Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells


 and
and
Abstract
:1. Introduction
2. Results
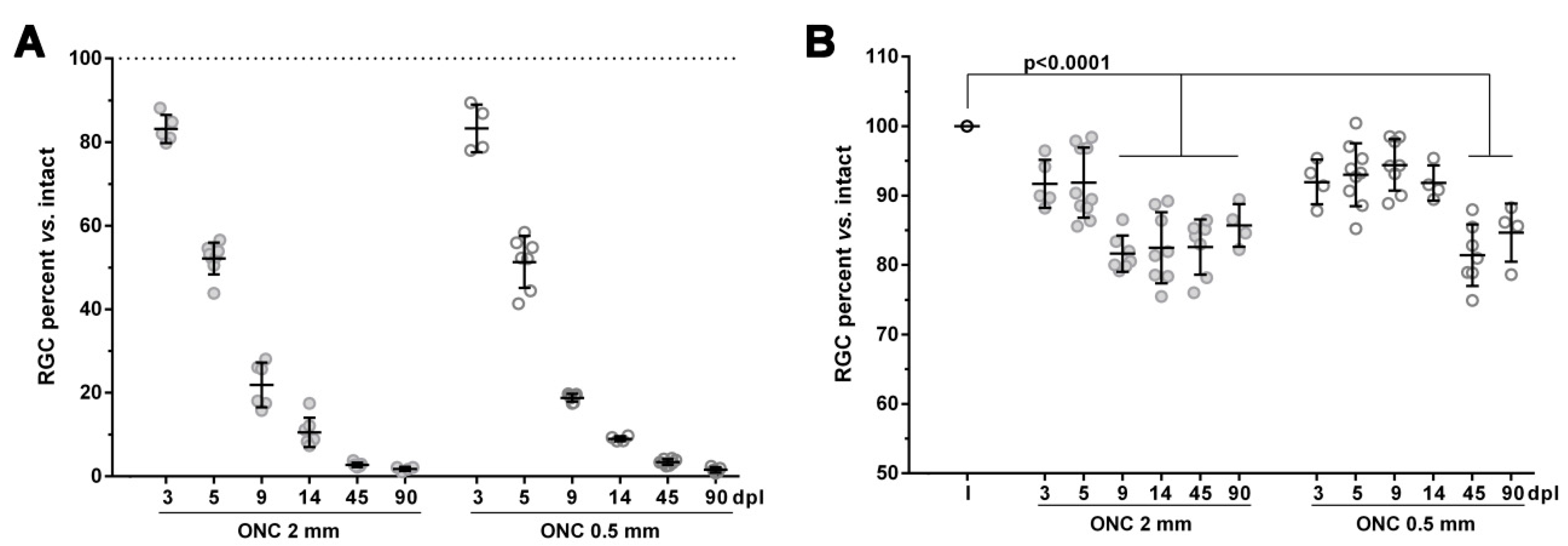
2.1. The Course of RGC Death in the Injured Retinas is not Affected by the Intraorbital Distance
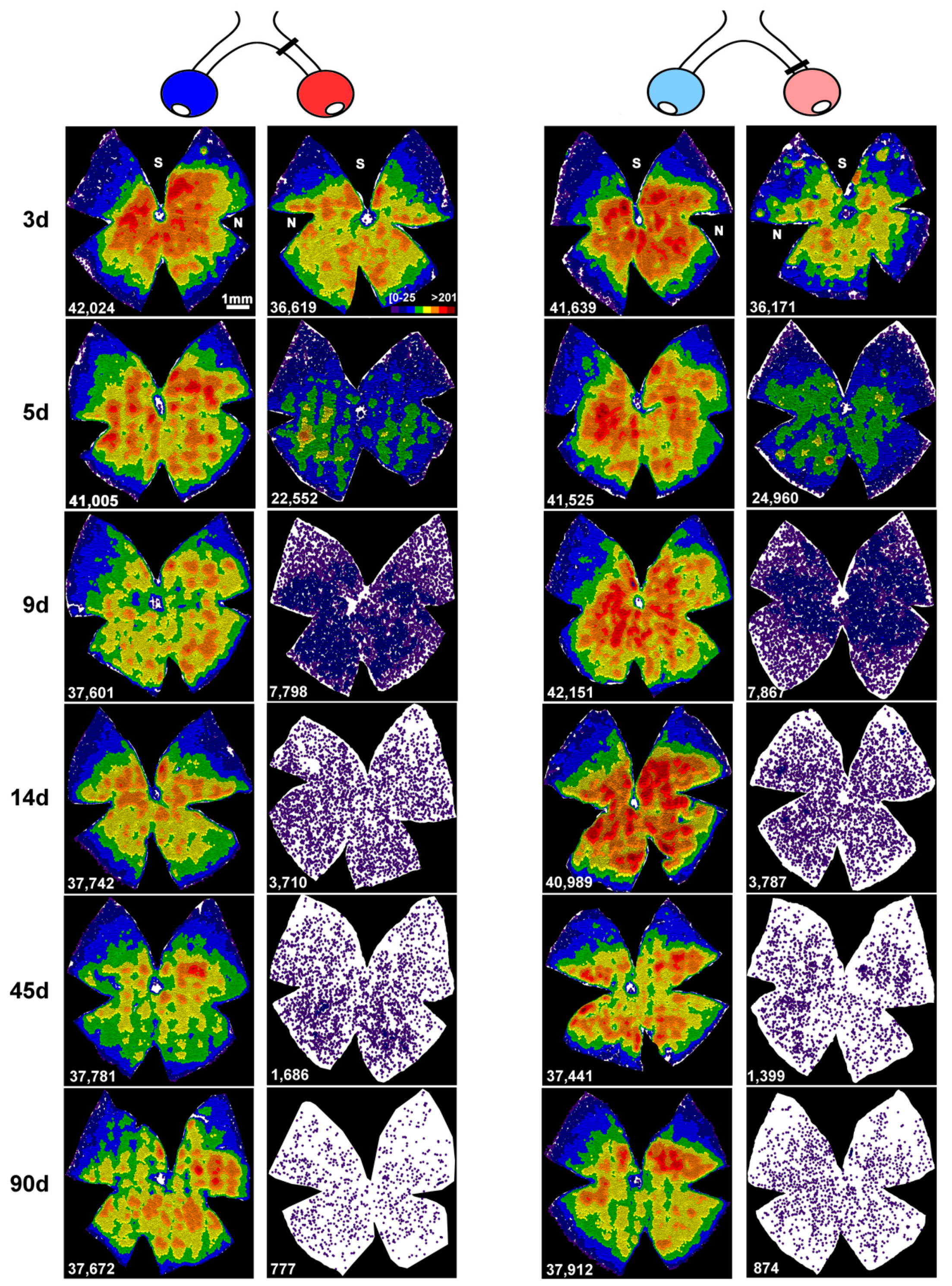
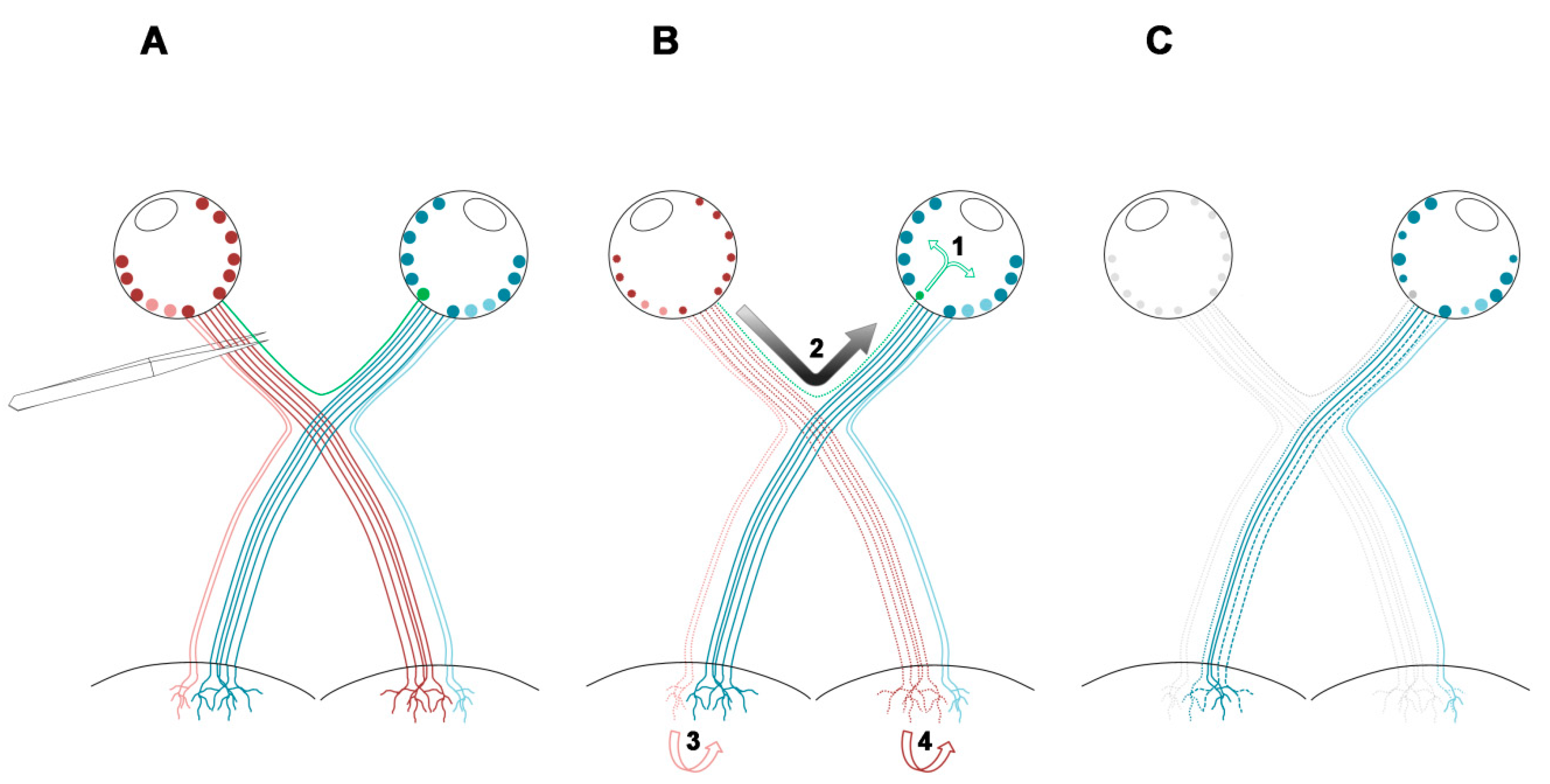
2.2. An Unilateral Axotomy Causes RGC Loss in the Contralateral Un-Injured Retina Whose Course Is Influenced by the Lesion Distance
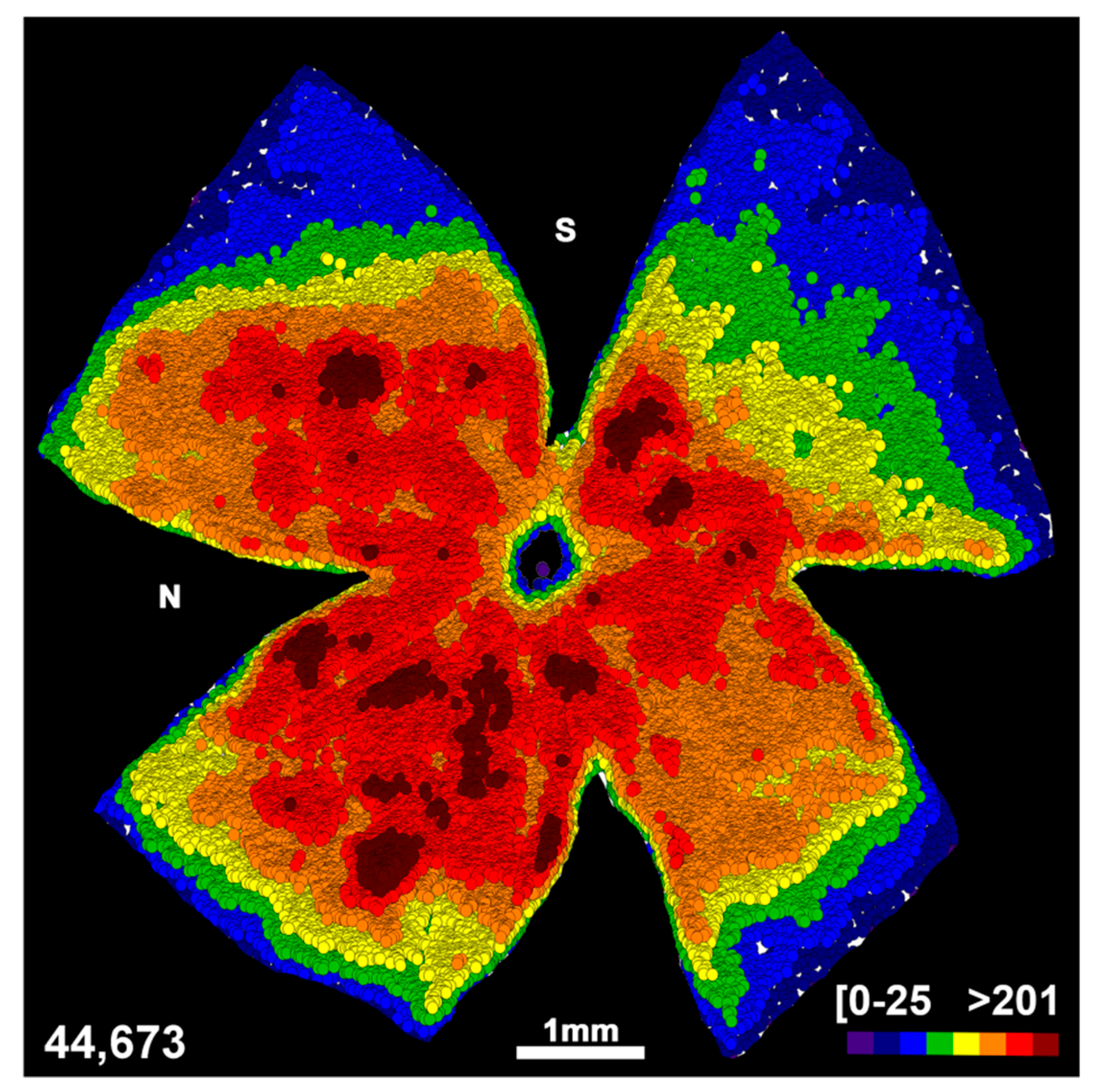
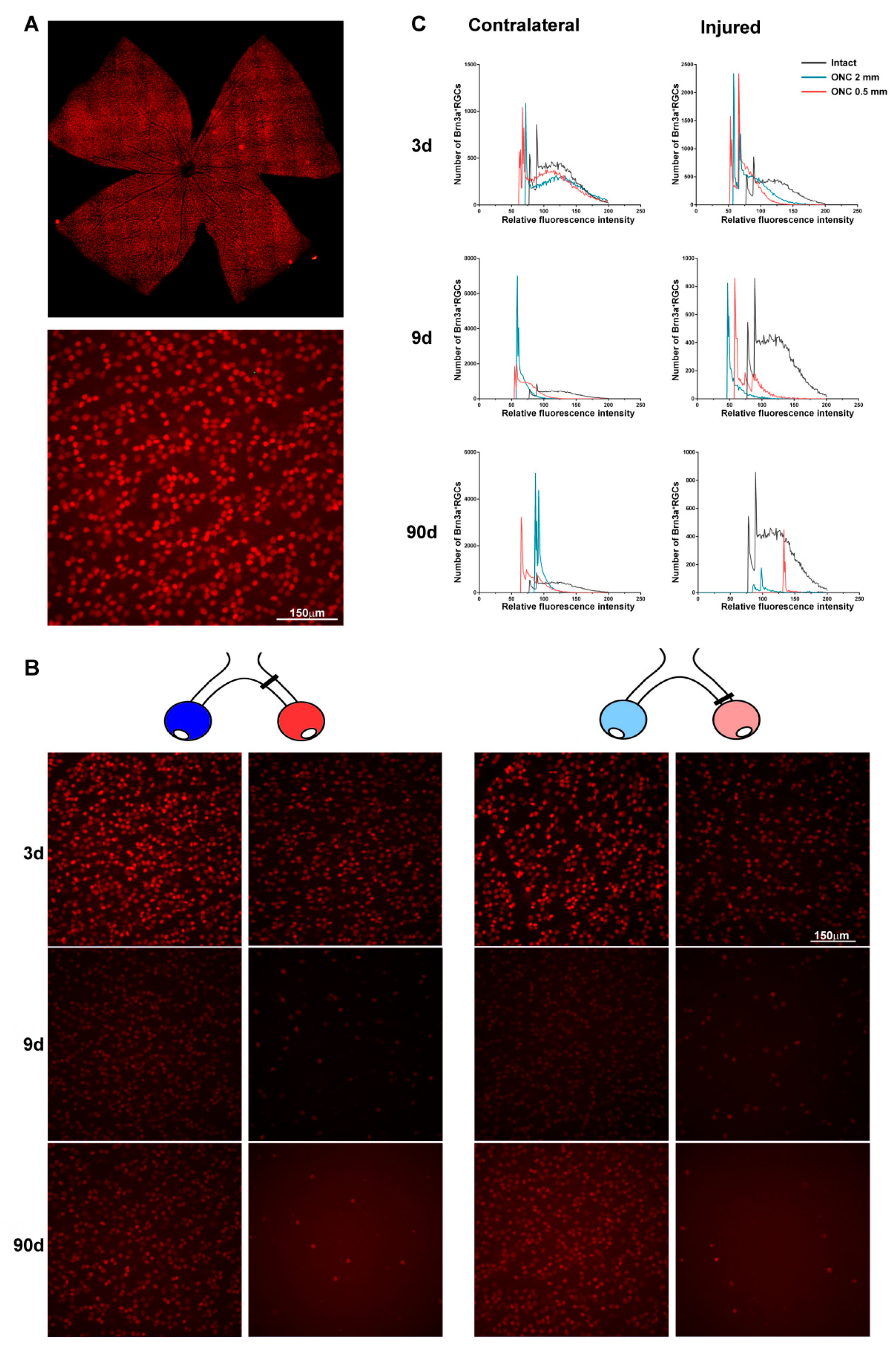
2.3. Transient Down-Regulation of Brn3a Expression in the Surviving RGCs
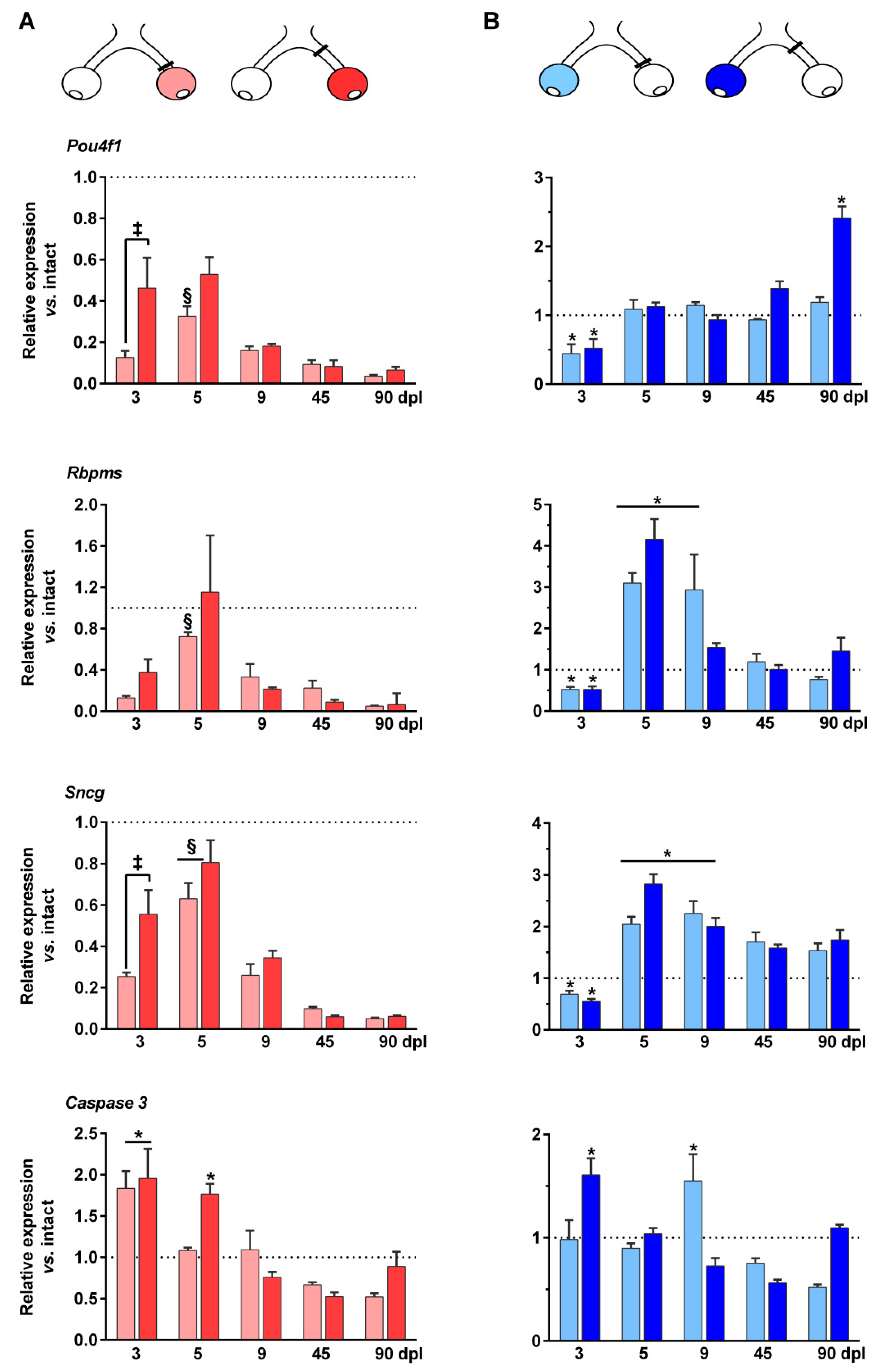
2.4. mRNA Regulation of RGC Markers in the Injured and Contralateral Retinas
2.5. Up-Regulation of Caspase 3 mRNA Expression in the Injured and Contralateral Retinas
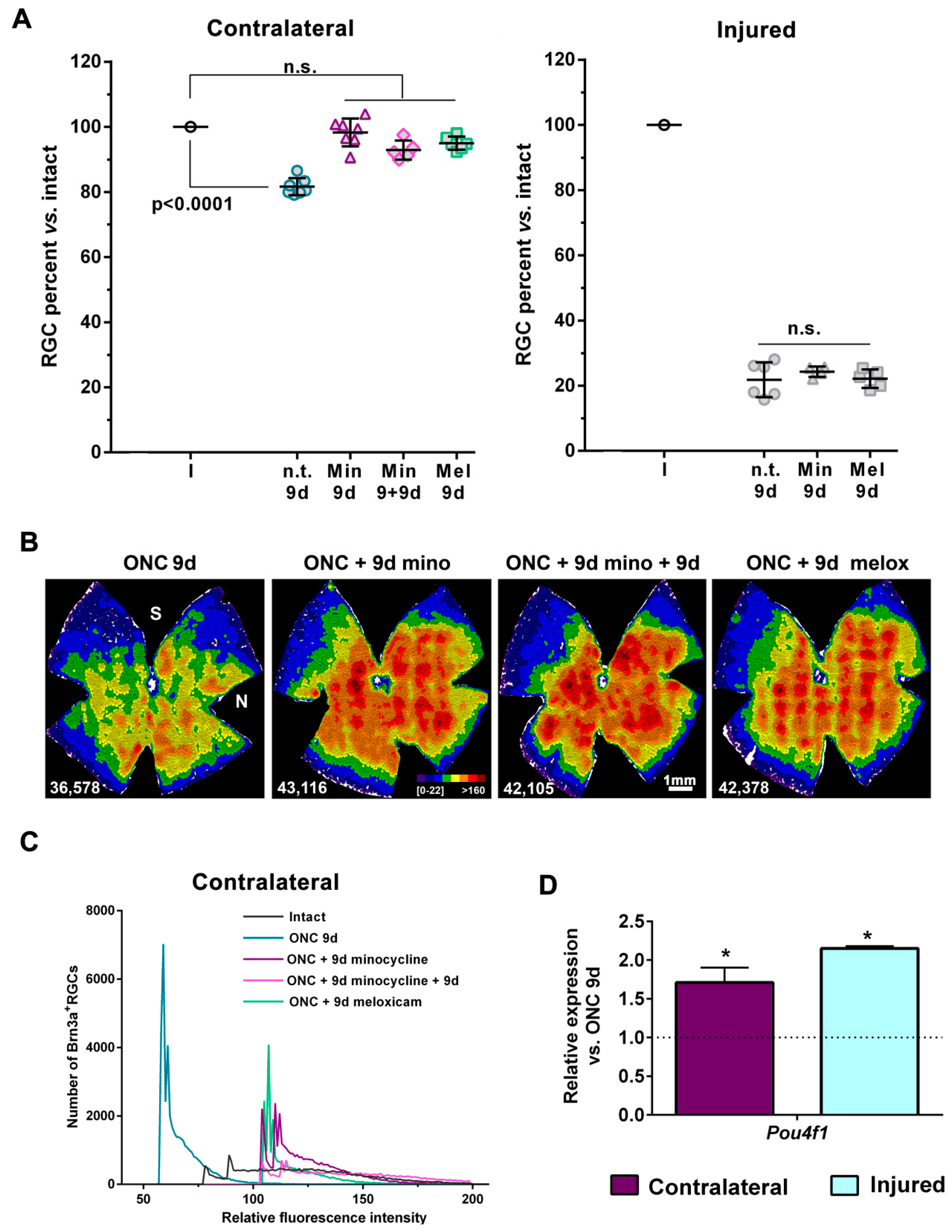
2.6. Treatment with Minocycline or Meloxicam Rescues RGCs in the Contralateral Retinas
2.7. Minocycline Treatment Reduces Microglial Activation and Increases the Expression of M2 Microglial Transcripts and of Anti-Inflammatory Mediators
3. Discussion
4. Materials and Methods
4.1. Animal Handling
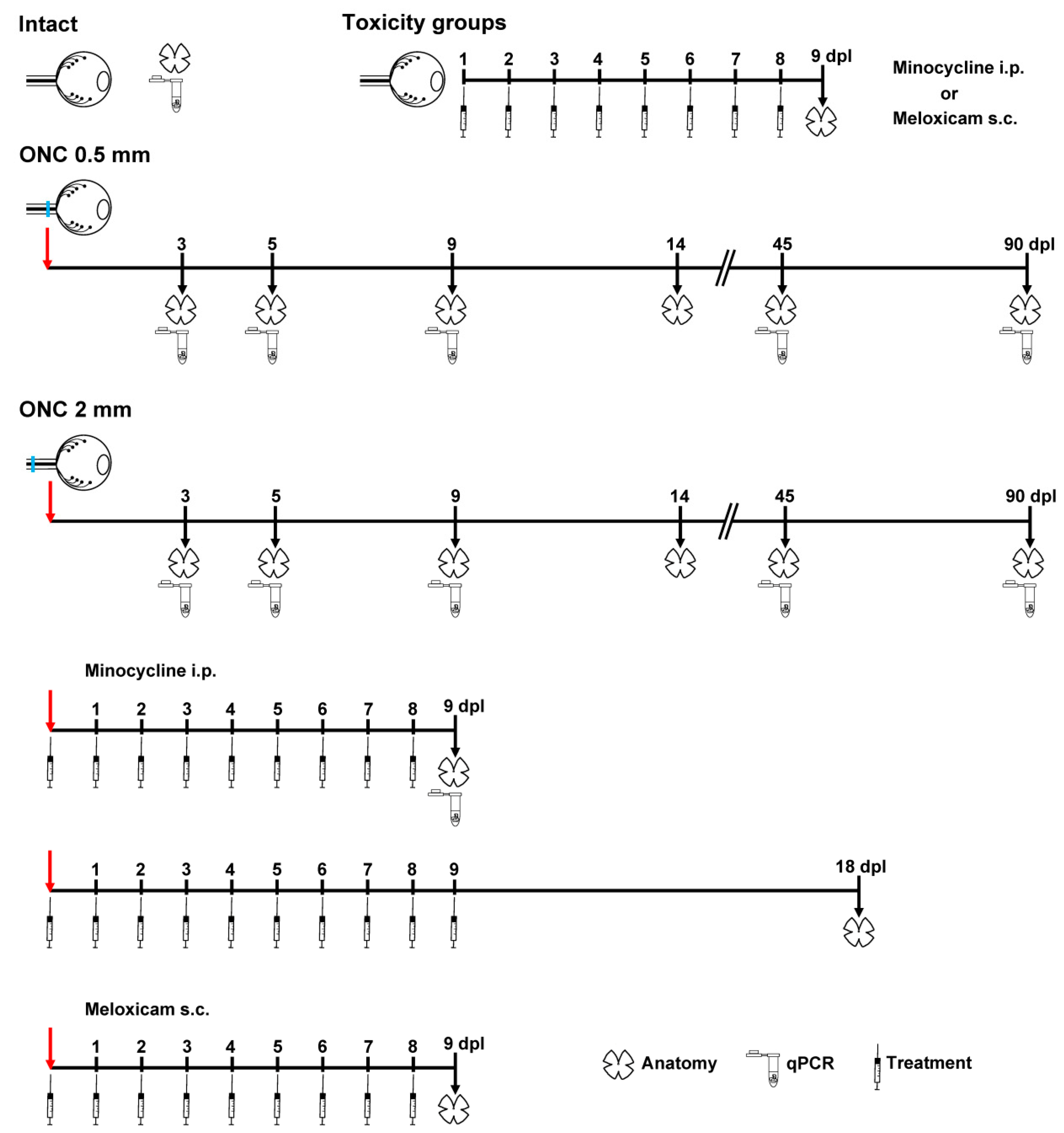
4.2. Animal Groups and Experimental Design
4.3. Optic Nerve Crush
4.4. Minocycline and Meloxicam Treatment
4.5. Immunodetection
4.6. Image Acquisition and Analysis
4.7. Quantitative Real Time PCR
4.8. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| NSAID | Non-steroidal anti-inflammatory drug |
| ONC | Optic nerve crush |
| RGC | Retinal ganglion cell |
References
- Nadal-Nicolas, F.M.; Sobrado-Calvo, P.; Jimenez-Lopez, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Long-Term Effect of Optic Nerve Axotomy on the Retinal Ganglion Cell Layer. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6095–6112. [Google Scholar] [CrossRef]
- Janssen, K.T.; Mac Nair, C.E.; Dietz, J.A.; Schlamp, C.L.; Nickells, R.W. Nuclear atrophy of retinal ganglion cells precedes the bax-dependent stage of apoptosis. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1805–1815. [Google Scholar] [CrossRef]
