Current and Future Concepts for the Treatment of Impaired Fracture Healing

 , ,
, ,
Abstract
:1. Clinical Challenge of Impaired Fracture Healing
2. Basic Biological Principles of Bone Regeneration
2.1. Hematoma Formation and Inflammatory Response
2.2. Proliferation and Differentiation
2.3. Ossification
2.4. Remodeling Phase
2.5. Mechanical Aspects of Fracture Healing
3. Current State-of-the-Art Therapy for Impaired Fracture Healing
3.1. Case History, Physical and Radiological Examination
3.2. Current Treatment Concepts
3.3. Conservative Therapy
3.4. Surgical Therapy
4. Novel Approaches for Impaired Fracture Healing
4.1. Molecular Targets for Local and/or Systemic Application
4.1.1. Parathyroid Hormone
4.1.2. Wnt Signaling and Sclerostin
4.1.3. Bone Morphogenic Proteins
4.1.4. Platelet Derived Growth Factor
4.1.5. Fibroblast Growth Factor
4.2. Novel Bone Grafts
4.2.1. Calcium-Based Bone Grafts
4.2.2. Bioactive Glass
4.2.3. Organic Bone Grafts
4.2.4. Innovative Biomaterials
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BMD | Bone mineral density |
| BMP | Bone morphogenic proteins |
| CPC | Calcium phosphate cements |
| DBM | Demineralized bone matrix |
| Dlx5 | Distal-less homebox 5 |
| ESTROT | European Society of Tissue Regeneration in Orthopedics and Traumatology |
| ESWT | Extracorporal shock wave therapy |
| FDA | Food and Drug Administration |
| FGF | Fibroblast growth factor |
| FGFR | FGF receptors |
| Fz | Frizzled receptor |
| GF/PL | Gas forming and particulate leaching |
| HA | Hydroxyapatite |
| IGF | Insulin like growth factor |
| IL | Interleukin |
| LIPUS | Low-intensity pulsed ultrasound |
| LRP5/6 | LDL receptor-related protein |
| MAPK | Mitogen-activated protein kinase |
| MDSC | Muscle-derived stomal cell |
| MSC | Mesenchymal stem cells |
| NBP | NEMO binding domain peptide |
| NCPs | Non-collagenous proteins |
| PDC | Periosteum-derived progenitor cells |
| PDGF | Platelet derived growth factor |
| PDGFRβ | PDGF receptor β |
| PLGA | Poly(lactic-co-glycolic acid) |
| PLLA | Poly(l-lactic acid) |
| PTH | Parathyroid hormone |
| PTHrP | Parathyroid hormone related protein |
| rhBMP-2 | Recombinant human BMP-2 |
| rhFGF-2 | Recombinant human FGF-2 |
| rhPDGF-BB | Recombinant human PDGF-BB homodimer |
| RIA | Reamer-Irrigator-Aspirator System |
| Runx2 | Runt-related transcription factor 2 |
| SC/PL | Solvent casting and particulate leaching |
| TCP | Tricalcium phosphate |
| TGF-β | Transforming growth factor-β |
| TNF-α | Tumor necrosis factor-α |
References
- Zura, R.; Xiong, Z.; Einhorn, T.; Watson, J.T.; Ostrum, R.F.; Prayson, M.J.; Della Rocca, G.J.; Metha, S.; McKinley, T.; Wang, Z.; et al. Epidemiology of fracture nonunion in 18 human bones. JAMA Surgery 2016, 151, e162775. [Google Scholar] [CrossRef] [PubMed]
- Antonova, E.; Le, T.K.; Burge, R.; Mershon, J. Tibia shaft fractures: Costly burden of nonunions. BMC Musculoskelet. Disord. 2013, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Rupp, M.; Biehl, C.; Budak, M.; Thormann, U.; Heiss, C.; Alt, V. Diaphyseal long bone nonunions — types, aetiology, economics, and treatment recommendations. Int. Orthop. 2018, 42, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Schmidmaier, G.; Moghaddam, A. Pseudarthrosen langer Röhrenknochen. Z. Orthop. Unfall. 2015, 153, 659–676. [Google Scholar] [CrossRef]
- Calori, G.; Mazza, E.L.; Mazzola, S.; Colombo, A.; Giardina, F.; Romanò, F.; Colombo, M. Non-unions. Clin. Cases Miner. Bone Metab. 2017, 14, 186. [Google Scholar] [CrossRef]
- Weber, B.; Čech, O. Pseudarthrosen: Pathophysiologie, Biomechanik, Therapie, Ergebnisse; Huber: Edison, NJ, USA, 1973. [Google Scholar]
- Einhorn, T.A. The cell and molecular biology of fracture healing. Clin. Orthop. Relat. Res. 1998, 355, 7–21. [Google Scholar] [CrossRef]
- Andrzejowski, P.; Giannoudis, P. V The “diamond concept” for long bone non-union management. J. Orthop. Traumatol. 2019, 20, 21. [Google Scholar] [CrossRef]
- McKibbin, B. The biology of fracture healing in long bones. J. Bone Joint Surg. Br. 1978, 60-B, 150–162. [Google Scholar] [CrossRef]
- Harwood, P.J.; Ferguson, D.O. (ii) An update on fracture healing and non-union. Orthop. Trauma 2015, 29, 228–242. [Google Scholar] [CrossRef]
- Kolar, P.; Schmidt-Bleek, K.; Schell, H.; Gaber, T.; Toben, D.; Schmidmaier, G.; Perka, C.; Buttgereit, F.; Duda, G.N. The early fracture hematoma and its potential role in fracture healing. Tissue Eng. Part B Rev. 2010, 16, 427–434. [Google Scholar] [CrossRef]
- Grundnes, O.; Reikerås, O. the Importance of the Hematoma for Fracture Healing in Rats. Acta Orthop. 1993, 64, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Mineo, K.; Tachibana, T.; Sumi, M.; Matsubara, T.; Hirohata, K. The osteogenetic potential of fracture haematoma. Subperiosteal and intramuscular transplantation of the haematoma. J. Bone Joint Surg. Br. 1990, 72, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Young, R.W. Cell proliferation and specialization during endochondral osteogenesis in young rats. J. Cell Biol. 1962, 14, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Bielby, R.; Jones, E.; McGonagle, D. The role of mesenchymal stem cells in maintenance and repair of bone. Injury 2007, 38, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Oe, K.; Miwa, M.; Sakai, Y.; Lee, S.Y.; Kuroda, R.; Kurosaka, M. An in vitro study demonstrating that haematomas found at the site of human fractures contain progenitor cells with multilineage capacity. J. Bone Jt. Surg. Ser. B 2007, 89, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M. Phylogenetic and functional relationships between coagulation and the innate immune response. Crit. Care Med. 2000, 28, 77–80. [Google Scholar] [CrossRef]
