The Ecology of Salicylic Acid Signaling: Primary, Secondary and Tertiary Effects with Applications in Agriculture

Abstract
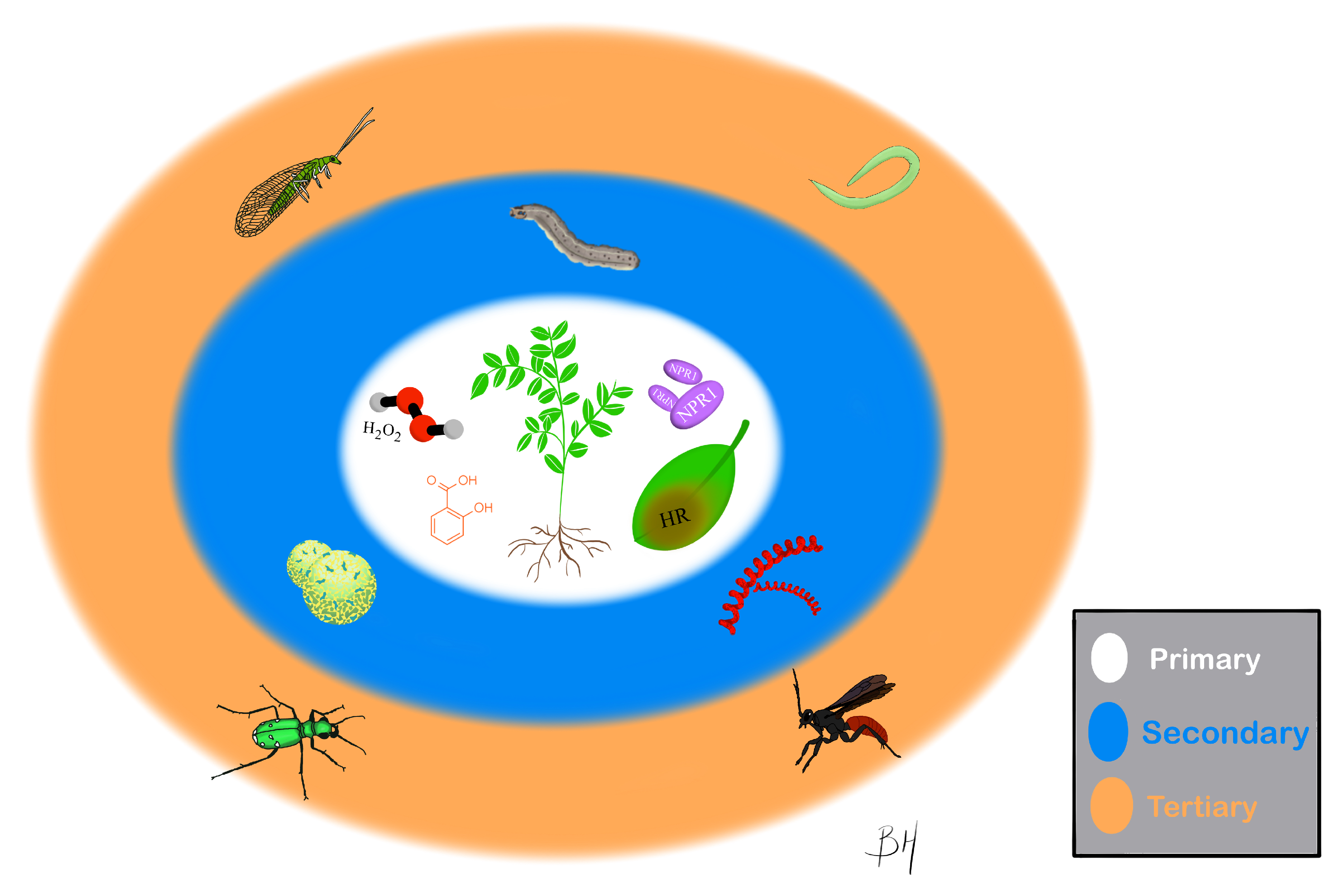
:1. Introduction
2. Primary Effects of SA
2.1. Production and Processing
2.2. Plant Response
2.3. Induction
3. Secondary Effects of SA
3.1. Direct Effects
3.2. Interactions
3.3. Temporal Considerations
4. Tertiary Effects of SA
4.1. Aboveground Natural Enemies
4.2. Belowground Natural Enemies
4.3. Connecting Aboveground and Belowground with SA
5. Applications of SA Induction for Control of Pests and Pathogens
5.1. Exogenous Induction
5.2. Genetic Approaches to Using SA in Agriculture
5.3. Natural Enemy Attraction
5.4. Opportunities
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SA | Salicylic Acid |
| JA | Jasmonic Acid |
| PAL | Phenylalanine ammonia lysase |
| ICS | Isochorismate synthase |
| MeSA | Methyl Salicylate |
| SAR | Systemic Acquired Resistance |
| PTI | Pattern Triggered Immunity |
| PR | Pathogenesis related |
| NPR | Non-expressor of PR |
| ETI | Effector Triggered Immunity |
| BTH | Benzo(1,2,3)thiadiazole-7-carbothioic acid S-methyl ester |
References
- Bruinsma, J. World Agriculture: Towards 2015/2030: An FAO Study; Routledge: London, UK, 2017. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.K.; Ramankutty, N.; Mueller, N.D.; West, P.C.; Foley, J.A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 2012, 3, 1293. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Roberts, M.J.; Schlenker, W.; Braun, N.; Little, B.B.; Rejesus, R.M.; Hammer, G.L. Greater sensitivity to drought accompanies maize yield increase in the US Midwest. Science 2014, 344, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Moreira, X.; Abdala-Roberts, L.; Gols, R.; Francisco, M. Plant domestication decreases both constitutive and induced chemical defences by direct selection against defensive traits. Sci. Rep. 2018, 8, 12678. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Lin, P.A.; Foolad, M.R.; Ali, J.G.; Rajotte, E.G.; Felton, G.W. Induced Plant Defenses Against Herbivory in Cultivated and Wild Tomato. J. Chem. Ecol. 2019, 45, 693–707. [Google Scholar] [CrossRef]
- Whitehead, S.R.; Turcotte, M.M.; Poveda, K. Domestication impacts on plant–herbivore interactions: A meta-analysis. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160034. [Google Scholar] [CrossRef]
- Chen, Y.H.; Gols, R.; Stratton, C.A.; Brevik, K.A.; Benrey, B. Complex tritrophic interactions in response to crop domestication: Predictions from the wild. Entomol. Exp. Appl. 2015, 157, 40–59. [Google Scholar] [CrossRef]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef]
- Vallad, G.E.; Goodman, R.M. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Sci. 2004, 44, 1920–1934. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.; Kaplan, I.; Braasch, J.; Chinnasamy, D.; Williams, L. Field responses of predaceous arthropods to methyl salicylate: A meta-analysis and case study in cranberries. Biol. Control 2011, 59, 294–303. [Google Scholar] [CrossRef]