- Galindo-Romero, C.; Aviles-Trigueros, M.; Jimenez-Lopez, M.; Valiente-Soriano, F.J.; Salinas-Navarro, M.; Nadal-Nicolas, F.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Axotomy-induced retinal ganglion cell death in adult mice: Quantitative and topographic time course analyses. Exp. Eye Res. 2011, 92, 377–387. [Google Scholar] [CrossRef]
- Galindo-Romero, C.; Valiente-Soriano, F.J.; Jimenez-Lopez, M.; Garcia-Ayuso, D.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Effect of brain-derived neurotrophic factor on mouse axotomized retinal ganglion cells and phagocytic microglia. Investig. Ophthalmol. Vis. Sci. 2013, 54, 974–985. [Google Scholar] [CrossRef]
- Nadal-Nicolas, F.M.; Jimenez-Lopez, M.; Sobrado-Calvo, P.; Nieto-Lopez, L.; Canovas-Martinez, I.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo, M. Brn3a as a marker of retinal ganglion cells: Qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3860–3868. [Google Scholar] [CrossRef]
- Sanchez-Migallon, M.C.; Valiente-Soriano, F.J.; Nadal-Nicolas, F.M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Apoptotic Retinal Ganglion Cell Death After Optic Nerve Transection or Crush in Mice: Delayed RGC Loss With BDNF or a Caspase 3 Inhibitor. Investig. Ophthalmol. Vis. Sci. 2016, 57, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Berkelaar, M.; Clarke, D.B.; Wang, Y.C.; Bray, G.M.; Aguayo, A.J. Axotomy results in delayed death and apoptosis of retinal ganglion cells in adult rats. J. Neurosci. 1994, 14, 4368–4374. [Google Scholar] [CrossRef] [PubMed]
- Magharious, M.M.; D’Onofrio, P.M.; Koeberle, P.D. Optic nerve transection: A model of adult neuron apoptosis in the central nervous system. J. Vis. Exp. 2011, 51, e2241. [Google Scholar] [CrossRef]
- Villegas-Perez, M.P.; Vidal-Sanz, M.; Rasminsky, M.; Bray, G.M.; Aguayo, A.J. Rapid and protracted phases of retinal ganglion cell loss follow axotomy in the optic nerve of adult rats. J. Neurobiol. 1993, 24, 23–36. [Google Scholar] [CrossRef]
- Vidal-Sanz, M.; Villegas-Perez, M.P.; Bray, G.M.; Aguayo, A.J. Persistent retrograde labeling of adult rat retinal ganglion cells with the carbocyanine dye diI. Exp. Neurol. 1988, 102, 92–101. [Google Scholar] [CrossRef]
