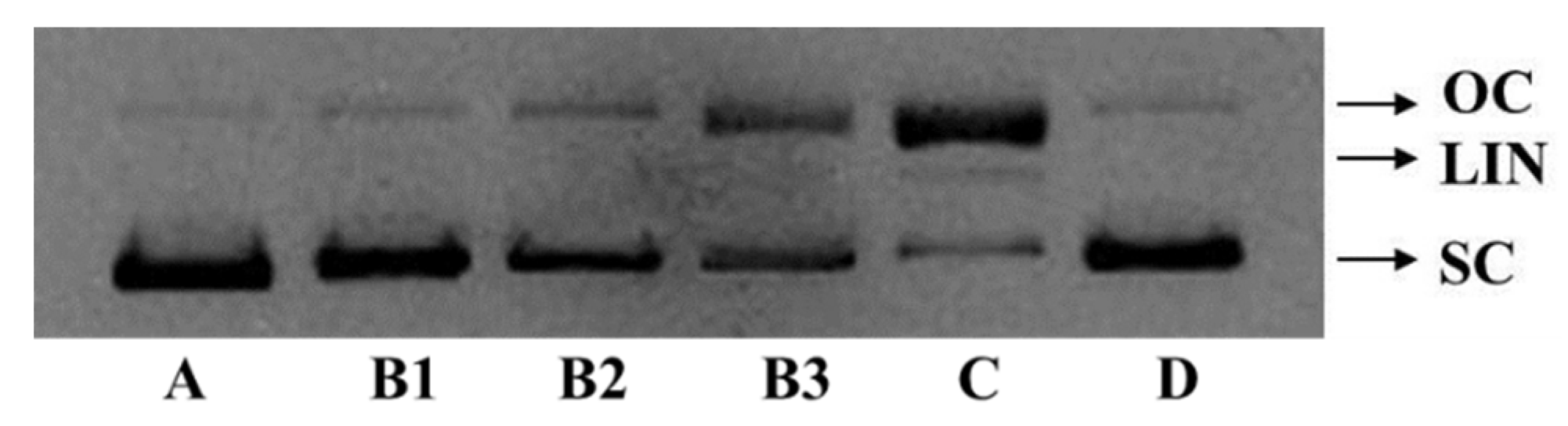
2.5.1. Protective Activity on Plasmid DNA (pBR322 DNA)
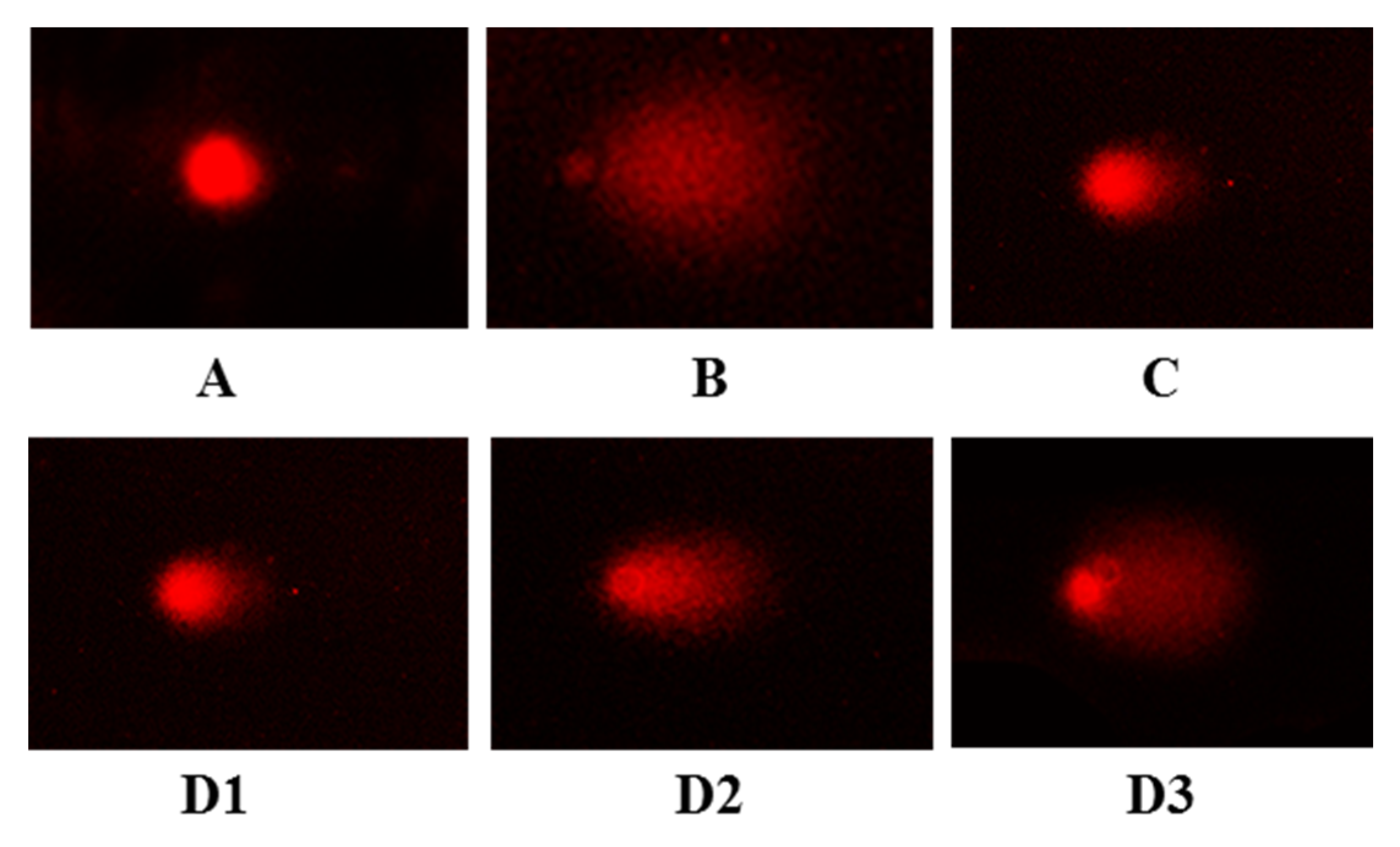
2.5.2. Protective Activity on DNA in H2O2-Induced HUVEC Injury Model
2.6. Relationship Among the Molecular Size, Amino Acid Composition, and Antioxidant Activity
3. Method and Materials
3.1. Materials
3.2. Cell Culture and Viability Assay
3.3. Protection of FPYLRH on Oxidative Damage HUVECs by H2O2
3.4. Hoechst 33,342 Staining Assay
3.5. Determination of the Levels of ROS in H2O2-Induced HUVECs
3.6. Determination of the Levels of Antioxidant Enzymes in H2O2-Induced HUVECs
3.7. Protective Effect of FPYLRH on pBR322 Plasmid DNA Damaged by H2O2
3.8. DNA Comet Assay
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhao, W.H.; Luo, Q.B.; Pan, X.; Chi, C.F.; Sun, K.L.; Wang, B. Preparation, identification, and activity evaluation of ten antioxidant peptides from protein hydrolysate of swim bladders of miiuy croaker (Miichthys miiuy). J. Funct. Foods 2018, 47, 503–511. [Google Scholar] [CrossRef]
- Wu, R.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H. Antioxidant and anti-freezing peptides from salmon collagen hydrolysate prepared by bacterial extracellular protease. Food Chem. 2018, 248, 346–352. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Zhao, G.X.; Yang, X.R.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Antioxidant peptides from the protein pydrolysate of spanish mackerel (Scomberomorous niphonius) muscle by in vitro gastrointestinal gigestion and their in vitro activities. Mar. Drugs 2019, 17, 531. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Sun, Q.; Song, G.; Huang, J. Extraction, identification and structure-activity relationship of antioxidant peptides from sesame (Sesamum indicum L.) protein hydrolysate. Food Res. Int. 2019, 116, 707–716. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products—A review. Trends Food Sci. Tech. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Chi, C.F.; Ma, J.H.; Luo, H.Y.; Xu, Y.F. Purification and characterisation of a novel antioxidant peptide derived from blue mussel (Mytilus edulis) protein hydrolysate. Food Chem. 2013, 138, 1713–1719. [Google Scholar] [CrossRef]
- Timón, M.L.; Andrés, A.I.; Otte, J.; Petrón, M.J. Antioxidant peptides (<3 kDa) identified on hard cow milk cheese with rennet from different origin. Food Res. Int. 2019, 120, 643–649. [Google Scholar]