- Einhorn, T.A.; Majeska, R.J.; Rush, E.B.; Levine, P.M.; Horowitz, M.C. The expression of cytokine activity by fracture callus. J. Bone Miner. Res. 1995, 10, 1272–1281. [Google Scholar] [CrossRef]
- Schmidt-Bleek, K.; Schell, H.; Lienau, J.; Schulz, N.; Hoff, P.; Pfaff, M.; Schmidt, G.; Martin, C.; Perka, C.; Buttgereit, F.; et al. Initial immune reaction and angiogenesis in bone healing. J. Tissue Eng. Regen. Med. 2014, 8, 120–130. [Google Scholar] [CrossRef]
- Yang, X.; Ricciardi, B.F.; Hernandez-Soria, A.; Shi, Y.; Pleshko Camacho, N.; Bostrom, M.P.G. Callus mineralization and maturation are delayed during fracture healing in interleukin-6 knockout mice. Bone 2007, 41, 928–936. [Google Scholar] [CrossRef]
- Wallace, A.; Cooney, T.E.; Englund, R.; Lubahn, J.D. Effects of interleukin-6 ablation on fracture healing in mice. J. Orthop. Res. 2011, 29, 1437–1442. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cho, T.J.; Kon, T.; Aizawa, T.; Tsay, A.; Fitch, J.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Impaired fracture healing in the absence of TNF-α signaling: The role of TNF-α in endochondral cartilage resorption. J. Bone Miner. Res. 2003, 18, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Glass, G.E.; Chan, J.K.; Freidin, A.; Feldmann, M.; Horwood, N.J.; Nanchahal, J. TNF-α promotes fracture repair by augmenting the recruitment and differentiation of muscle-derived stromal cells. Proc. Natl. Acad. Sci. USA 2011, 108, 1585–1590. [Google Scholar] [CrossRef]
- Brighton, C.T.; Hunt, R.M. Early histological and ultrastructural changes in medullary fracture callus. J. Bone Jt. Surg. Ser. A 1991, 73, 832–847. [Google Scholar] [CrossRef]
- Lieberman, J.R.; Daluiski, A.; Einhorn, T.A. The role of growth factors in the repair of bone biology and clinical applications. J. Bone Jt. Surg. Ser. A 2002, 84, 1032–1044. [Google Scholar] [CrossRef]
- Ford, J.L.; Robinson, D.E.; Scammell, B.E. The fate of soft callus chondrocytes during long bone fracture repair. J. Orthop. Res. 2003, 21, 54–61. [Google Scholar] [CrossRef]
- Manolagas, S.C. Birth and death of bone cells: Basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr. Rev. 2000, 21, 115–137. [Google Scholar]
- Schenk, R.; Willenegger, H. [On the histology of primary bone healing]. Langenbecks Arch. Klin. Chir. Ver. Dtsch. Z. Chir. 1964, 308, 440–452. [Google Scholar]
- Gerstenfeld, L.C.; Alkhiary, Y.M.; Krall, E.A.; Nicholls, F.H.; Stapleton, S.N.; Fitch, J.L.; Bauer, M.; Kayal, R.; Graves, D.T.; Jepsen, K.J.; et al. Three-dimensional reconstruction of fracture callus morphogenesis. J. Histochem. Cytochem. 2006, 54, 1215–1228. [Google Scholar] [CrossRef]
- Marsell, R.; Einhorn, T.A. The biology of fracture healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef]
- Thompson, Z.; Miclau, T.; Hu, D.; Helms, J.A. A model for intramembranous ossification during fracture healing. J. Orthop. Res. 2002, 20, 1091–1098. [Google Scholar] [CrossRef]
- Calori, G.M.; Phillips, M.; Jeetle, S.; Tagliabue, L.; Giannoudis, P.V. Classification of non-union: Need for a new scoring system? Injury 2008, 39, S59–S63. [Google Scholar] [CrossRef]
- Gebauer, D.; Mayr, E.; Orthner, E.; Ryaby, J.P. Low-intensity pulsed ultrasound: Effects on nonunions. Ultrasound Med. Biol. 2005, 31, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Schoellner, C.; Rompe, J.-D.; Decking, J.; Heine, J. Die hochenergetische extrakorporale Stoßwellentherapie (ESWT) bei Pseudarthrose. Orthopade 2002, 31, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Lou, S.; Lv, H.; Li, Z.; Zhang, L.; Tang, P. The effects of low-intensity pulsed ultrasound on fresh fracture. Medicine (Baltimore) 2017, 96, e8181. [Google Scholar] [CrossRef]
- Leighton, R.; Watson, J.T.; Giannoudis, P.; Papakostidis, C.; Harrison, A.; Steen, R.G. Healing of fracture nonunions treated with low-intensity pulsed ultrasound (LIPUS): A systematic review and meta-analysis. Injury 2017, 48, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Schlickewei, W.; Schlickewei, C. The Use of Bone Substitutes in the Treatment of Bone Defects – the Clinical View and History. Macromol. Symp. 2007, 253, 10–23. [Google Scholar] [CrossRef]
- Cox, G.; McGonagle, D.; Boxall, S.A.; Buckley, C.T.; Jones, E.; Giannoudis, P.V. The use of the reamer-irrigator-aspirator to harvest mesenchymal stem cells. J. Bone Joint Surg. Br. 2011, 93-B, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Madison, R.D.; Nowotarski, P.J. The Reamer-Irrigator-Aspirator in Nonunion Surgery. Orthop. Clin. N. Am. 2019, 50, 297–304. [Google Scholar] [CrossRef]
- Calori, G.M.; Colombo, M.; Mazza, E.L.; Mazzola, S.; Malagoli, E.; Mineo, G.V. Incidence of donor site morbidity following harvesting from iliac crest or RIA graft. Injury 2014, 45, S116–S120. [Google Scholar] [CrossRef]
- Schmidmaier, G.; Herrmann, S.; Green, J.; Weber, T.; Scharfenberger, A.; Haas, N.P.; Wildemann, B. Quantitative assessment of growth factors in reaming aspirate, iliac crest, and platelet preparation. Bone 2006, 39, 1156–1163. [Google Scholar] [CrossRef]
- Laurencin, C.; Khan, Y.; El-Amin, S.F. Bone graft substitutes. Expert Rev. Med. Devices 2006, 3, 49–57. [Google Scholar] [CrossRef]
- Herrera, M.; Chapman, C.B.; Roh, M.; Strauch, R.J.; Rosenwasser, M.P. Treatment of unstable distal radius fractures with cancellous allograft and external fixation. J. Hand Surg. Am. 1999, 24, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Lasanianos, N.; Mouzopoulos, G.; Garnavos, C. The use of freeze-dried cancelous allograft in the management of impacted tibial plateau fractures. Injury 2008, 39, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Flierl, M.A.; Smith, W.R.; Mauffrey, C.; Irgit, K.; Williams, A.E.; Ross, E.; Peacher, G.; Hak, D.J.; Stahel, P.F. Outcomes and complication rates of different bone grafting modalities in long bone fracture nonunions: A retrospective cohort study in 182 patients. J. Orthop. Surg. Res. 2013, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef]