- Villegas-Perez, M.P.; Vidal-Sanz, M.; Bray, G.M.; Aguayo, A.J. Influences of peripheral nerve grafts on the survival and regrowth of axotomized retinal ganglion cells in adult rats. J. Neurosci. 1988, 8, 265–280. [Google Scholar] [CrossRef]
- Vidal-Sanz, M.; Galindo-Romero, C.; Valiente-Soriano, F.J.; Nadal-Nicolas, F.M.; Ortin-Martinez, A.; Rovere, G.; Salinas-Navarro, M.; Lucas-Ruiz, F.; Sanchez-Migallon, M.C.; Sobrado-Calvo, P.; et al. Shared and Differential Retinal Responses against Optic Nerve Injury and Ocular Hypertension. Front. Neurosci. 2017, 11, 235. [Google Scholar] [CrossRef]
- Bodeutsch, N.; Siebert, H.; Dermon, C.; Thanos, S. Unilateral injury to the adult rat optic nerve causes multiple cellular responses in the contralateral site. J. Neurobiol. 1999, 38, 116–128. [Google Scholar] [CrossRef]
- Macharadze, T.; Goldschmidt, J.; Marunde, M.; Wanger, T.; Scheich, H.; Zuschratter, W.; Gundelfinger, E.D.; Kreutz, M.R. Interretinal transduction of injury signals after unilateral optic nerve crush. Neuroreport 2009, 20, 301–305. [Google Scholar] [CrossRef]
- Panagis, L.; Thanos, S.; Fischer, D.; Dermon, C.R. Unilateral optic nerve crush induces bilateral retinal glial cell proliferation. Eur. J. Neurosci. 2005, 21, 2305–2309. [Google Scholar] [CrossRef]
- Cen, L.P.; Han, M.; Zhou, L.; Tan, L.; Liang, J.J.; Pang, C.P.; Zhang, M. Bilateral retinal microglial response to unilateral optic nerve transection in rats. Neuroscience 2015, 311, 56–66. [Google Scholar] [CrossRef]
- Lovasik, J.V.; Kergoat, H.; Gagnon, M. Experimentally reduced perfusion of one eye impairs retinal function in both eyes. Optom. Vis. Sci. 2005, 82, 850–857. [Google Scholar] [CrossRef]
- Sanchez-Migallon, M.C.; Valiente-Soriano, F.J.; Salinas-Navarro, M.; Nadal-Nicolas, F.M.; Jimenez-Lopez, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Nerve fibre layer degeneration and retinal ganglion cell loss long term after optic nerve crush or transection in adult mice. Exp. Eye Res. 2018, 170, 40–50. [Google Scholar] [CrossRef]
- Lam, W.L.; Chow, P.H.; Chik, K.P.; Yew, D.T. Bilateral biochemical and biophysical retinal changes after unilateral ocular trauma in the mouse. Neurosci. Lett. 1996, 218, 1–4. [Google Scholar] [CrossRef]