- Wong, F.C.; Xiao, J.; Ong, M.G.L.; Pang, M.J.; Wong, S.J.; Teh, L.K.; Chai, T.T. Identification and characterization of antioxidant peptides from hydrolysate of blue-spotted stingray and their stability against thermal, pH and simulated gastrointestinal digestion treatments. Food Chem. 2019, 271, 614–622. [Google Scholar] [CrossRef]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish (Rhopilema esculentum) on mice skin photoaging induced by UV irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar] [CrossRef]
- Chen, T.; Hou, H. Protective effect of gelatin polypeptides from Pacific cod (Gadus macrocephalus) against UV irradiation-induced damages by inhibiting inflammation and improving transforming growth Factor-β/Smad signaling pathway. J. Photoch. Photobio. B 2016, 162, 633–640. [Google Scholar] [CrossRef]
- Yang, X.R.; Zhang, L.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Purification and characterization of antioxidant peptides derived from protein hydrolysate of the marine bivalve mollusk Tergillarca granosa. Mar. Drugs 2019, 17, 251. [Google Scholar] [CrossRef]
- Zheng, L.; Yu, H.; Wei, H.; Xing, Q.; Zou, Y.; Zhou, Y.; Peng, J. Antioxidative peptides of hydrolysate prepared from fish skin gelatin using ginger protease activate antioxidant response element-mediated gene transcription in IPEC-J2 cells. J. Funct. Foods 2018, 51, 104–112. [Google Scholar] [CrossRef]
- He, Y.; Pan, X.; Chi, C.F.; Sun, K.L.; Wang, B. Ten new pentapeptides from protein hydrolysate of miiuy croaker (Miichthys miiuy) muscle: Preparation, identification, and antioxidant activity evaluation. LWT 2019, 105, 1–8. [Google Scholar] [CrossRef]
- Che, R.; Sun, Y.; Sun, D.; Xu, T. Characterization of the Miiuy Croaker (Miichthys miiuy) Transcriptome and Development of Immune-Relevant Genes and Molecular Markers. PLoS ONE 2014, 9, e94046. [Google Scholar] [CrossRef]
- Zhao, Y.Q.; Zeng, L.; Yang, Z.S.; Huang, F.F.; Ding, G.F.; Wang, B. Anti-fatigue effect by peptide fraction from protein hydrolysate of croceine croaker (Pseudosciaena crocea) swim bladder through inhibiting the oxidative reactions including DNA damage. Mar. Drugs 2016, 14, 221. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, Z.; Xie, C.; Gong, W.; Hu, Z.; Peng, Y. A novel mechanism of Gamma-aminobutyric acid (GABA) protecting human umbilical vein endothelial cells (HUVECs) against H2O2-induced oxidative injury. Comp. Biochem. Phy. C 2019, 217, 68–75. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, Y.Q.; Hu, F.Y.; Wang, B. Preparation and identification of antioxidant peptides from protein hydrolysate of skate (Raja porosa) cartilage. J. Funct. Foods 2016, 25, 220–230. [Google Scholar] [CrossRef]