- Hierholzer, C.; Sama, D.; Toro, J.B.; Peterson, M.; Helfet, D.L. Plate Fixation of Ununited Humeral Shaft Fractures. J. Bone Jt. Surg. 2006, 88, 1442–1447. [Google Scholar]
- Peterson, R.S.; Andhare, R.A.; Rousche, K.T.; Knudson, W.; Wang, W.; Grossfield, J.B.; Thomas, R.O.; Hollingsworth, R.E.; Knudson, C.B. CD44 modulates Smad1 activation in the BMP-7 signaling pathway. J. Cell Biol. 2004, 166, 1081–1091. [Google Scholar] [CrossRef]
- Wildemann, B.; Burkhardt, N.; Luebberstedt, M.; Vordemvenne, T.; Schmidmaier, G. Proliferating and differentiating effects of three different growth factors on pluripotent mesenchymal cells and osteoblast like cells. J. Orthop. Surg. Res. 2007, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Masquelet, A.C.; Fitoussi, F.; Begue, T.; Muller, G.P. Reconstruction of the long bones by the induced membrane and spongy autograft. In Annales de Chirurgie Plastique et Esthetique; Elsevier: Paris, France, 2000; Volume 45, pp. 346–353. [Google Scholar]
- Morelli, I.; Drago, L.; George, D.A.; Gallazzi, E.; Scarponi, S.; Romanò, C.L. Masquelet technique: Myth or reality? A systematic review and meta-analysis. Injury 2016, 47, S68–S76. [Google Scholar] [CrossRef]
- Silva, B.C.; Costa, A.G.; Cusano, N.E.; Kousteni, S.; Bilezikian, J.P. Catabolic and anabolic actions of parathyroid hormone on the skeleton. J. Endocrinol. Invest. 2011, 34, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobnig, H.; Turner, R.T. The effects of programmed administration of human parathyroid hormone fragment (1–34) on bone histomorphometry and serum chemistry in rats. Endocrinology 1997, 138, 4607–4612. [Google Scholar] [CrossRef] [PubMed]
- Neer, R.M.; Arnaud, C.D.; Zanchetta, J.R.; Prince, R.; Gaich, G.A.; Reginster, J.-Y.; Hodsman, A.B.; Eriksen, E.F.; Ish-Shalom, S.; Genant, H.K.; et al. Effect of Parathyroid Hormone (1-34) on Fractures and Bone Mineral Density in Postmenopausal Women with Osteoporosis. N. Engl. J. Med. 2001, 344, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, S.L.; Bone, H.G.; Ettinger, M.P.; Hanley, D.A.; Lindsay, R.; Zanchetta, J.R.; Blosch, C.M.; Mathisen, A.L.; Morris, S.A.; Marriott, T.B. Effect of Recombinant Human Parathyroid Hormone (1-84) on Vertebral Fracture and Bone Mineral Density in Postmenopausal Women with Osteoporosis. Ann. Intern. Med. 2007, 146, 326. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.D.; Hattersley, G.; Riis, B.J.; Williams, G.C.; Lau, E.; Russo, L.A.; Alexandersen, P.; Zerbini, C.A.F.; Hu, M.; Harris, A.G.; et al. Effect of Abaloparatide vs Placebo on New Vertebral Fractures in Postmenopausal Women With Osteoporosis. JAMA 2016, 316, 722. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.D.; Hattersley, G.; Lau, E.; Fitzpatrick, L.A.; Harris, A.G.; Williams, G.C.; Hu, M.-Y.; Riis, B.J.; Russo, L.; Christiansen, C. Bone mineral density response rates are greater in patients treated with abaloparatide compared with those treated with placebo or teriparatide: Results from the ACTIVE phase 3 trial. Bone 2019, 120, 137–140. [Google Scholar] [CrossRef]
- Jilka, R.L. Molecular and cellular mechanisms of the anabolic effect of intermittent PTH. Bone 2007, 40, 1434–1446. [Google Scholar] [CrossRef] [Green Version]
- Nozaka, K.; Miyakoshi, N.; Kasukawa, Y.; Maekawa, S.; Noguchi, H.; Shimada, Y. Intermittent administration of human parathyroid hormone enhances bone formation and union at the site of cancellous bone osteotomy in normal and ovariectomized rats. Bone 2008, 42, 90–97. [Google Scholar] [CrossRef]
- Aspenberg, P.; Genant, H.K.; Johansson, T.; Nino, A.J.; See, K.; Krohn, K.; García-Hernández, P.A.; Recknor, C.P.; Einhorn, T.A.; Dalsky, G.P.; et al. Teriparatide for acceleration of fracture repair in humans: A prospective, randomized, double-blind study of 102 postmenopausal women with distal radial fractures. J. Bone Miner. Res. 2010, 25, 404–414. [Google Scholar] [CrossRef]
- Peichl, P.; Holzer, L.A.; Maier, R.; Holzer, G. Parathyroid hormone 1-84 accelerates fracture-healing in pubic bones of elderly osteoporotic women. J. Bone Jt. Surg. Ser. A 2011, 93, 1583–1587. [Google Scholar] [CrossRef] [Green Version]
- Bukata, S.V.; Puzas, J.E. Orthopedic Uses of Teriparatide. Curr. Osteoporos. Rep. 2010, 8, 28–33. [Google Scholar] [CrossRef]
- Zati, A.; Sarti, D.; Malaguti, M.C.; Pratelli, L.; Rohe, M.E.; Krege, J.H. Teriparatide in the treatment of a loose hip prosthesis. J. Rheumatol. 2011, 38, 778–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhao, J.J.; Mitlak, B.H.; Wang, O.; Genant, H.K.; Eriksen, E.F. Recombinant Human Parathyroid Hormone (1-34) [Teriparatide] Improves Both Cortical and Cancellous Bone Structure. J. Bone Miner. Res. 2003, 18, 1932–1941. [Google Scholar] [CrossRef] [PubMed]
- Ogura, K.; Iimura, T.; Makino, Y.; Sugie-Oya, A.; Takakura, A.; Takao-Kawabata, R.; Ishizuya, T.; Moriyama, K.; Yamaguchi, A. Short-term intermittent administration of parathyroid hormone facilitates osteogenesis by different mechanisms in cancellous and cortical bone. Bone Rep. 2016, 5, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Bellido, T.; Ali, A.A.; Gubrij, I.; Plotkin, L.I.; Fu, Q.; O’Brien, C.A.; Manolagas, S.C.; Jilka, R.L. Chronic elevation of parathyroid hormone in mice reduces expression of sclerostin by osteocytes: A novel mechanism for hormonal control of osteoblastogenesis. Endocrinology 2005, 146, 4577–4583. [Google Scholar] [CrossRef]
- Ten Dijke, P.; Krause, C.; de Gorter, D.J.; Löwik, C.W.; van Bezooijen, R.L. Osteocyte-Derived Sclerostin Inhibits Bone Formation: Its Role in Bone Morphogenetic Protein and Wnt Signaling. J. Bone Jt. Surgery-Am. Vol. 2008, 90, 31–35. [Google Scholar] [CrossRef]
