Behavioral Impairments and Oxidative Stress in the Brain, Muscle, and Gill Caused by Chronic Exposure of C70 Nanoparticles on Adult Zebrafish

 ,
, 

Abstract
:1. Introduction
2. Results
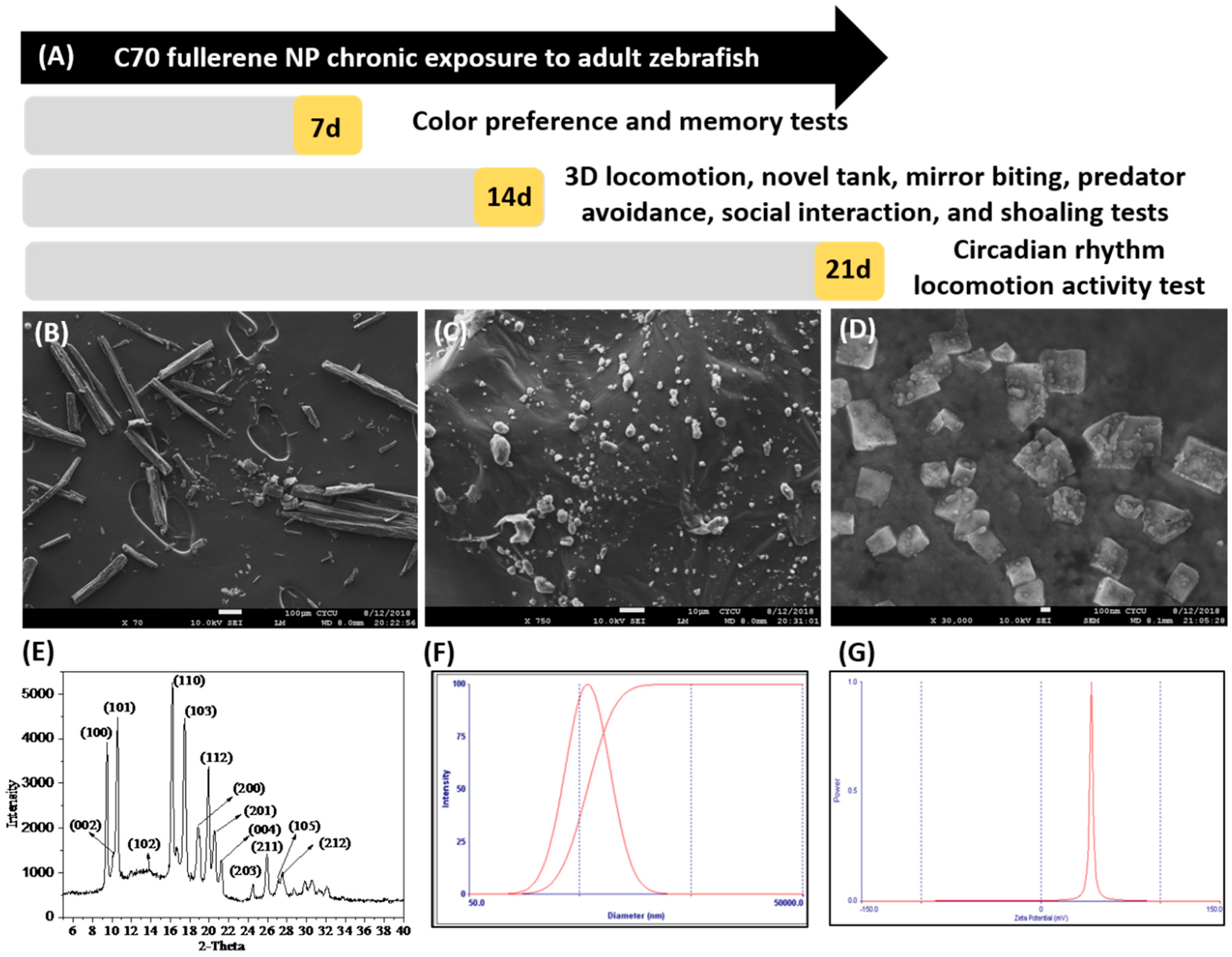
2.1. Physical Property Characterization of C70 NPs (Nanoparticles)
2.2. Low-Dose Exposure of C70 NPs Reduced Locomotion and Exploration Behaviors
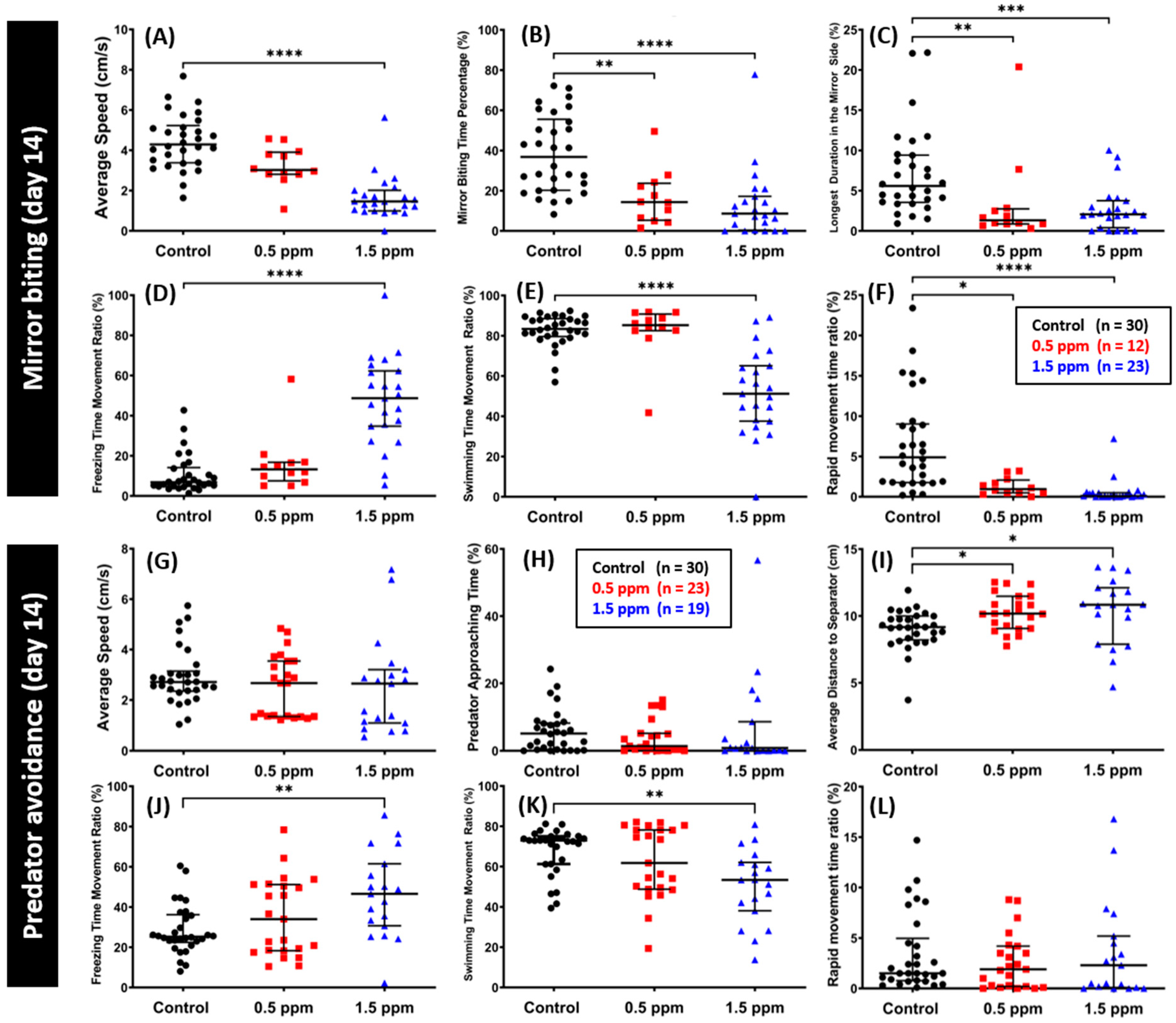
2.3. Low Doses of C70 NPs Exposure Reduced Aggression and Predator Avoidance
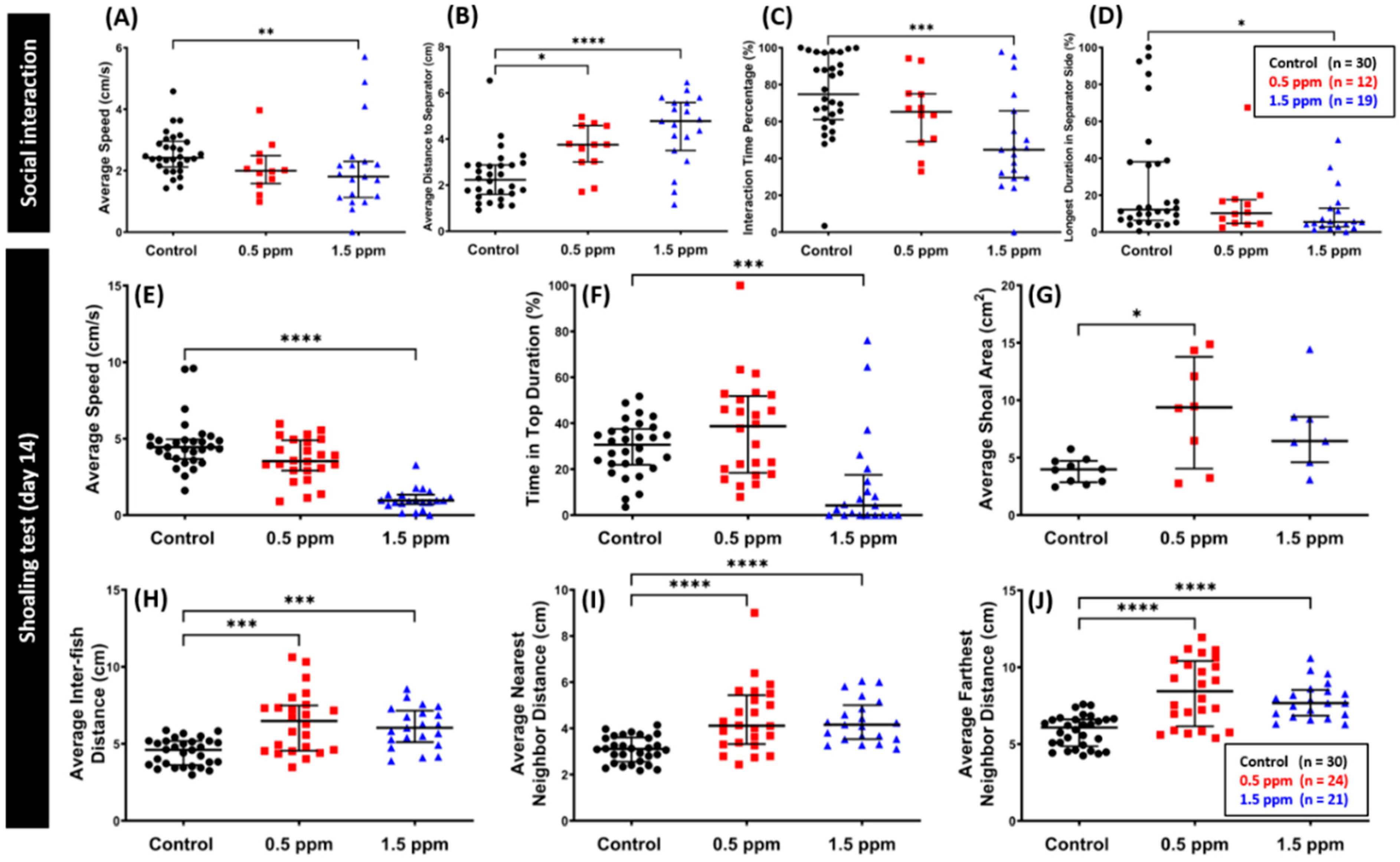
2.4. Low Doses of C70 NPs Exposure Dysregulated Social Interaction and Shoaling Behavior
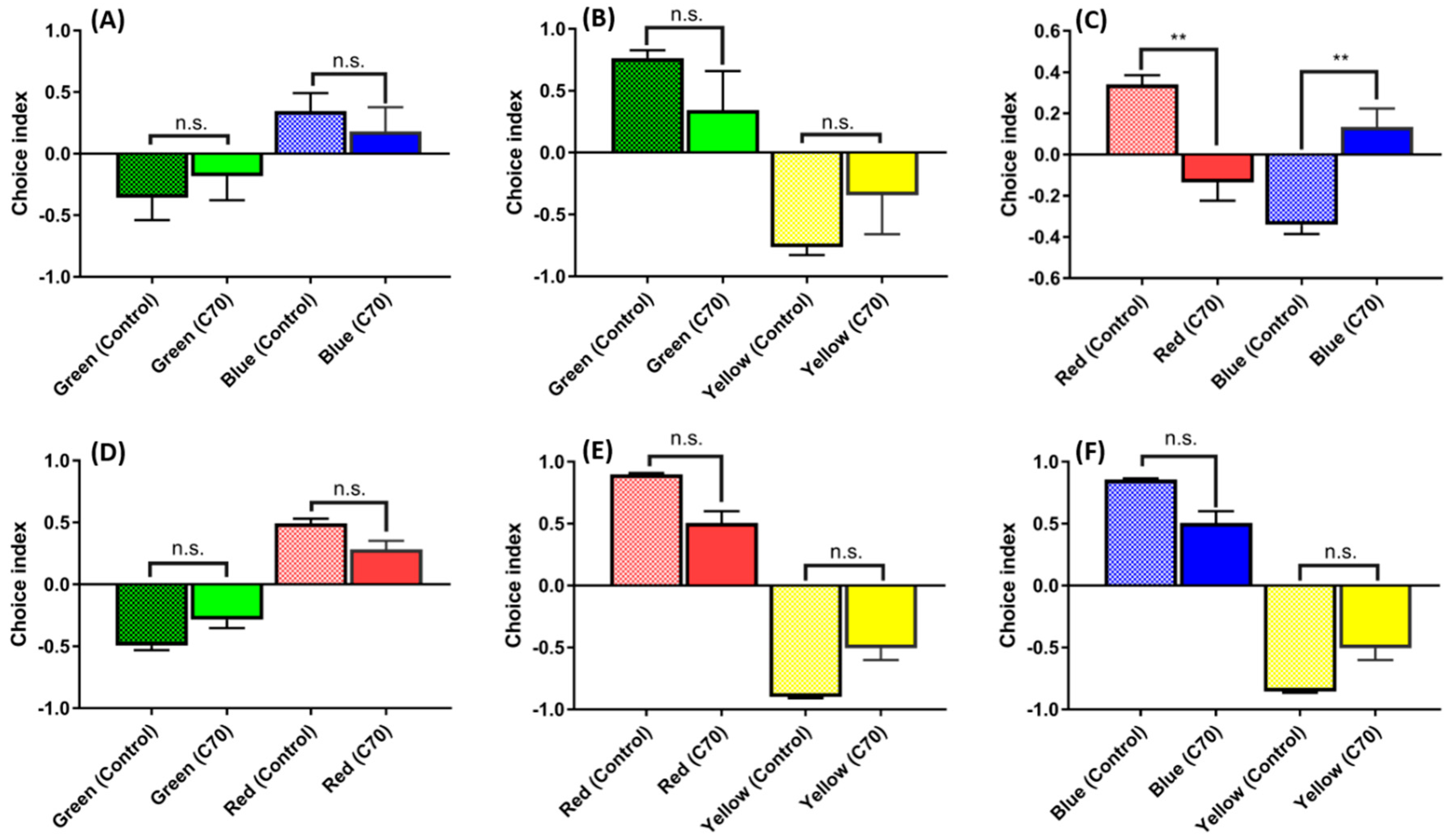