- Koornneef, A.; Pieterse, C.M. Cross talk in defense signaling. Plant Physiol. 2008, 146, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2002, 5, 325–331. [Google Scholar] [CrossRef]
- Vos, I.A.; Moritz, L.; Pieterse, C.M.; Van Wees, S. Impact of hormonal crosstalk on plant resistance and fitness under multi-attacker conditions. Front. Plant Sci. 2015, 6, 639. [Google Scholar] [CrossRef]
- Caarls, L.; Pieterse, C.M.; Van Wees, S. How salicylic acid takes transcriptional control over jasmonic acid signaling. Front. Plant Sci. 2015, 6, 170. [Google Scholar] [CrossRef]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef]
- Mahdi, J.; Mahdi, A.; Mahdi, A.; Bowen, I. The historical analysis of aspirin discovery, its relation to the willow tree and antiproliferative and anticancer potential. Cell Prolif. 2006, 39, 147–155. [Google Scholar] [CrossRef]
- Kumar, D.; Haq, I.; Chapagai, D.; Tripathi, D.; Donald, D.; Hossain, M.; Devaiah, S. Hormone signaling: Current perspectives on the roles of salicylic acid and its derivatives in plants. In The Formation, Structure and Activity of Phytochemicals; Springer: Berlin/Heidelberg, Germany, 2015; pp. 115–136. [Google Scholar]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Heil, M.; Ton, J. Long-distance signalling in plant defence. Trends Plant Sci. 2008, 13, 264–272. [Google Scholar] [CrossRef]
- Shah, J. Plants under attack: Systemic signals in defence. Curr. Opin. Plant Biol. 2009, 12, 459–464. [Google Scholar] [CrossRef]
- Shulaev, V.; Silverman, P.; Raskin, I. Airborne signalling by methyl salicylate in plant pathogen resistance. Nature 1997, 385, 718. [Google Scholar] [CrossRef]
- D’Maris Amick Dempsey, A.C.; Vlot, M.C.W.; Daniel, F.K. Salicylic acid biosynthesis and metabolism. Arab. Book Am. Soc. Plant Biol. 2011, 9, e0156. [Google Scholar]
- Chen, Z.; Zheng, Z.; Huang, J.; Lai, Z.; Fan, B. Biosynthesis of salicylic acid in plants. Plant Signal. Behav. 2009, 4, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, D.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Chapagai, D.; Dean, P.; Davenport, M. Biotic and abiotic stress signaling mediated by salicylic acid. In Elucidation of Abiotic Stress Signaling in Plants; Springer: Berlin/Heidelberg, Germany, 2015; pp. 329–346. [Google Scholar]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef]
- Raskin, I.; Skubatz, H.; Tang, W.; Meeuse, B.J. Salicylic acid levels in thermogenic and non-thermogenic plants. Ann. Bot. 1990, 66, 369–373. [Google Scholar] [CrossRef]
- Chen, Z.; Iyer, S.; Caplan, A.; Klessig, D.F.; Fan, B. Differential accumulation of salicylic acid and salicylic acid-sensitive catalase in different rice tissues. Plant Physiol. 1997, 114, 193–201. [Google Scholar] [CrossRef]