- Delgado-Calle, J.; Tu, X.; Pacheco-Costa, R.; McAndrews, K.; Edwards, R.; Pellegrini, G.G.; Kuhlenschmidt, K.; Olivos, N.; Robling, A.; Peacock, M.; et al. Control of Bone Anabolism in Response to Mechanical Loading and PTH by Distinct Mechanisms Downstream of the PTH Receptor. J. Bone Miner. Res. 2017, 32, 522–535. [Google Scholar] [CrossRef]
- Bennett, C.N.; Longo, K.A.; Wright, W.S.; Suva, L.J.; Lane, T.F.; Hankenson, K.D.; MacDougald, O.A. Regulation of osteoblastogenesis and bone mass by Wnt10b. Proc. Natl. Acad. Sci. USA 2005, 102, 3324–3329. [Google Scholar] [CrossRef] [Green Version]
- Cawthorn, W.P.; Bree, A.J.; Yao, Y.; Du, B.; Hemati, N.; Martinez-Santibañez, G.; MacDougald, O.A. Wnt6, Wnt10a and Wnt10b inhibit adipogenesis and stimulate osteoblastogenesis through a β-catenin-dependent mechanism. Bone 2012, 50, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Whetstone, H.C.; Lin, A.C.; Nadesan, P.; Wei, Q.; Poon, R.; Alman, B.A. Beta-catenin signaling plays a disparate role in different phases of fracture repair: Implications for therapy to improve bone healing. PLoS Med. 2007, 4, e249. [Google Scholar] [CrossRef] [Green Version]
- Kakar, S.; Einhorn, T.A.; Vora, S.; Miara, L.J.; Hon, G.; Wigner, N.A.; Toben, D.; Jacobsen, K.A.; Al-Sebaei, M.O.; Song, M.; et al. Enhanced chondrogenesis and Wnt signaling in PTH-treated fractures. J. Bone Miner. Res. 2007, 22, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.M.; Lee, E.H. Transcriptional regulatory cascades in Runx2-dependent bone development. Tissue Eng. Part B Rev. 2013, 19, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtowicz, A.M.; Templeman, K.L.; Hutmacher, D.W.; Guldberg, R.E.; García, A.J. Runx2 overexpression in bone marrow stromal cells accelerates bone formation in critical-sized femoral defects. Tissue Eng. Part A 2010, 16, 2795–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunkow, M.E.; Gardner, J.C.; Van Ness, J.; Paeper, B.W.; Kovacevich, B.R.; Proll, S.; Skonier, J.E.; Zhao, L.; Sabo, P.J.; Fu, Y.; et al. Bone dysplasia sclerosteosis results from loss of the SOST gene product, a novel cystine knot-containing protein. Am. J. Hum. Genet. 2001, 68, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Balemans, W.; Ebeling, M.; Patel, N.; Van Hul, E.; Olson, P.; Dioszegi, M.; Lacza, C.; Wuyts, W.; Van Den Ende, J.; Willems, P.; et al. Increased bone density in sclerosteosis is due to the deficiency of a novel secreted protein (SOST). Hum. Mol. Genet. 2001, 10, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Cosman, F.; Crittenden, D.B.; Adachi, J.D.; Binkley, N.; Czerwinski, E.; Ferrari, S.; Hofbauer, L.C.; Lau, E.; Lewiecki, E.M.; Miyauchi, A.; et al. Romosozumab Treatment in Postmenopausal Women with Osteoporosis. N. Engl. J. Med. 2016, 375, 1532–1543. [Google Scholar] [CrossRef]
- Agholme, F.; Li, X.; Isaksson, H.; Ke, H.Z.; Aspenberg, P. Sclerostin antibody treatment enhances metaphyseal bone healing in rats. J. Bone Miner. Res. 2010, 25, 2412–2418. [Google Scholar] [CrossRef]
- Wozney, J.M. Overview of bone morphogenetic proteins. Spine (Phila. Pa. 1976) 2002, 27, S2–S8. [Google Scholar] [CrossRef]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone morphogenetic protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-S.; Kim, H.-J.; Li, Q.-L.; Chi, X.-Z.; Ueta, C.; Komori, T.; Wozney, J.M.; Kim, E.-G.; Choi, J.-Y.; Ryoo, H.-M.; et al. Runx2 Is a common target of transforming growth factor beta 1 and bone morphogenetic protein 2, and cooperation between Runx2 and Smad5 induces osteoblast-specific gene expression in the pluripotent mesenchymal precursor cell line C2C12. Mol. Cell. Biol. 2000, 20, 8783–8792. [Google Scholar] [CrossRef] [Green Version]
- Miyama, K.; Yamada, G.; Yamamoto, T.S.; Takagi, C.; Miyado, K.; Sakai, M.; Ueno, N.; Shibuya, H. A BMP-inducible gene, Dlx5, regulates osteoblast differentiation and mesoderm induction. Dev. Biol. 1999, 208, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; De Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Govender, S.; Csimma, C.; Genant, H.K.; Valentin-Opran, A.; Amit, Y.; Arbel, R.; Aro, H.; Atar, D.; Bishay, M.; Börner, M.G.; et al. Recombinant human bone morphogenetic protein-2 for treatment of open tibial fractures: A prospective, controlled, randomized study of four hundred and fifty patients. J. Bone Jt. Surg. Ser. A 2002, 84, 2123–2134. [Google Scholar] [CrossRef]
- Jones, A.L.; Bucholz, R.W.; Bosse, M.J.; Mirza, S.K.; Lyon, T.R.; Webb, L.X.; Pollak, A.N.; Golden, J.D.; Valentin-Opran, A. Recombinant human BMP-2 and allograft compared with autogenous bone graft for reconstruction of diaphyseal tibial fractures with cortical defects: A randomized, controlled trial. J. Bone Jt. Surg. Ser. A 2006. [Google Scholar] [CrossRef]
- Burkus, J.K.; Transfeldt, E.E.; Kitchel, S.H.; Watkins, R.G.; Balderston, R.A. Clinical and radiographic outcomes of anterior lumbar interbody fusion using recombinant human bone morphogenetic protein-2. Spine (Phila. Pa. 1976) 2002, 27, 2396–2408. [Google Scholar] [CrossRef]
- Burkus, J.K.; Gornet, M.F.; Dickman, C.A.; Zdeblick, T.A. Anterior lumbar interbody fusion using rhBMP-2 with tapered interbody cages. J. Spinal Disord. Tech. 2002, 15, 337–349. [Google Scholar] [CrossRef]
- Martin, B.I.; Lurie, J.D.; Tosteson, A.N.A.; Deyo, R.A.; Farrokhi, F.R.; Mirza, S.K. Use of bone morphogenetic protein among patients undergoing fusion for degenerative diagnoses in the United States, 2002 to 2012. Spine J. 2015, 15, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Carragee, E.J.; Hurwitz, E.L.; Weiner, B.K. A critical review of recombinant human bone morphogenetic protein-2 trials in spinal surgery: Emerging safety concerns and lessons learned. Spine J. 2011, 11, 471–491. [Google Scholar] [CrossRef]