2.5. Low Doses of C70 NPs Exposure Dysregulated the Color Preference
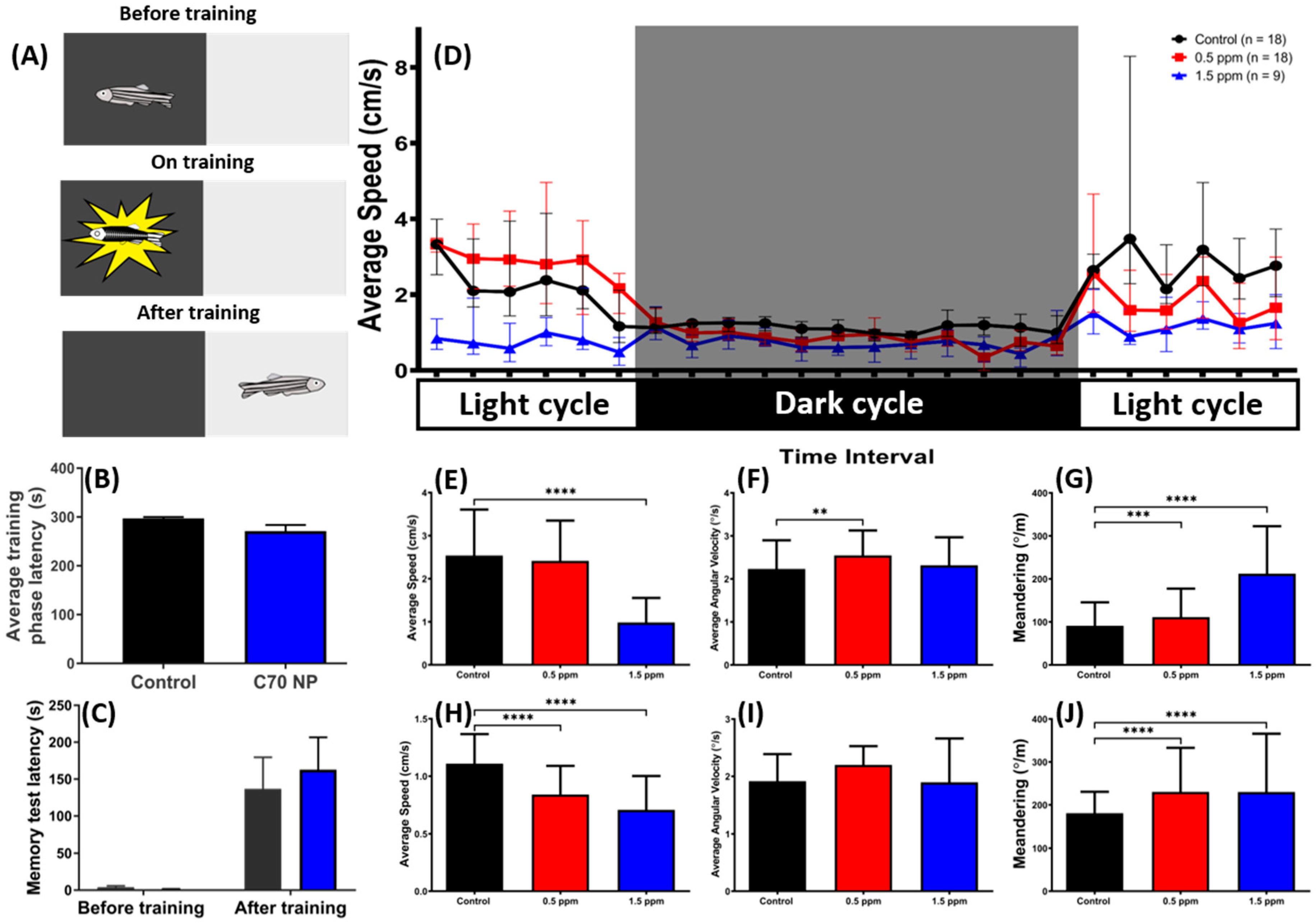
2.6. Low Doses of C70 NPs Exposure Dysregulated the Circadian Rhythm but Did Not Alter Short-Term Memory
2.7. Impact of C70 NPs Exposure on Biomarker Expression in the Muscle, Brain, and Gill
2.8. Clustering of Zebrafish Behavior Distinguished Carbon NPs (C60 and C70) Based on Their Exposure Concentration
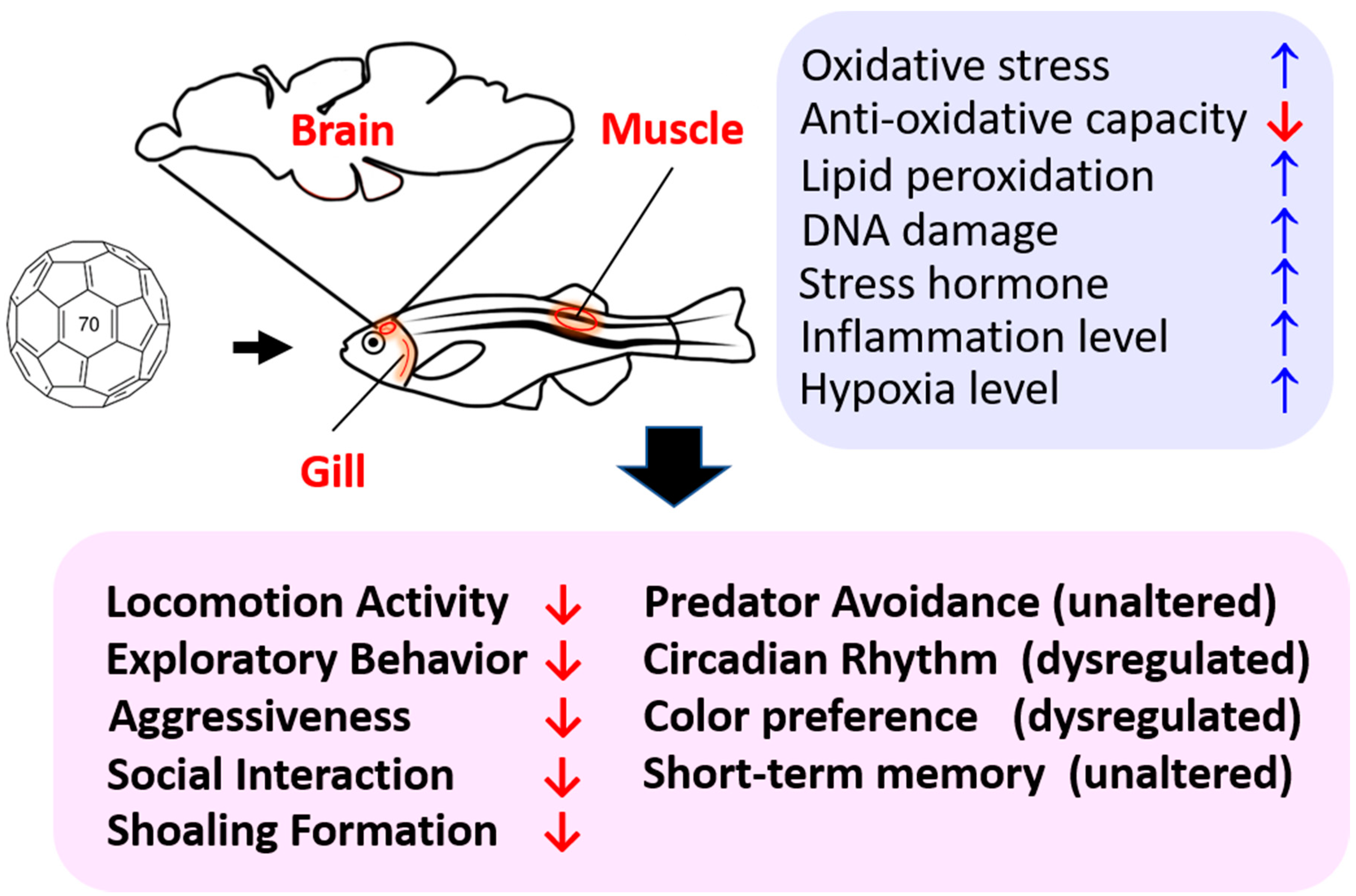
3. Discussion
4. Summary and Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Animals
5.3. C70 NPS Suspensions
5.4. Characterization of C70 NPs
5.5. Zebrafish Exposure Protocol
5.6. Behavioral Endpoints
5.6.1. 3D Locomotion Test
5.6.2. Novel Tank Test