- Tao, J.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Bioactive peptides from cartilage protein hydrolysate of spotless smoothhound and their antioxidant activity in vitro. Mar. Drugs 2018, 16, 100. [Google Scholar] [CrossRef]
- Yang, B.; Oo, T.N.; Rizzo, V. Lipid rafts mediate H2O2 prosurvival effects in cultured endothelial cells. FASEB J. 2006, 20, 1501–1503. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.X.; Zhao, Y.Q.; Qiu, Y.T.; Chi, C.F.; Wang, B. Identification and active evaluation of antioxidant peptides from protein hydrolysates of skipjack tuna (Katsuwonus pelamis) head. Antioxidants 2019, 8, 318. [Google Scholar] [CrossRef] [PubMed]
- León-López, A.; Fuentes-Jiménez, L.; Hernández-Fuentes, A.D.; Campos-Montiel, R.G.; Aguirre-Álvarez, G. Hydrolysed collagen from sheepskins as a source of functional peptides with antioxidant activity. Int. J. Mol. Sci. 2019, 20, 3931. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Zhang, Z.; Lin, S. Effects of pulsed electric field on intracellular antioxidant activity and antioxidant enzyme regulating capacities of pine nut (Pinus koraiensis) peptide QDHCH in HepG2 cells. Food Chem. 2017, 237, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Pepe, G.; Sommella, E.; Ventre, G.; Scala, M.C.; Adesso, S.; Ostacolo, C.; Marzocco, S.; Novellino, E.; Campiglia, P. Antioxidant peptides released from gastrointestinal digestion of ″Stracchino″ soft cheese: Characterization, in vitro intestinal protection and bioavailability. J. Funct. Foods 2016, 26, 494–505. [Google Scholar] [CrossRef]
- Limmongkon, A.; Pankam, J.; Somboon, T.; Wongshaya, P.; Nopprang, P. Evaluation of the DNA damage protective activity of the germinated peanut (Arachis hypogaea) in relation to antioxidant and anti-inflammatory activity. LWT 2019, 101, 259–268. [Google Scholar] [CrossRef]
- Afanasieva, K.; Sivolob, A. Physical principles and new applications of comet assay. Biophys. Chem. 2018, 238, 1–7. [Google Scholar] [CrossRef]
- Collins, A.R. The comet assay for DNA damage and repair: Principles, applications, and limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Xue, Z.; Liu, Z.; Wu, M.; Zhuang, S.; Yu, W. Effect of rapeseed peptide on DNA damage and apoptosis in Hela cells. Exp. Toxicol. Pathol. 2010, 62, 519–523. [Google Scholar] [CrossRef]
- Malik, Q.; Herbert, K.E. Oxidative and non-oxidative DNA damage and cardiovascular disease. Free Radical Res. 2012, 46, 554–564. [Google Scholar] [CrossRef]
- Zaremba, T.; Oliński, R. Oxidative DNA damage—Analysis and clinical significance. Postepy Biochem. 2010, 56, 124–138. [Google Scholar] [PubMed]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhakar, S.; Naz, R.A. Preparation of potent antioxidant peptide from edible part of shortclub cuttlefish against radical mediated lipid and DNA damage. LWT 2015, 64, 593–601. [Google Scholar] [CrossRef]