- Tannoury, C.A.; An, H.S. Complications with the use of bone morphogenetic protein 2 (BMP-2) in spine surgery. Spine J. 2014, 14, 552–559. [Google Scholar] [CrossRef]
- Boraiah, S.; Paul, O.; Hawkes, D.; Wickham, M.; Lorich, D.G. Complications of recombinant human BMP-2 for treating complex tibial plateau fractures: A preliminary report. Clin. Orthop. Relat. Res. 2009, 467, 3257–3262. [Google Scholar] [CrossRef] [Green Version]
- James, A.W.; LaChaud, G.; Shen, J.; Asatrian, G.; Nguyen, V.; Zhang, X.; Ting, K.; Soo, C. A Review of the clinical side effects of bone morphogenetic protein-2. Tissue Eng. Part B Rev. 2016, 22, 284. [Google Scholar] [CrossRef] [PubMed]
- Rihn, J.A.; Makda, J.; Hong, J.; Patel, R.; Hilibrand, A.S.; Anderson, D.G.; Vaccaro, A.R.; Albert, T.J. The use of RhBMP-2 in single-level transforaminal lumbar interbody fusion: A clinical and radiographic analysis. Eur. Spine J. 2009, 18, 1629–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, B.B.; Bae, H.W.; Kropf, M.A.; Patel, V.V.; Zhao, L.; Wong, P.; Delamarter, R.B. Leakage of rhBMP-2 from absorbable collagen sponges during use in anterior cervical discectomy and fusion quantification and follow-up of clinical and radiographic consequences. Spine J. 2005, 5, S55. [Google Scholar] [CrossRef]
- Kim, H.K.W.; Aruwajoye, O.; Du, J.; Kamiya, N. Local administration of bone morphogenetic protein-2 and bisphosphonate during non-weight-bearing treatment of ischemic osteonecrosis of the femoral head: An experimental investigation in immature pigs. J. Bone Jt. Surgery-Am. Vol. 2014, 96, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Robin, B.N.; Chaput, C.D.; Zeitouni, S.; Rahm, M.D.; Zerris, V.A.; Sampson, H.W. Cytokine-mediated inflammatory reaction following posterior cervical decompression and fusion associated with recombinant human bone morphogenetic protein-2: A case study. Spine (Phila. Pa. 1976) 2010, 35, E1350–E1354. [Google Scholar] [CrossRef] [PubMed]
- Zara, J.N.; Siu, R.K.; Zhang, X.; Shen, J.; Ngo, R.; Lee, M.; Li, W.; Chiang, M.; Chung, J.; Kwak, J.; et al. High doses of bone morphogenetic protein 2 induce structurally abnormal bone and inflammation In vivo. Tissue Eng. Part A 2011, 17, 1389. [Google Scholar] [CrossRef] [Green Version]
- Xiong, C.; Daubs, M.D.; Montgomery, S.R.; Aghdasi, B.; Inoue, H.; Tian, H.; Suzuki, A.; Tan, Y.; Hayashi, T.; Ruangchainikom, M.; et al. BMP-2 adverse reactions treated with human dose equivalent dexamethasone in a rodent model of soft-tissue inflammation. Spine (Phila. Pa. 1976) 2013, 38, 1640–1647. [Google Scholar] [CrossRef]
- Glaeser, J.D.; Salehi, K.; Kanim, L.E.A.; Sheyn, D.; Napier, Z.; Behrens, P.H.; Garcia, L.; Cuéllar, J.M.; Bae, H.W. Anti-Inflammatory Peptide Attenuates Edema and Promotes BMP-2-Induced Bone Formation in Spine Fusion. Tissue Eng. Part A 2018, 24, 1641–1651. [Google Scholar] [CrossRef]
- Andrew, J.G.; Hoyland, J.A.; Freemont, A.J.; Marsh, D.R. Platelet-derived growth factor expression in normally healing human fractures. Bone 1995, 16, 455–460. [Google Scholar] [CrossRef]
- Hollinger, J.O.; Hart, C.E.; Hirsch, S.N.; Lynch, S.; Friedlaender, G.E. Recombinant human platelet-derived growth factor: Biology and clinical applications. J. Bone Jt. Surg. Ser. A 2008, 90, 48–54. [Google Scholar] [CrossRef]
- Fiedler, J.; Röderer, G.; Günther, K.-P.; Brenner, R.E. BMP-2, BMP-4, and PDGF-bb stimulate chemotactic migration of primary human mesenchymal progenitor cells. J. Cell. Biochem. 2002, 87, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Nishimura, M.; Sekiya, K.; Suehiro, F.; Kanawa, M.; Nikawa, H.; Hamada, T.; Kato, Y. Comprehensive analysis of chemotactic factors for bone marrow mesenchymal stem cells. Stem Cells Dev. 2007, 16, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.; Leonidou, A.; Lester, M.; Heliotis, M.; Mantalaris, A.; Tsiridis, E. Investigating the role of PDGF as a potential drug therapy in bone formation and fracture healing. Expert Opin. Investig. Drugs 2009, 18, 1633–1654. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.; Naot, D.; Callon, K.; Porteous, F.; Horne, A.; Wattie, D.; Watson, M.; Cornish, J.; Browett, P.; Grey, A. Imatinib promotes osteoblast differentiation by inhibiting PDGFR signaling and inhibits osteoclastogenesis by both direct and stromal cell-dependent mechanisms. J. Bone Miner. Res. 2007, 22, 1679–1689. [Google Scholar] [CrossRef]
- Tokunaga, A.; Oya, T.; Ishii, Y.; Motomura, H.; Nakamura, C.; Ishizawa, S.; Fujimori, T.; Nabeshima, Y.I.; Umezawa, A.; Kanamori, M.; et al. PDGF receptor β is a potent regulator of mesenchymal stromal cell function. J. Bone Miner. Res. 2008, 23, 1519–1528. [Google Scholar] [CrossRef]
- Wang, X.; Matthews, B.G.; Yu, J.; Novak, S.; Grcevic, D.; Sanjay, A.; Kalajzic, I. PDGF modulates BMP2-induced osteogenesis in periosteal progenitor cells. JBMR Plus 2019, 3, e10127. [Google Scholar] [CrossRef]
- Chen, W.; Baylink, D.J.; Brier-Jones, J.; Neises, A.; Kiroyan, J.B.; Rundle, C.H.; Lau, K.-H.W.; Zhang, X.-B. PDGFB-based stem cell gene therapy increases bone strength in the mouse. Proc. Natl. Acad. Sci. USA 2015, 112, E3893–E3900. [Google Scholar] [CrossRef] [Green Version]
- DiGiovanni, C.W.; Lin, S.S.; Baumhauer, J.F.; Daniels, T.; Younger, A.; Glazebrook, M.; Anderson, J.; Anderson, R.; Evangelista, P.; Lynch, S.E.; et al. Recombinant human platelet-derived growth factor-BB and beta-tricalcium phosphate (rhPDGF-BB/β-TCP): An alternative to autogenous bone graft. J. Bone Jt. Surgery-Am. Vol. 2013, 95, 1184–1192. [Google Scholar] [CrossRef] [Green Version]