5.6.3. Aggression Test
5.6.4. Predator-Avoidance Test
5.6.5. Social Interaction Test
5.6.7. Shoaling Test
5.6.8. Circadian Rhythm Test
5.6.9. Color Preferences Test
5.6.10. Short-Term Memory Test
5.7. Total Protein Extraction
5.8. Biochemical Parameter Assay
5.9. Statistical Analysis
5.10. PCA, Heatmap, and Clustering Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Afreen, S.; Muthoosamy, K.; Manickam, S.; Hashim, U. Functionalized fullerene (C60) as a potential nanomediator in the fabrication of highly sensitive biosensors. Biosens. Bioelectron. 2015, 63, 354–364. [Google Scholar] [CrossRef]
- Bogdanović, G.; Đorđević, A. Carbon nanomaterials: Biologically active fullerene derivatives. Srpski Arhiv Celokupno Lekarstvo 2016, 144, 222–231. [Google Scholar] [CrossRef]
- Liu, F.-f.; Zhao, J.; Wang, S.; Du, P.; Xing, B. Effects of solution chemistry on adsorption of selected pharmaceuticals and personal care products (PPCPs) by graphenes and carbon nanotubes. Environ. Sci. Technol. 2014, 48, 13197–13206. [Google Scholar] [CrossRef] [PubMed]
- Ju-Nam, Y.; Lead, J.R. Manufactured nanoparticles: An overview of their chemistry, interactions and potential environmental implications. Sci. Total Environ. 2008, 400, 396–414. [Google Scholar] [CrossRef] [PubMed]
- Usepa, U. Nanotechnology white papper; U.S. Environmental Protection Agency: Washington, DC, USA, 2007; p. 20460.