- Kanamori, A.; Nakamura, M.; Nakanishi, Y.; Yamada, Y.; Negi, A. Long-term glial reactivity in rat retinas ipsilateral and contralateral to experimental glaucoma. Exp. Eye Res. 2005, 81, 48–56. [Google Scholar] [CrossRef]
- Lonngren, U.; Napankangas, U.; Lafuente, M.; Mayor, S.; Lindqvist, N.; Vidal-Sanz, M.; Hallbook, F. The growth factor response in ischemic rat retina and superior colliculus after brimonidine pre-treatment. Brain Res. Bull. 2006, 71, 208–218. [Google Scholar] [CrossRef]
- Ramirez, A.I.; Salazar, J.J.; de Hoz, R.; Rojas, B.; Gallego, B.I.; Salobrar-Garcia, E.; Valiente-Soriano, F.J.; Trivino, A.; Ramirez, J.M. Macro- and microglial responses in the fellow eyes contralateral to glaucomatous eyes. Prog. Brain Res. 2015, 220, 155–172. [Google Scholar]
- Kim, H.S.; Suh, Y.H. Minocycline and neurodegenerative diseases. Behav. Brain Res. 2009, 196, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Imagama, S.; Ohgomori, T.; Hirano, K.; Uchimura, K.; Sakamoto, K.; Hirakawa, A.; Takeuchi, H.; Suzumura, A.; Ishiguro, N.; et al. Minocycline selectively inhibits M1 polarization of microglia. Cell Death Dis. 2013, 4, e525. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Mesa, N.; Zarzuelo, A.; Galvez, J. What is behind the non-antibiotic properties of minocycline? Pharmacol. Res. 2013, 67, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Abcouwer, S.F.; Lin, C.M.; Shanmugam, S.; Muthusamy, A.; Barber, A.J.; Antonetti, D.A. Minocycline prevents retinal inflammation and vascular permeability following ischemia-reperfusion injury. J. Neuroinflamm. 2013, 10, 149. [Google Scholar] [CrossRef]
- Hakan, T.; Toklu, H.Z.; Biber, N.; Celik, H.; Erzik, C.; Ogunc, A.V.; Cetinel, S.; Sener, G. Meloxicam exerts neuroprotection on spinal cord trauma in rats. Int. J. Neurosci. 2011, 121, 142–148. [Google Scholar] [CrossRef]
- Sui, Y.; Stanic, D.; Tomas, D.; Jarrott, B.; Horne, M.K. Meloxicam reduces lipopolysaccharide-induced degeneration of dopaminergic neurons in the rat substantia nigra pars compacta. Neurosci. Lett. 2009, 460, 121–125. [Google Scholar] [CrossRef]
- Nadal-Nicolas, F.M.; Jimenez-Lopez, M.; Salinas-Navarro, M.; Sobrado-Calvo, P.; Alburquerque-Bejar, J.J.; Vidal-Sanz, M.; Agudo-Barriuso, M. Whole number, distribution and co-expression of brn3 transcription factors in retinal ganglion cells of adult albino and pigmented rats. PLoS ONE 2012, 7, e49830. [Google Scholar] [CrossRef]