- Daniels, T.R.; Younger, A.S.E.; Penner, M.J.; Wing, K.J.; Le, I.L.D.; Russell, I.S.; Lalonde, K.-A.; Evangelista, P.T.; Quiton, J.D.; Glazebrook, M.; et al. Prospective randomized controlled trial of hindfoot and ankle fusions treated with rhPDGF-BB in combination with a β-TCP-collagen matrix. Foot Ankle Int. 2015, 36, 739–748. [Google Scholar] [CrossRef]
- Coffin, J.D.; Homer-Bouthiette, C.; Hurley, M.M. Fibroblast growth factor 2 and its receptors in bone biology and disease. J. Endocr. Soc. 2018, 2, 657–671. [Google Scholar] [CrossRef]
- Charoenlarp, P.; Rajendran, A.K.; Iseki, S. Role of fibroblast growth factors in bone regeneration. Inflamm. Regen. 2017, 37, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, H.; Oka, H.; Jingushi, S.; Izumi, T.; Fukunaga, M.; Sato, K.; Matsushita, T.; Nakamura, K. A local application of recombinant human fibroblast growth factor 2 for tibial shaft fractures: A randomized, placebo-controlled trial. J. Bone Miner. Res. 2010, 25, 2459–2467. [Google Scholar] [CrossRef] [PubMed]
- Mayahara, H.; Ito, T.; Nagai, H.; Miyajima, H.; Tsukuda, R.; Taketomi, S.; Mizoguchi, J.; Kato, K. In vivo stimulation of endosteal bone formation by basic fibroblast growth factor in rats. Growth Factors 1993, 9, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, H.; Kurokawa, T.; Hanada, K.; Hiyama, Y.; Tamura, M.; Ogata, E.; Matsumoto, T. Stimulation of fracture repair by recombinant human basic fibroblast growth factor in normal and streptozotocin-diabetic rats. Endocrinology 1994, 135, 774–781. [Google Scholar] [CrossRef]
- Nakamura, T.; Hara, Y.; Tagawa, M.; Tamura, M.; Yuge, T.; Fukuda, H.; Nigi, H. Recombinant human basic fibroblast growth factor accelerates fracture healing by enhancing callus remodeling in experimental dog tibial fracture. J. Bone Miner. Res. 1998, 13, 942–949. [Google Scholar] [CrossRef]
- Kawaguchi, H.; Jingushi, S.; Izumi, T.; Fukunaga, M.; Matsushita, T.; Nakamura, T.; Mizuno, K.; Nakamura, T.; Nakamura, K. Local application of recombinant human fibroblast growth factor-2 on bone repair: A dose–escalation prospective trial on patients with osteotomy. J. Orthop. Res. 2007, 25, 480–487. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, P.; Li, X.; Doetschman, T.; Coffin, J.D.; Drissi, H.; Hurley, M.M. Exported 18-kDa isoform of fibroblast growth factor-2 is a critical determinant of bone mass in mice. J. Biol. Chem. 2009, 284, 3170–3182. [Google Scholar] [CrossRef] [Green Version]
- Hurley, M.M.; Adams, D.J.; Wang, L.; Jiang, X.; Burt, P.M.; Du, E.; Xiao, L. Accelerated fracture healing in transgenic mice overexpressing an anabolic isoform of fibroblast growth factor 2. J. Cell. Biochem. 2016, 117, 599–611. [Google Scholar] [CrossRef]
- Naganawa, T.; Xiao, L.; Coffin, J.D.; Doetschman, T.; Sabbieti, M.G.; Agas, D.; Hurley, M.M. Reduced expression and function of bone morphogenetic protein-2 in bones of Fgf2 null mice. J. Cell. Biochem. 2008, 103, 1975–1988. [Google Scholar] [CrossRef]
- Van Gastel, N.; Stegen, S.; Stockmans, I.; Moermans, K.; Schrooten, J.; Graf, D.; Luyten, F.P.; Carmeliet, G. Expansion of murine periosteal progenitor cells with fibroblast growth factor 2 reveals an intrinsic endochondral ossification program mediated by bone morphogenetic protein 2. Stem Cells 2014, 32, 2407–2418. [Google Scholar] [CrossRef]
- Hurley, M.M.; Tetradis, S.; Huang, Y.-F.; Hock, J.; Kream, B.E.; Raisz, L.G.; Sabbieti, M.G. Parathyroid hormone regulates the expression of fibroblast growth factor-2 mRNA and fibroblast growth factor receptor mRNA in osteoblastic cells. J. Bone Miner. Res. 1999, 14, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Gronowicz, G.; M. Hurley, M. Fibroblast Growth Factor-2, Bone Homeostasis and Fracture Repair. Curr. Pharm. Des. 2013, 19, 3354–3363. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Xiao, L.; Doetschman, T.; Coffin, D.J.; Hurley, M.M. Fibroblast Growth Factor 2 Stimulation of Osteoblast Differentiation and Bone Formation Is Mediated by Modulation of the Wnt Signaling Pathway. J. Biol. Chem. 2011, 286, 40575–40583. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ito, A.; Sogo, Y.; Li, X.; Tsurushima, H.; Oyane, A. Ascorbate–apatite composite and ascorbate–FGF-2–apatite composite layers formed on external fixation rods and their effects on cell activity in vitro. Acta Biomater. 2009, 5, 2647–2656. [Google Scholar] [CrossRef]
- Rhee, Y.; Bivi, N.; Farrow, E.; Lezcano, V.; Plotkin, L.I.; White, K.E.; Bellido, T. Parathyroid hormone receptor signaling in osteocytes increases the expression of fibroblast growth factor-23 in vitro and in vivo. Bone 2011, 49, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Makrythanasis, P.; Temtamy, S.; Aglan, M.S.; Otaify, G.A.; Hamamy, H.; Antonarakis, S.E. A novel homozygous mutation in FGFR3 causes tall stature, severe lateral tibial deviation, scoliosis, hearing impairment, camptodactyly, and arachnodactyly. Hum. Mutat. 2014, 35, 959–963. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Legeai-Mallet, L. Achondroplasia: Development, pathogenesis, and therapy. Dev. Dyn. 2017, 246, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Rundle, C.H.; Miyakoshi, N.; Ramirez, E.; Wergedal, J.E.; Lau, K.-H.W.; Baylink, D.J. Expression of the fibroblast growth factor receptor genes in fracture repair. Clin. Orthop. Relat. Res. 2002, 403, 253–263. [Google Scholar] [CrossRef]