- Innocenzi, P.; Brusatin, G. Fullerene-based organi—Inorganic nanocomposites and their applications. Chem. Mater. 2001, 13, 3126–3139. [Google Scholar] [CrossRef]
- Zhu, X.; Sollogoub, M.; Zhang, Y. Biological applications of hydrophilic C60 derivatives (hC60s)−A structural perspective. Eur. J. Med. Chem. 2016, 115, 438–452. [Google Scholar] [CrossRef]
- Soleyman, R.; Hirbod, S.; Adeli, M. Advances in the biomedical application of polymer-functionalized carbon nanotubes. Biomater. Sci. 2015, 3, 695–711. [Google Scholar] [CrossRef]
- Bakry, R.; Vallant, R.M.; Najam-ul-Haq, M.; Rainer, M.; Szabo, Z.; Huck, C.W.; Bonn, G.K. Medicinal applications of fullerenes. Int. J. Nanomed. 2007, 2, 639. [Google Scholar]
- Harrison, B.S.; Atala, A. Carbon nanotube applications for tissue engineering. Biomaterials 2007, 28, 344–353. [Google Scholar] [CrossRef]
- Kamat, J.P.; Devasagayam, T.P.; Priyadarsini, K.; Mohan, H.; Mittal, J.P. Oxidative damage induced by the fullerene C60 on photosensitization in rat liver microsomes. Chem. Biol. Interact. 1998, 114, 145–159. [Google Scholar] [CrossRef]
- Wolff, D.J.; Papoiu, A.D.; Mialkowski, K.; Richardson, C.F.; Schuster, D.I.; Wilson, S.R. Inhibition of nitric oxide synthase isoforms by tris-malonyl-C60-fullerene adducts. Arch. Biochem. Biophys. 2000, 378, 216–223. [Google Scholar] [CrossRef]
- Oberdörster, E. Manufactured nanomaterials (fullerenes, C60) induce oxidative stress in the brain of juvenile largemouth bass. Environ. Health Perspect. 2004, 112, 1058–1062. [Google Scholar] [CrossRef]
- Zhu, S.; Oberdörster, E.; Haasch, M.L. Toxicity of an engineered nanoparticle (fullerene, C60) in two aquatic species, Daphnia and fathead minnow. Marine Environ. Res. 2006, 62, S5–S9. [Google Scholar] [CrossRef]
- Sarasamma, S.; Audira, G.; Juniardi, S.; Sampurna, B.; Lai, Y.-H.; Hao, E.; Chen, J.-R.; Hsiao, C.-D. Evaluation of the Effects of Carbon 60 Nanoparticle Exposure to Adult Zebrafish: A Behavioral and Biochemical Approach to Elucidate the Mechanism of Toxicity. Int. J. Mol. Sci. 2018, 19, 3853. [Google Scholar] [CrossRef] [PubMed]
- Semmler, M.; Seitz, J.; Erbe, F.; Mayer, P.; Heyder, J.; Oberdörster, G.; Kreyling, W. Long-term clearance kinetics of inhaled ultrafine insoluble iridium particles from the rat lung, including transient translocation into secondary organs. Inhalation Toxicol. 2004, 16, 453–459. [Google Scholar] [CrossRef] [PubMed]
- John, A.A.; Subramanian, A.P.; Vellayappan, M.V.; Balaji, A.; Mohandas, H.; Jaganathan, S.K. Carbon nanotubes and graphene as emerging candidates in neuroregeneration and neurodrug delivery. Int. J. Nanomed. 2015, 10, 4267. [Google Scholar]
- Yang, X.; Ebrahimi, A.; Li, J.; Cui, Q. Fullerene–biomolecule conjugates and their biomedicinal applications. Int. J. Nanomed. 2014, 9, 77. [Google Scholar] [CrossRef]
- Hurt, R.H.; Monthioux, M.; Kane, A. Toxicology of carbon nanomaterials: Status, trends, and perspectives on the special issue. Carbon 2006, 44, 1028–1033. [Google Scholar] [CrossRef]
- Monteiro-Riviere, N.; Inman, A.; Zhang, L. Limitations and relative utility of screening assays to assess engineered nanoparticle toxicity in a human cell line. Toxicol. Appl. Pharmacol. 2009, 234, 222–235. [Google Scholar] [CrossRef]
- Kroll, A.; Pillukat, M.H.; Hahn, D.; Schnekenburger, J. Current in vitro methods in nanoparticle risk assessment: Limitations and challenges. Eur. J. Pharm. 2009, 72, 370–377. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Zebrafish: A complete animal model to enumerate the nanoparticle toxicity. J. Nanobiotechnol. 2016, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, A.L. Zebrafish assays for drug toxicity screening. Exp. Opin. Drug Metabol. Toxicol. 2006, 2, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed]
- De Esch, C.; Slieker, R.; Wolterbeek, A.; Woutersen, R.; de Groot, D. Zebrafish as potential model for developmental neurotoxicity testing: A mini review. Neurotoxicology 2012, 34, 545–553. [Google Scholar] [CrossRef]
- Scholz, S.; Fischer, S.; Gündel, U.; Küster, E.; Luckenbach, T.; Voelker, D. The zebrafish embryo model in environmental risk assessment—Applications beyond acute toxicity testing. Environ. Sci. Pollut. Res. 2008, 15, 394–404. [Google Scholar] [CrossRef]
- Seda, B.C.; Ke, P.C.; Mount, A.S.; Klaine, S.J. Toxicity of aqueous C70-gallic acid suspension in Daphnia magna. Environ. Toxicol. 2012, 31, 215–220. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. In Molecular, Clinical and Environmental Toxicology; Springer: Cham, Switzerland, 2012; pp. 133–164. [Google Scholar]
- Karen, S.; Brown, T.M. Principles of Toxicology; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Herculano, A.M.; Maximino, C. Serotonergic modulation of zebrafish behavior: Towards a paradox. Progress Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 50–66. [Google Scholar] [CrossRef]
- Kokel, D.; Peterson, R.T. Using the zebrafish photomotor response for psychotropic drug screening. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 105, pp. 517–524. [Google Scholar]