- Rodriguez, A.R.; de Sevilla Muller, L.P.; Brecha, N.C. The RNA binding protein RBPMS is a selective marker of ganglion cells in the mammalian retina. J. Comp. Neurol. 2014, 522, 1411–1443. [Google Scholar] [CrossRef]
- Surgucheva, I.; Weisman, A.D.; Goldberg, J.L.; Shnyra, A.; Surguchov, A. Gamma-synuclein as a marker of retinal ganglion cells. Mol. Vis. 2008, 14, 1540–1548. [Google Scholar] [PubMed]
- Gallego, B.I.; Salazar, J.J.; de Hoz, R.; Rojas, B.; Ramirez, A.I.; Salinas-Navarro, M.; Ortin-Martinez, A.; Valiente-Soriano, F.J.; Aviles-Trigueros, M.; Villegas-Perez, M.P.; et al. IOP induces upregulation of GFAP and MHC-II and microglia reactivity in mice retina contralateral to experimental glaucoma. J. Neuroinflamm. 2012, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilla, A.M.; Diekmann, H.; Fischer, D. Microglia Are Irrelevant for Neuronal Degeneration and Axon Regeneration after Acute Injury. J. Neurosci. 2017, 37, 6113–6124. [Google Scholar] [CrossRef] [PubMed]
- Karlstetter, M.; Nothdurfter, C.; Aslanidis, A.; Moeller, K.; Horn, F.; Scholz, R.; Neumann, H.; Weber, B.H.; Rupprecht, R.; Langmann, T. Translocator protein (18 kDa) (TSPO) is expressed in reactive retinal microglia and modulates microglial inflammation and phagocytosis. J. Neuroinflamm. 2014, 11, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mages, K.; Grassmann, F.; Jagle, H.; Rupprecht, R.; Weber, B.H.F.; Hauck, S.M.; Grosche, A. The agonistic TSPO ligand XBD173 attenuates the glial response thereby protecting inner retinal neurons in a murine model of retinal ischemia. J. Neuroinflamm. 2019, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Appel, S.; Le, W. Minocycline inhibits microglial activation and protects nigral cells after 6-hydroxydopamine injection into mouse striatum. Brain Res. 2001, 909, 187–193. [Google Scholar] [CrossRef]
- Dommergues, M.A.; Plaisant, F.; Verney, C.; Gressens, P. Early microglial activation following neonatal excitotoxic brain damage in mice: A potential target for neuroprotection. Neuroscience 2003, 121, 619–628. [Google Scholar] [CrossRef]