- Chen, M.F.; Gong, F.; Zhang, Y.Y.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.J. Preventive effect of YGDEY from Tilapia fish skin gelatin hydrolysates against alcohol-induced damage in HepG2 cells through ROS-mediated signaling pathways. Nutrients 2019, 11, E392. [Google Scholar] [CrossRef]
- Li, Z.; Wang, B.; Chi, C.; Gong, Y.; Luo, H.; Ding, G. Influence of average molecular weight on antioxidant and functional properties of cartilage collagen hydrolysates from Sphyrna lewini, Dasyatis akjei and Raja porosa. Food Res. Int. 2013, 51, 283–293. [Google Scholar] [CrossRef]
- Chi, C.; Wang, B.; Li, Z.; Hu, F.; Luo, H. Influence of different hydrolysis conditions on physicochemical, antioxidant and functional properties of collagen hydrolysates from cartilages of Sphyrna lewini, Dasyatis akjei and Raja porosa. J. Aquat. Food Prod. Technol. 2016, 25, 616–632. [Google Scholar] [CrossRef]
- Qiu, Y.T.; Wang, Y.M.; Yang, X.R.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Gelatin and antioxidant peptides from gelatin hydrolysate of skipjack tuna (Katsuwonus pelamis) scales: Preparation, identification and activity evaluation. Mar. Drugs 2019, 17, 565. [Google Scholar] [CrossRef]
- Zhang, J.B.; Zhao, Y.Q.; Wang, Y.M.; Chi, C.F.; Wang, B. Eight collagen peptides from hydrolysate fraction of spanish mackerel skins: Isolation, identification, and in vitro antioxidant activity evaluation. Mar. Drugs 2019, 17, 224. [Google Scholar] [CrossRef]
- Sheih, I.C.; Wu, T.K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 100, 3419–3425. [Google Scholar] [CrossRef]
- Chen, C.; Chi, Y.J.; Zhao, M.Y.; Lv, L. Purification and identification of antioxidant peptides from egg white protein hydrolysates. Amino Acids 2012, 43, 457–466. [Google Scholar] [CrossRef]
- Gimenez, B.; Aleman, A.; Montero, P.; Gomez-Guillen, M.C. Antioxidant and functional properties of gelatin hydrolysates obtained from skin of sole and squid. Food Chem. 2009, 114, 976–983. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, Y.; Chi, C.; Hu, F.; Deng, S.; Ma, J. Isolation and characterization of collagen and antioxidant collagen peptides from scales of croceine croaker (Pseudosciaena crocea). Mar. Drugs 2013, 11, 4641–4661. [Google Scholar] [CrossRef] [PubMed]
- Memarpoor-Yazdi, M.; Asoodeh, A.; Chamani, J. A novel antioxidant and antimicrobial peptide from hen egg white lysozyme hydrolysates. J. Funct. Foods 2012, 4, 278–286. [Google Scholar] [CrossRef]