- Stewart, A.M.; Gerlai, R.; Kalueff, A.V. Developing highER-throughput zebrafish screens for in-vivo CNS drug discovery. Front. Behav. Neurosci. 2015, 9, 14. [Google Scholar] [CrossRef]
- Horie, M.; Nishio, K.; Kato, H.; Shinohara, N.; Nakamura, A.; Fujita, K.; Kinugasa, S.; Endoh, S.; Yoshida, Y.; Hagihara, Y. In vitro evaluation of cellular influences induced by stable fullerene C70 medium dispersion: Induction of cellular oxidative stress. Chemosphere 2013, 93, 1182–1188. [Google Scholar] [CrossRef]
- Usenko, C.Y.; Harper, S.L.; Tanguay, R.L. In vivo evaluation of carbon fullerene toxicity using embryonic zebrafish. Carbon 2007, 45, 1891–1898. [Google Scholar] [CrossRef]
- Moretz, J.A.; Martins, E.P.; Robison, B.D. Behavioral syndromes and the evolution of correlated behavior in zebrafish. Behav. Ecol. 2007, 18, 556–562. [Google Scholar] [CrossRef]
- Speedie, N.; Gerlai, R. Alarm substance induced behavioral responses in zebrafish (Danio rerio). Behav. Brain Res. 2008, 188, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zheng, Y.; Zhang, Y.; Li, J.; Zhang, H.; Wang, H. Effects of β-diketone antibiotic mixtures on behavior of zebrafish (Danio rerio). Chemosphere 2016, 144, 2195–2205. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.; Raymond, J.; Hester, J.; Kyzar, E.; Gaikwad, S.; Bruce, I.; Fryar, C.; Chanin, S.; Enriquez, J.; Bagawandoss, S. Assessing social behavior phenotypes in adult zebrafish: Shoaling, social preference, and mirror biting tests. In Zebrafish Protocols for Neurobehavioral Research; Springer: Cham, Switzerland, 2012; pp. 231–246. [Google Scholar]
- Glass, A.S.; Dahm, R. The zebrafish as a model organism for eye development. Ophthalmic Res. 2004, 36, 4–24. [Google Scholar] [CrossRef]
- Bault, Z.A.; Peterson, S.M.; Freeman, J.L. Directional and color preference in adult zebrafish: Implications in behavioral and learning assays in neurotoxicology studies. J. Appl. Toxicol. JAT 2015, 35, 1502–1510. [Google Scholar] [CrossRef]
- Park, J.S.; Ryu, J.H.; Choi, T.I.; Bae, Y.K.; Lee, S.; Kang, H.J.; Kim, C.H. Innate color preference of zebrafish and its use in behavioral analyses. Mol. Cells 2016, 39, 750–755. [Google Scholar] [CrossRef] [Green Version]
- Sarasamma, S.; Audira, G.; Juniardi, S.; Sampurna, B.; Liang, S.-T.; Hao, E.; Lai, Y.-H.; Hsiao, C.-D. Zinc Chloride exposure inhibits brain acetylcholine levels, produces neurotoxic signatures, and diminishes memory and motor activities in adult zebrafish. Int. J. Mol. Sci. 2018, 19, 3195. [Google Scholar] [CrossRef] [Green Version]
- López-Olmeda, J.F.; Madrid, J.A.; Sánchez-Vázquez, F.J. Light and temperature cycles as zeitgebers of zebrafish (Danio rerio) circadian activity rhythms. Chronobiol. Int. 2006, 23, 537–550. [Google Scholar] [CrossRef]
- Larner, S.F.; Wang, J.; Goodman, J.; O’Donoghue Altman, M.B.; Xin, M.; Wang, K.K. In vitro neurotoxicity resulting from exposure of cultured neural cells to several types of nanoparticles. J. Cell Death 2017, 10, 1179670717694523. [Google Scholar] [CrossRef] [Green Version]
- Ershova, E.; Sergeeva, V.; Chausheva, A.; Zheglo, D.; Nikitina, V.; Smirnova, T.; Kameneva, L.; Porokhovnik, L.; Kutsev, S.; Troshin, P. Toxic and DNA damaging effects of a functionalized fullerene in human embryonic lung fibroblasts. Mutat. Res. Genetic Toxicol. Environ. Mutagen. 2016, 805, 46–57. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Inomata, A.; Ogata, A.; Nakae, D. Comparative effects of sulfhydryl compounds on target organellae, nuclei and mitochondria, of hydroxylated fullerene-induced cytotoxicity in isolated rat hepatocytes. J. Appl. Toxicol. 2015, 35, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Usenko, C.Y.; Harper, S.L.; Tanguay, R.L. Fullerene C60 exposure elicits an oxidative stress response in embryonic zebrafish. Toxicol. Appl. Pharmacol. 2008, 229, 44–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayes, C.M.; Gobin, A.M.; Ausman, K.D.; Mendez, J.; West, J.L.; Colvin, V.L. Nano-C60 cytotoxicity is due to lipid peroxidation. Biomaterials 2005, 26, 7587–7595. [Google Scholar] [CrossRef] [PubMed]
- Block, M.; Wu, X.; Pei, Z.; Li, G.; Wang, T.; Qin, L.; Wilson, B.; Yang, J.; Hong, J.; Veronesi, B. Nanometer size diesel exhaust particles are selectively toxic to dopaminergic neurons: The role of microglia, phagocytosis, and NADPH oxidase. FASEB J. 2004, 18, 1618–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, S.J.; Crompton, L.A.; Sood, A.; Saunders, M.; Boyle, N.T.; Buckley, A.; Minogue, A.M.; McComish, S.F.; Jiménez-Moreno, N.; Cordero-Llana, O. Nanoparticle-induced neuronal toxicity across placental barriers is mediated by autophagy and dependent on astrocytes. Nat. Nanotechnol. 2018, 13, 427. [Google Scholar] [CrossRef] [PubMed]
- Schmidel, A.J.; Assmann, K.L.; Werlang, C.C.; Bertoncello, K.T.; Francescon, F.; Rambo, C.L.; Beltrame, G.M.; Calegari, D.; Batista, C.B.; Blaser, R.E. Subchronic atrazine exposure changes defensive behaviour profile and disrupts brain acetylcholinesterase activity of zebrafish. Neurotoxicol. Teratol. 2014, 44, 62–69. [Google Scholar] [CrossRef]