- Valverde-Franco, G.; Liu, H.; Davidson, D.; Chai, S.; Valderrama-Carvajal, H.; Goltzman, D.; Ornitz, D.M.; Henderson, J.E. Defective bone mineralization and osteopenia in young adult FGFR3-/- mice. Hum. Mol. Genet. 2003, 13, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Su, N.; Sun, Q.; Li, C.; Lu, X.; Qi, H.; Chen, S.; Yang, J.; Du, X.; Zhao, L.; He, Q.; et al. Gain-of-function mutation in FGFR3 in mice leads to decreased bone mass by affecting both osteoblastogenesis and osteoclastogenesis. Hum. Mol. Genet. 2010, 19, 1199–1210. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Luo, F.; Xu, W.; Wang, Z.; Sun, X.; Xu, M.; Huang, J.; Zhang, D.; Tan, Q.; Chen, B.; et al. FGFR3 deficient mice have accelerated fracture repair. Int. J. Biol. Sci. 2017, 13, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, N.; Li, X.; Tang, Y.; Yang, J.; Wen, X.; Guo, J.; Tang, J.; Du, X.; Chen, L. Deletion of FGFR3 in osteoclast lineage cells results in increased bone mass in mice by inhibiting osteoclastic bone resorption. J. Bone Miner. Res. 2016, 31, 1676–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Yang, Z.; Li, Y.; Zhang, X.; De Bruijn, J.D.; De Groot, K. Osteoinduction by calcium phosphate biomaterials. J. Mater. Sci. Mater. Med. 1998, 9, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Zwingenberger, S.; Nich, C.; Valladares, R.D.; Yao, Z.; Stiehler, M.; Goodman, S.B. Recommendations and considerations for the use of biologics in orthopedic surgery. BioDrugs 2012, 26, 245–256. [Google Scholar] [CrossRef]
- Beuerlein, M.J.S.; McKee, M.D. Calcium sulfates: What Is the evidence? J. Orthop. Trauma 2010, 24, S46–S51. [Google Scholar] [CrossRef]
- Rizzi, S.C.; Heath, D.J.; Coombes, A.G.A.; Bock, N.; Textor, M.; Downes, S. Biodegradable polymer/hydroxyapatite composites: Surface analysis and initial attachment of human osteoblasts. J. Biomed. Mater. Res. 2001, 55, 475–486. [Google Scholar] [CrossRef]
- Klein, C.P.A.T.; Driessen, A.A.; de Groot, K.; van den Hooff, A. Biodegradation behavior of various calcium phosphate materials in bone tissue. J. Biomed. Mater. Res. 1983, 17, 769–784. [Google Scholar] [CrossRef]
- Daculsi, G.; Legeros, R.Z.; Nery, E.; Lynch, K.; Kerebel, B. Transformation of biphasic calcium phosphate ceramics in vivo: Ultrastructural and physicochemical characterization. J. Biomed. Mater. Res. 1989, 23, 883–894. [Google Scholar] [CrossRef]
- Chow, L.C.; Takagi, S. A Natural Bone Cement - A Laboratory Novelty Led to the Development of Revolutionary New Biomaterials. J. Res. Natl. Inst. Stand. Technol. 2001, 106, 1029–1033. [Google Scholar] [CrossRef]
- Xu, H.H.K.; Weir, M.D.; Burguera, E.F.; Fraser, A.M. Injectable and macroporous calcium phosphate cement scaffold. Biomaterials 2006, 27, 4279–4287. [Google Scholar] [CrossRef]
- Weir, M.D.; Xu, H.H.K. Osteoblastic induction on calcium phosphate cement-chitosan constructs for bone tissue engineering. J. Biomed. Mater. Res. Part A 2010, 94, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-S.; Park, M.S.; Gwak, S.-J.; Choi, C.Y.; Kim, B.-S. Accelerated bonelike apatite growth on porous polymer/ceramic composite scaffolds in vitro. Tissue Eng. 2006, 12, 2997–3006. [Google Scholar] [CrossRef]
- Jeon, O.H.; Panicker, L.M.; Lu, Q.; Chae, J.J.; Feldman, R.A.; Elisseeff, J.H. Human iPSC-derived osteoblasts and osteoclasts together promote bone regeneration in 3D biomaterials. Sci. Rep. 2016, 6, 26761. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-S.; Park, M.S.; Jeon, O.; Choi, C.Y.; Kim, B.-S. Poly(lactide-co-glycolide)/hydroxyapatite composite scaffolds for bone tissue engineering. Biomaterials 2006, 27, 1399–1409. [Google Scholar] [CrossRef]
- Wang, P.; Song, Y.; Weir, M.D.; Sun, J.; Zhao, L.; Simon, C.G.; Xu, H.H.K. A self-setting iPSMSC-alginate-calcium phosphate paste for bone tissue engineering. Dent. Mater. 2016, 32, 252–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hench, L.L.; Splinter, R.J.; Allen, W.C.; Greenlee, T.K. Bonding mechanisms at the interface of ceramic prosthetic materials. J. Biomed. Mater. Res. 1971, 5, 117–141. [Google Scholar] [CrossRef]
- Moimas, L.; Biasotto, M.; Di Lenarda, R.; Olivo, A.; Schmid, C. Rabbit pilot study on the resorbability of three-dimensional bioactive glass fibre scaffolds. Acta Biomater. 2006, 2, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Allan, I.; Newman, H.; Wilson, M. Antibacterial activity of particulate Bioglass® against supra- and subgingival bacteria. Biomaterials 2001, 22, 1683–1687. [Google Scholar] [CrossRef]
- Stoor, P.; Söderling, E.; Salonen, J.I. Antibacterial effects of a bioactive glass paste on oral microorganisms. Acta Odontol. Scand. 1998, 56, 161–165. [Google Scholar] [CrossRef]
- Coraça-Huber, D.C.; Fille, M.; Hausdorfer, J.; Putzer, D.; Nogler, M. Efficacy of antibacterial bioactive glass S53P4 against S. aureus biofilms grown on titanium discs in vitro. J. Orthop. Res. 2014, 32, 175–177. [Google Scholar] [CrossRef]
- Drago, L.; Romanò, D.; De Vecchi, E.; Vassena, C.; Logoluso, N.; Mattina, R.; Romanò, C.L. Bioactive glass BAG-S53P4 for the adjunctive treatment of chronic osteomyelitis of the long bones: An in vitro and prospective clinical study. BMC Infect. Dis. 2013, 13, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aurégan, J.-C. Bioactive glass for long bone infection: A systematic review. Injury 2015, 46, S3–S7. [Google Scholar] [CrossRef]
- Baino, F.; Hamzehlou, S.; Kargozar, S. Bioactive glasses: Where are we and where are we going? J. Funct. Biomater. 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, A.; Veis, A. Phosphorylated proteins and control over apatite nucleation, crystal growth, and inhibition. Chem. Rev. 2008, 108, 4670–4693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, W.J.; Silver, F.H. Mineral deposition in the extracellular matrices of vertebrate tissues: Identification of possible apatite nucleation sites on type I collagen. Cells. Tissues. Organs 2009, 189, 20–24. [Google Scholar] [CrossRef]
- Nudelman, F.; Pieterse, K.; George, A.; Bomans, P.H.H.; Friedrich, H.; Brylka, L.J.; Hilbers, P.A.J.; de With, G.; Sommerdijk, N.A.J.M. The role of collagen in bone apatite formation in the presence of hydroxyapatite nucleation inhibitors. Nat. Mater. 2010, 9, 1004–1009. [Google Scholar] [CrossRef] [Green Version]