- Tikka, T.; Fiebich, B.L.; Goldsteins, G.; Keinanen, R.; Koistinaho, J. Minocycline, a tetracycline derivative, is neuroprotective against excitotoxicity by inhibiting activation and proliferation of microglia. J. Neurosci. 2001, 21, 2580–2588. [Google Scholar] [CrossRef]
- Balkema, G.W.; Drager, U.C. Origins of uncrossed retinofugal projections in normal and hypopigmented mice. Vis. Neurosci. 1990, 4, 595–604. [Google Scholar] [CrossRef]
- Drager, U.C.; Olsen, J.F. Origins of crossed and uncrossed retinal projections in pigmented and albino mice. J. Comp. Neurol. 1980, 191, 383–412. [Google Scholar] [CrossRef]
- Lund, R.D. Uncrossed Visual Pathways of Hooded and Albino Rats. Science 1965, 149, 1506–1507. [Google Scholar] [CrossRef] [PubMed]
- Lund, R.D.; Land, P.W.; Boles, J. Normal and abnormal uncrossed retinotectal pathways in rats: An HRP study in adults. J. Comp. Neurol. 1980, 189, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Nicolas, F.M.; Salinas-Navarro, M.; Jimenez-Lopez, M.; Sobrado-Calvo, P.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Displaced retinal ganglion cells in albino and pigmented rats. Front. Neuroanat. 2014, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montori, S.; Dos-Anjos, S.; Martinez-Villayandre, B.; Regueiro-Purrinos, M.M.; Gonzalo-Orden, J.M.; Ruano, D.; Fernandez-Lopez, A. Age and meloxicam attenuate the ischemia/reperfusion-induced down-regulation in the NMDA receptor genes. Neurochem. Int. 2010, 56, 878–885. [Google Scholar] [CrossRef]
- Di Pierdomenico, J.; Scholz, R.; Valiente-Soriano, F.J.; Sanchez-Migallon, M.C.; Vidal-Sanz, M.; Langmann, T.; Agudo-Barriuso, M.; Garcia-Ayuso, D.; Villegas-Perez, M.P. Neuroprotective Effects of FGF2 and Minocycline in Two Animal Models of Inherited Retinal Degeneration. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4392–4403. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Nicolas, F.M.; Valiente-Soriano, F.J.; Salinas-Navarro, M.; Jimenez-Lopez, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Retino-retinal projection in juvenile and young adult rats and mice. Exp. Eye Res. 2015, 134, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Tzekov, R.; Passaglia, C.L. Retinal cross talk in the mammalian visual system. J. Neurophysiol. 