- Lim, S.; Choi, A.H.; Kwon, M.; Joung, E.J.; Shin, T.; Lee, S.G.; Kim, N.G.; Kim, H.R. Evaluation of antioxidant activities of various solvent extract from Sargassum serratifolium and its major antioxidant components. Food Chem. 2019, 278, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Ghassemi-Barghi, N.; Varshosaz, J.; Etebari, M.; Dehkordi, A.J. Role of recombinant human erythropoietin loading chitosan-tripolyphosphate nanoparticles in busulfan-induced genotoxicity: Analysis of DNA fragmentation via comet assay in cultured HepG2 cells. Toxicol. in Vitro 2016, 36, 46–52. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Control | Model | NAc | FPYLRH (S8, µM) | ||
|---|---|---|---|---|---|---|
| 200 | 100 | 50 | ||||
| Cell Number (n) | 104 | 117 | 121 | 111 | 107 | 118 |
| HDNA (%) | 91.9 ± 4.2 | 18.2 ± 3.2 ### | 80.7 ± 3.8 *** | 78.4 ± 7.9 *** | 53.2 ± 6.2 *** | 37.5 ± 5.1 *** |
| TDNA (%) | 8.1 ± 4.2 | 81.8 ± 3.2 ### | 19.3 ± 4.8 * | 21.6 ± 7.9 *** | 46.8 ± 6.2 *** | 62.5 ± 5.1 *** |
| CL (pix) | 68.3 ± 3.3 | 77.3 ± 13.5 # | 66.2 ± 3.3 *** | 58.4 ± 5.7 *** | 60.0 ± 7.3 *** | 76.1 ± 10.6 * |
| TL (pix) | 7.3 ± 0.33 | 59.3 ± 6.6 ### | 27.2 ± 1.6 *** | 23.4 ± 2.4 *** | 25.0 ± 3.3 *** | 45.1 ± 6.5 *** |
| TM | 0.6 ± 0.03 | 46.4 ± 8.1 ### | 5.2 ± 0.24 *** | 5.0 ± 0.5 *** | 11.7 ± 1.4 *** | 28.1 ± 3.9 *** |
| OTM | 2.3 ± 0.1 | 24.5 ± 4.3 ### | 4.7 ± 0.2 *** | 4.7 ± 0.5 *** | 10.4 ± 1.2 *** | 17.2 ± 2.4 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, S.-Y.; Wang, Y.-M.; Zhao, Y.-Q.; Chi, C.-F.; Wang, B. Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H2O2-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury. Int. J. Mol. Sci. 2019, 20, 5425. https://doi.org/10.3390/ijms20215425
Cai S-Y, Wang Y-M, Zhao Y-Q, Chi C-F, Wang B. Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H2O2-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury. International Journal of Molecular Sciences. 2019; 20(21):5425. https://doi.org/10.3390/ijms20215425
Chicago/Turabian StyleCai, Shi-Ying, Yu-Mei Wang, Yu-Qin Zhao, Chang-Feng Chi, and Bin Wang. 2019. "Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H2O2-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury" International Journal of Molecular Sciences 20, no. 21: 5425. https://doi.org/10.3390/ijms20215425
APA StyleCai, S. -Y., Wang, Y. -M., Zhao, Y. -Q., Chi, C. -F., & Wang, B. (2019). Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H2O2-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury. International Journal of Molecular Sciences, 20(21), 5425. https://doi.org/10.3390/ijms20215425