- Itoh, S.; Kikuchi, M.; Koyama, Y.; Takakuda, K.; Shinomiya, K.; Tanaka, J. Development of a hydroxyapatite/collagen nanocomposite as a medical device. Cell Transplant. 2004, 13, 451–462. [Google Scholar] [CrossRef]
- Walsh, W.R.; Oliver, R.A.; Christou, C.; Lovric, V.; Walsh, E.R.; Prado, G.R.; Haider, T. Critical size bone defect healing using collagen-calcium phosphate bone graft materials. PLoS ONE 2017, 12, e0168883. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Cheung, T.F.; Fan, V.C.; Sin, K.M.; Wong, C.W.Y.; Leung, G.K.K. Applications of three-dimensional printing in surgery. Surg. Innov. 2017, 24, 82–88. [Google Scholar] [CrossRef]
- Bauermeister, A.J.; Zuriarrain, A.; Newman, M.I. Three-dimensional printing in plastic and reconstructive surgery. Ann. Plast. Surg. 2016, 77, 569–576. [Google Scholar] [CrossRef]
- Fang, C.; Cai, H.; Kuong, E.; Chui, E.; Siu, Y.C.; Ji, T.; Drstvenšek, I. Surgical applications of three-dimensional printing in the pelvis and acetabulum: From models and tools to implants. Unfallchirurg 2019, 122, 278–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guvendiren, M.; Molde, J.; Soares, R.M.D.; Kohn, J. Designing biomaterials for 3D printing. ACS Biomater. Sci. Eng. 2016, 2, 1679–1693. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, R.; Inzana, J.A.; Schwarz, E.M.; Kates, S.L.; Awad, H.A. 3D printing of calcium phosphate ceramics for bone tissue engineering and drug delivery. Ann. Biomed. Eng. 2017, 45, 23–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anitha, A.; Menon, D.; Sivanarayanan, T.B.; Koyakutty, M.; Mohan, C.C.; Nair, S.V.; Nair, M.B. Bioinspired composite matrix containing hydroxyapatite–silica core–shell nanorods for bone tissue engineering. ACS Appl. Mater. Interfaces 2017, 9, 26707–26718. [Google Scholar]
- Turner, R.J.; Renshaw, J.C.; Hamilton, A. Biogenic hydroxyapatite: A new material for the preservation and restoration of the built environment. ACS Appl. Mater. Interfaces 2017, 9, 31401–31410. [Google Scholar] [CrossRef] [Green Version]
- Richards, D.J.; Coyle, R.C.; Tan, Y.; Jia, J.; Wong, K.; Toomer, K.; Menick, D.R.; Mei, Y. Inspiration from heart development: Biomimetic development of functional human cardiac organoids. Biomaterials 2017, 142, 112–123. [Google Scholar] [CrossRef]
- Löffler, S.; Seyock, S.; Nybom, R.; Jacobson, G.B.; Richter-Dahlfors, A. Electrochemically triggered release of acetylcholine from scCO2 impregnated conductive polymer films evokes intracellular Ca2+ signaling in neurotypic SH-SY5Y cells. J. Control. Release 2016, 243, 283–290. [Google Scholar] [CrossRef]
- Wang, T.; Wang, D.; Liu, J.; Feng, B.; Zhou, F.; Zhang, H.; Zhou, L.; Yin, Q.; Zhang, Z.; Cao, Z.; et al. Acidity-triggered ligand-presenting nanoparticles to overcome sequential drug delivery barriers to tumors. Nano Lett. 2017, 17, 5429–5436. [Google Scholar] [CrossRef]
- Hang, C.; Zou, Y.; Zhong, Y.; Zhong, Z.; Meng, F. NIR and UV-responsive degradable hyaluronic acid nanogels for CD44-targeted and remotely triggered intracellular doxorubicin delivery. Colloids Surf. B Biointerfaces 2017, 158, 547–555. [Google Scholar] [CrossRef]
- He, M.; Wang, Q.; Shi, Z.; Xie, Y.; Zhao, W.; Zhao, C. Inflammation-responsive self-regulated drug release from ultrathin hydrogel coating. Colloids Surf. B Biointerfaces 2017, 158, 518–526. [Google Scholar] [CrossRef]
- Minkowitz, R.B. Removal of painful orthopaedic implants after fracture union. J. Bone Jt. Surg. 2007, 89, 1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Witte, F.; Lu, F.; Wang, J.; Li, J.; Qin, L. Current status on clinical applications of magnesium-based orthopaedic implants: A review from clinical translational perspective. Biomaterials 2017, 112, 287–302. [Google Scholar] [CrossRef] [PubMed]





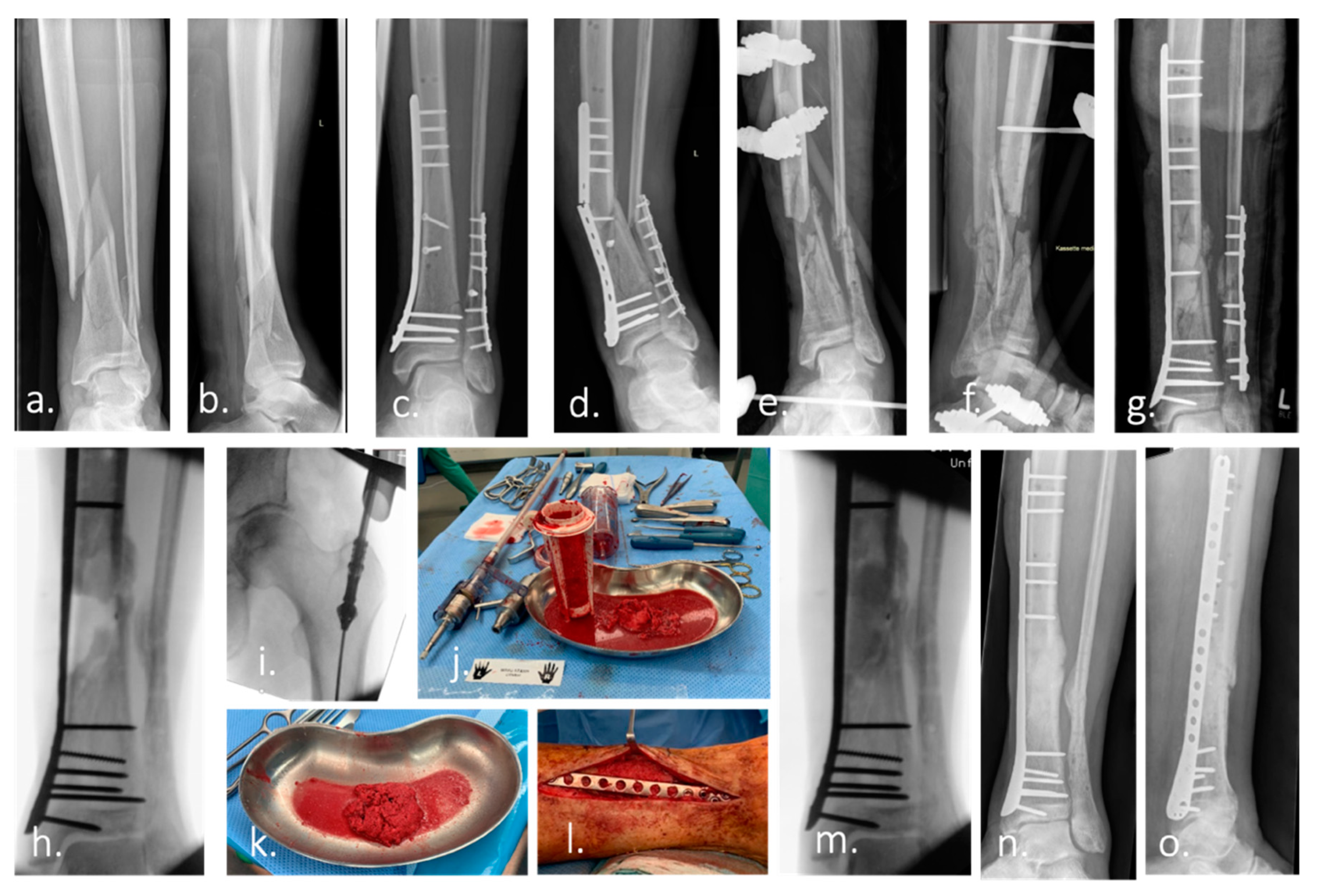{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient-Related Cause | Patient-Unrelated Cause |
|---|---|
| Age Gender (Male) | Open fracture (initial bone loss and soft tissue injury) Displaced fracture |
| Smoking 1 Alcohol 1 | Wedge and multi-fragmentary fracture pattern Injury location (tibia) |
| High Body mass index 1 Malnutrition 1 (Calcium, Vitamin D, etc.) | Soft tissue damage - vascularisation 2 Open reduction - quality of primary osteosynthesis 2 |
| Osteoporosis 2 Diabetes 2 | Compartment syndrome 2 Presence of fracture gap post-surgery 2 |
| Peripheral vascular disease 2 Chronic inflammatory disease 2 | Poor mechanical stability by initial implant 2 |
| Medication 2 (Insulin, Chemotherapeutics, Steroids, Antibiotics, Anticoagulants, NSAIDs, Opiates) | |
| Renal insufficiency Radio therapy |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlickewei, C.W.; Kleinertz, H.; Thiesen, D.M.; Mader, K.; Priemel, M.; Frosch, K.-H.; Keller, J. Current and Future Concepts for the Treatment of Impaired Fracture Healing. Int. J. Mol. Sci. 2019, 20, 5805. https://doi.org/10.3390/ijms20225805
Schlickewei CW, Kleinertz H, Thiesen DM, Mader K, Priemel M, Frosch K-H, Keller J. Current and Future Concepts for the Treatment of Impaired Fracture Healing. International Journal of Molecular Sciences. 2019; 20(22):5805. https://doi.org/10.3390/ijms20225805
Chicago/Turabian StyleSchlickewei, Carsten W., Holger Kleinertz, Darius M. Thiesen, Konrad Mader, Matthias Priemel, Karl-Heinz Frosch, and Johannes Keller. 2019. "Current and Future Concepts for the Treatment of Impaired Fracture Healing" International Journal of Molecular Sciences 20, no. 22: 5805. https://doi.org/10.3390/ijms20225805
APA StyleSchlickewei, C. W., Kleinertz, H., Thiesen, D. M., Mader, K., Priemel, M., Frosch, K. -H., & Keller, J. (2019). Current and Future Concepts for the Treatment of Impaired Fracture Healing. International Journal of Molecular Sciences, 20(22), 5805. https://doi.org/10.3390/ijms20225805