2016, 115, 3018–3029. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Hollander, H. A small population of retinal ganglion cells projecting to the retina of the other eye. An experimental study in the rat and the rabbit. Exp. Brain Res. 1988, 71, 611–617. [Google Scholar] [CrossRef]
- Murcia-Belmonte, V.; Coca, Y.; Vegar, C.; Negueruela, S.; de Juan, R.C.; Valino, A.J.; Sala, S.; DaSilva, R.; Kania, A.; Borrell, V.; et al. A Retino-retinal Projection Guided by Unc5c Emerged in Species with Retinal Waves. Curr. Biol. 2019, 29, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Avellaneda-Chevrier, V.K.; Wang, X.; Hooper, M.L.; Chauhan, B.C. The retino-retinal projection: Tracing retinal ganglion cells projecting to the contralateral retina. Neurosci. Lett. 2015, 591, 105–109. [Google Scholar] [CrossRef]
- Salinas-Navarro, M.; Mayor-Torroglosa, S.; Jimenez-Lopez, M.; Aviles-Trigueros, M.; Holmes, T.M.; Lund, R.D.; Villegas-Perez, M.P.; Vidal-Sanz, M. A computerized analysis of the entire retinal ganglion cell population and its spatial distribution in adult rats. Vis. Res. 2009, 49, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salinas-Navarro, M.; Jimenez-Lopez, M.; Valiente-Soriano, F.J.; Alarcon-Martinez, L.; Aviles-Trigueros, M.; Mayor, S.; Holmes, T.; Lund, R.D.; Villegas-Perez, M.P.; Vidal-Sanz, M. Retinal ganglion cell population in adult albino and pigmented mice: A computerized analysis of the entire population and its spatial distribution. Vis. Res. 2009, 49, 637–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo-Romero, C.; Jimenez-Lopez, M.; Garcia-Ayuso, D.; Salinas-Navarro, M.; Nadal-Nicolas, F.M.; Agudo-Barriuso, M.; Villegas-Perez, M.P.; Aviles-Trigueros, M.; Vidal-Sanz, M. Number and spatial distribution of intrinsically photosensitive retinal ganglion cells in the adult albino rat. Exp. Eye Res. 2013, 108, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Post-Lesion or Treatment | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Group | Retina | Intact | 3d | 5d | 9d | 14d | 18d | 45d | 90d | |
| Intact | Mean | 44,646 | ||||||||