- Weis, J.S.; Candelmo, A. Pollutants and fish predator/prey behavior: A review of laboratory and field approaches. Curr. Zool. 2012, 58, 9–20. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Scott, G.R.; Sloman, K.A. The effects of environmental pollutants on complex fish behaviour: Integrating behavioural and physiological indicators of toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef]
- MohanKumar, S.M.; Campbell, A.; Block, M.; Veronesi, B. Particulate matter, oxidative stress and neurotoxicity. Neurotoxicology 2008, 29, 479–488. [Google Scholar] [CrossRef]
- De Lorenzo, A.D. The olfactory neuron and the blood-brain barrier. In Ciba Foundation Symposium-Internal Secretions of the Pancreas (Colloquia on Endocrinology); John Wiley and Sons: Chichester, UK, 1970; pp. 151–176. [Google Scholar]
- Long, T.C.; Saleh, N.; Tilton, R.D.; Lowry, G.V.; Veronesi, B. Titanium dioxide (P25) produces reactive oxygen species in immortalized brain microglia (BV2): Implications for nanoparticle neurotoxicity. Environ. Sci. Technol. 2006, 40, 4346–4352. [Google Scholar] [CrossRef] [PubMed]
- Gernhöfer, M.; Pawert, M.; Schramm, M.; Müller, E.; Triebskorn, R. Ultrastructural biomarkers as tools to characterize the health status of fish in contaminated streams. J. Aquat. Ecosyst. Stress Recovery 2001, 8, 241–260. [Google Scholar] [CrossRef]
- Gerlai, R. Phenomics: Fiction or the future? Trends Neurosci. 2002, 25, 506–509. [Google Scholar] [CrossRef]
- Stewart, A.M.; Kaluyeva, A.A.; Poudel, M.K.; Nguyen, M.; Song, C.; Kalueff, A.V. Building zebrafish neurobehavioral phenomics: Effects of common environmental factors on anxiety and locomotor activity. Zebrafish 2015, 12, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Collymore, C.; Tolwani, R.J.; Rasmussen, S. The behavioral effects of single housing and environmental enrichment on adult zebrafish (Danio rerio). J. Am. Assoc. Lab. Animal Sci. 2015, 54, 280–285. [Google Scholar]
- James, A.M.; Yau-Huei, W.; Cheng-Yoong, P.; MURPHY, M.P. Altered mitochondrial function in fibroblasts containing MELAS or MERRF mitochondrial DNA mutations. Biochem. J. 1996, 318, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Eder, K.J.; Clifford, M.A.; Hedrick, R.P.; Köhler, H.-R.; Werner, I. Expression of immune-regulatory genes in juvenile Chinook salmon following exposure to pesticides and infectious hematopoietic necrosis virus (IHNV). Fish Shellfish Immunol. 2008, 25, 508–516. [Google Scholar] [CrossRef]
- Jin, Y.; Pan, X.; Cao, L.; Ma, B.; Fu, Z. Embryonic exposure to cis-bifenthrin enantioselectively induces the transcription of genes related to oxidative stress, apoptosis and immunotoxicity in zebrafish (Danio rerio). Fish Shellfish Immunol. 2013, 34, 717–723. [Google Scholar] [CrossRef]
- Dandekar, A.; Mendez, R.; Zhang, K. Cross talk between ER stress, oxidative stress, and inflammation in health and disease. In Stress Responses; Springer: Cham, Switzerland, 2015; pp. 205–214. [Google Scholar]
- Condello, C.; Yuan, P.; Schain, A.; Grutzendler, J. Microglia constitute a barrier that prevents neurotoxic protofibrillar Aβ42 hotspots around plaques. Nat. Commun. 2015, 6, 6176. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Lu, J.; Zhao, D. Effects of starch-coating of magnetite nanoparticles on cellular uptake, toxicity and gene expression profiles in adult zebrafish. Sci. Total Environ. 2018, 622, 930–941. [Google Scholar] [CrossRef]
- Isaacson, C.W.; Usenko, C.Y.; Tanguay, R.L.; Field, J.A. Quantification of fullerenes by LC/ESI-MS and its application to in vivo toxicity assays. Anal. Chem. 2007, 79, 9091–9097. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Jang, M.-H.; Kim, J.-Y.; Kim, S.D. Effect of preparation methods on toxicity of fullerene water suspensions to Japanese medaka embryos. Sci. Total Environ. 2010, 408, 5606–5612. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A versatile setup for measuring multiple behavior endpoints in zebrafish. Inventions 2018, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Escudero, A.; Vicente-Page, J.; Hinz, R.C.; Arganda, S.; De Polavieja, G.G. idTracker: Tracking individuals in a group by automatic identification of unmarked animals. Nat. Methods 2014, 11, 743. [Google Scholar] [CrossRef]
- Weber, D.N.; Hoffmann, R.G.; Hoke, E.S.; Tanguay, R.L. Bisphenol A exposure during early development induces sex-specific changes in adult zebrafish social interactions. J. Toxicol. Environ. Health Part A 2015, 78, 50–66. [Google Scholar] [CrossRef] [Green Version]
- Audira, G.; Sarasamma, S.; Chen, J.-R.; Juniardi, S.; Sampurna, B.; Liang, S.-T.; Lai, Y.-H.; Lin, G.-M.; Hsieh, M.-C.; Hsiao, C.-D. Zebrafish Mutants carrying leptin a (lepa) gene deficiency display obesity, anxiety, less aggression and fear, and circadian rhythm and color preference dysregulation. Int. J. Mol. Sci. 2018, 19, 4038. [Google Scholar] [CrossRef] [Green Version]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Han, L.; Hsiao, C.-D. Establishing simple image-based methods and a cost-effective instrument for toxicity assessment on circadian rhythm dysregulation in fish. Biol. Open 2019, 8, bio041871. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarker | WT (n = 10) | C70 (0.5 ppm) | C70 (1.5 ppm) | Unit | Significance | ANOVA F Value | p Value |