| SD | 695 | |||||||||
| Toxicity | Mean | 43,364 | ||||||||
| Minocycline | SD | 1926 | ||||||||
| Toxicity | Mean | 42,896 | ||||||||
| Meloxican | SD | 1593 | ||||||||
| ONC 0.5 mm | Contralateral | Mean | 41,057 | 41,525 | 42,151 | 40,989 | 36,349 *** | 37,800 *** | ||
| SD | 1436 | 2022 | 1652 | 1137 | 1971 | 1874 | ||||
| Injured | Mean | 37,178 * | 22,910 | 8444 | 4022 | 1494 | 687 | |||
| SD | 2561 | 2779 | 402 | 269 | 295 | 304 | ||||
| ONC 2 mm | Contralateral | Mean | 40,939 | 41,005 | 36,117 **** | 36,349 *** | 36,877 *** | 38,262 ** | ||
| SD | 1546 | 2257 | 1425 | 1971 | 1772 | 1366 | ||||
| Injured | Mean | 37,120 * | 23,285 | 9759 | 5128 | 1209 | 797 | |||
| SD | 1501 | 1696 | 2385 | 1891 | 236 | 224 | ||||
| ONC 2 mm + Mino | Contralateral | Mean | 43,888 ††† | |||||||
| SD | 1919 | |||||||||
| Injured | Mean | 10,854 | ||||||||
| SD | 722 | |||||||||
| ONC 2 mm + Mino + 9d | Contralateral | Mean | 41,478 † | |||||||
| SD | 1299 | |||||||||
| ONC 2 mm + Melox | Contralateral | Mean | 42,423 † | |||||||
| SD | 918 | |||||||||
| Injured | Mean | 9901 | ||||||||
| SD | 1252 | |||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucas-Ruiz, F.; Galindo-Romero, C.; Rodríguez-Ramírez, K.T.; Vidal-Sanz, M.; Agudo-Barriuso, M. Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells. Int. J. Mol. Sci. 2019, 20, 5733. https://doi.org/10.3390/ijms20225733
Lucas-Ruiz F, Galindo-Romero C, Rodríguez-Ramírez KT, Vidal-Sanz M, Agudo-Barriuso M. Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells. International Journal of Molecular Sciences. 2019; 20(22):5733. https://doi.org/10.3390/ijms20225733
Chicago/Turabian StyleLucas-Ruiz, Fernando, Caridad Galindo-Romero, Kristy T. Rodríguez-Ramírez, Manuel Vidal-Sanz, and Marta Agudo-Barriuso. 2019. "Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells" International Journal of Molecular Sciences 20, no. 22: 5733. https://doi.org/10.3390/ijms20225733
APA StyleLucas-Ruiz, F., Galindo-Romero, C., Rodríguez-Ramírez, K. T., Vidal-Sanz, M., & Agudo-Barriuso, M. (2019). Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells. International Journal of Molecular Sciences, 20(22), 5733. https://doi.org/10.3390/ijms20225733