|---|---|---|---|---|---|---|---|
| Muscle | |||||||
| ROS | 2.95 ± 0.12 | 5.62 ± 0.26 *** | 8.46 ± 0.25 **** | U/ug of total protein | YES | F (2, 6) = 157.2 | p < 0.0001 |
| CAT | 3.03 ± 0.13 | 5.01 ± 0.44 ** | 3.82 ± 0.01 NS | U/ug of total protein | YES | F (2, 6) = 14.10 | p = 0.0054 |
| SOD | 6.19 ± 0.78 | 14.38 ± 0.34 *** | 14.34 ± 0.54 *** | U/ug of total protein | YES | F (2, 6) = 66.33 | p < 0.0001 |
| TBARS | 5.60 ± 0.34 | 7.39 ± 0.36 * | 8.37 ± 0.29 ** | ng/ug of total protein | YES | F (2, 6) = 17.92 | p = 0.0029 |
| MDA | 0.15 ± 0.00 | 0.20 ± 0.01 * | 0.22 ± 0.01 * | ng/ug of total protein | YES | F (2, 6) = 8.826 | p = 0.0163 |
| Cortisol | 17.95 ± 0.90 | 20.87 ± 1.17 NS | 23.95 ± 0.66 ** | pg/ug of total protein | YES | F (2, 6) = 10.3 | p = 0.0114 |
| Hif1-α | 11.65 ± 0.54 | 17.60 ± 0.87 ** | 18.45 ± 1.00 ** | pg/ug of total protein | YES | F (2, 6) = 20.05 | p = 0.0022 |
| ssDNA | 0.61 ± 0.01 | 0.81 ± 0.07 * | 0.89 ± 0.05 ** | U/ug of total protein | YES | F (2, 6) = 9.408 | p = 0.0141 |
| TNF-α | 6.86 ± 0.41 | 8.90 ± 0.33 ** | 11.76 ± 0.26 *** | pg/ug of total protein | YES | F (2, 6) = 52.24 | p = 0.0002 |
| IL1-β | 0.42 ± 0.05 | 0.47 ± 0.02 NS | 0.57 ± 0.03 * | ng/ug of total protein | YES | F (2, 6) = 5.946 | p = 0.0377 |
| ATP | 363.60 ± 9.02 | 343.60 ± 15.23 NS | 282.20 ± 9.30 ** | ng/ug of total protein | YES | F (2, 6) = 13.5 | p = 0.0060 |
| CK | 3.06 ± 0.14 | 4.78 ± 0.34 ** | 2.79 ± 0.14 NS | pg/ug of total protein | YES | F (2, 6) = 22.17 | p = 0.0017 |
| Brain | |||||||
| ROS | 0.90 ± 0.03 | 1.61 ± 0.19 NS | 3.53 ± 0.64 ** | U/ug of total protein | YES | F (2, 6) = 12.38 | p = 0.0074 |
| SOD | 2.20 ± 0.06 | 3.44 ± 0.08 **** | 3.28 ± 0.09 *** | U/ug of total protein | YES | F (2, 6) = 73.1 | p < 0.0001 |
| Cortisol | 3.47 ± 0.13 | 4.76 ± 0.08 * | 4.91 ± 0.51 * | pg/ug of total protein | YES | F (2, 6) = 6.561 | p = 0.0309 |
| Hif1-α | 4.26 ± 0.11 | 4.91 ± 0.16 NS | 6.86 ± 0.37 *** | pg/ug of total protein | YES | F (2, 6) = 32.09 | p = 0.0006 |
| ssDNA | 0.13 ± 0.01 | 0.28 ± 0.09 NS | 0.37 ± 0.02 * | U/ug of total protein | YES | F (2, 6) = 5.274 | p = 0.0477 |
| ACh | 4.44 ± 0.15 | 4.61 ± 0.08 NS | 2.93 ± 0.31 ** | U/ug of total protein | YES | F (2, 6) = 20.92 | p = 0.0020 |
| AChE | 0.70 ± 0.10 | 0.77 ± 0.02 NS | 1.03 ± 0.08 ** | U/ug of total protein | YES | F (2, 6) = 12.75 | p = 0.0069 |
| Melatonin | 1.02 ± 0.03 | 0.94 ± 0.04 NS | 0.68 ± 0.03 *** | pg/ug of total protein | YES | F (2, 6) = 31.66 | p = 0.0006 |
| Serotonin | 0.20 ± 0.01 | 0.22 ± 0.01 NS | 0.14 ± 0.00 ** | ng/ug of total protein | YES | F (2, 6) = 31.05 | p = 0.0007 |
| Dopamine | 9.04 ± 0.08 | 9.30 ± 0.20 NS | 6.92 ± 0.09 **** | pg/ug of total protein | YES | F (2, 6) = 95.27 | p < 0.0001 |
| COX-1 | 0.16 ± 0.00 | 0.18 ± 0.00 NS | 0.38 ± 0.01 **** | U/pg of total protein | YES | F (2, 6) = 258.2 | p < 0.0001 |
| COX-2 | 0.74 ± 0.01 | 0.94 ± 0.12 NS | 0.64 ± 0.03 NS | U/pg of total protein | YES | F (2, 6) = 4.32 | p = 0.0688 |
| Gills | |||||||
| CAT | 1.44 ± 0.03 | 1.64 ± 0.02 ** | 1.30 ± 0.03 * | U/ug of total protein | YES | F (2, 6) = 44.21 | p = 0.0003 |
| TBARS | 1.71 ± 0.14 | 1.64 ± 0.06 NS | 2.30 ± 0.68 NS | ng/ug of total protein | NO | F (2, 6) = 0.821 | p = 0.4842 |
| MDA | 0.08 ± 0.00 | 0.09 ± 0.00 NS | 0.11 ± 0.00 ** | ng/ug of total protein | YES | F (2, 6) = 21.29 | p = 0.0019 |
| TNF-α | 3.06 ± 0.10 | 2.98 ± 0.03 NS | 8.68 ± 0.74 *** | pg/ug of total protein | YES | F (2, 6) = 56.94 | p = 0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarasamma, S.; Audira, G.; Samikannu, P.; Juniardi, S.; Siregar, P.; Hao, E.; Chen, J.-R.; Hsiao, C.-D. Behavioral Impairments and Oxidative Stress in the Brain, Muscle, and Gill Caused by Chronic Exposure of C70 Nanoparticles on Adult Zebrafish. Int. J. Mol. Sci. 2019, 20, 5795. https://doi.org/10.3390/ijms20225795
Sarasamma S, Audira G, Samikannu P, Juniardi S, Siregar P, Hao E, Chen J-R, Hsiao C-D. Behavioral Impairments and Oxidative Stress in the Brain, Muscle, and Gill Caused by Chronic Exposure of C70 Nanoparticles on Adult Zebrafish. International Journal of Molecular Sciences. 2019; 20(22):5795. https://doi.org/10.3390/ijms20225795
Chicago/Turabian StyleSarasamma, Sreeja, Gilbert Audira, Prabu Samikannu, Stevhen Juniardi, Petrus Siregar, Erwei Hao, Jung-Ren Chen, and Chung-Der Hsiao. 2019. "Behavioral Impairments and Oxidative Stress in the Brain, Muscle, and Gill Caused by Chronic Exposure of C70 Nanoparticles on Adult Zebrafish" International Journal of Molecular Sciences 20, no. 22: 5795. https://doi.org/10.3390/ijms20225795
APA StyleSarasamma, S., Audira, G., Samikannu, P., Juniardi, S., Siregar, P., Hao, E., Chen, J.-R., & Hsiao, C.-D. (2019). Behavioral Impairments and Oxidative Stress in the Brain, Muscle, and Gill Caused by Chronic Exposure of C70 Nanoparticles on Adult Zebrafish. International Journal of Molecular Sciences, 20(22), 5795. https://doi.org/10.3390/ijms20225795