- Raskin, I.; Ehmann, A.; Melander, W.R.; Meeuse, B.J. Salicylic acid: A natural inducer of heat production in Arum lilies. Science 1987, 237, 1601–1602. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Manthe, B.; Schulz, M.; Schnabl, H. Effects of salicylic acid on growth and stomatal movements ofVicia faba L.: Evidence for salicylic acid metabolization. J. Chem. Ecol. 1992, 18, 1525–1539. [Google Scholar] [CrossRef]
- Rate, D.N.; Cuenca, J.V.; Bowman, G.R.; Guttman, D.S.; Greenberg, J.T. The gain-of-function Arabidopsis acd6 mutant reveals novel regulation and function of the salicylic acid signaling pathway in controlling cell death, defenses, and cell growth. Plant Cell 1999, 11, 1695–1708. [Google Scholar] [CrossRef] [PubMed]
- Janda, T.; Gondor, O.K.; Yordanova, R.; Szalai, G.; Pál, M. Salicylic acid and photosynthesis: Signalling and effects. Acta Physiol. Plant. 2014, 36, 2537–2546. [Google Scholar] [CrossRef] [Green Version]
- Rajjou, L.; Belghazi, M.; Huguet, R.; Robin, C.; Moreau, A.; Job, C.; Job, D. Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination and establishment of early defense mechanisms. Plant Physiol. 2006, 141, 910–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.A. Systemic acquired resistance and salicylic acid: Past, present, and future. Mol. Plant-Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanacker, H.; Lu, H.; Rate, D.N.; Greenberg, J.T. A role for salicylic acid and NPR1 in regulating cell growth in Arabidopsis. Plant J. 2001, 28, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Kadioglu, A.; Saruhan, N.; Sağlam, A.; Terzi, R.; Acet, T. Exogenous salicylic acid alleviates effects of long term drought stress and delays leaf rolling by inducing antioxidant system. Plant Growth Regul. 2011, 64, 27–37. [Google Scholar] [CrossRef]
- Lu, H.; Greenberg, J.T.; Holuigue, L. Salicylic acid signaling networks. Front. Plant Sci. 2016, 7, 238. [Google Scholar] [CrossRef] [Green Version]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef]
- Maruri-López, I.; Aviles-Baltazar, N.Y.; Buchala, A.; Serrano, M. Intra and extracellular journey of the phytohormone salicylic acid. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Heath, M.C. Hypersensitive response-related death. In Programmed Cell Death in Higher Plants; Springer: Berlin/Heidelberg, Germany, 2000; pp. 77–90. [Google Scholar]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Innes, R. The positives and negatives of NPR: A unifying model for salicylic acid signaling in plants. Cell 2018, 173, 1314–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudisha, J.; Sharathchandra, R.; Amruthesh, K.; Kumar, A.; Shetty, H.S. Pathogenesis related proteins in plant defense response. In Plant Defence: Biological Control; Springer: Berlin/Heidelberg, Germany, 2012; pp. 379–403. [Google Scholar]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Shah, L.; Rahman, S.; Riaz, M.W.; Yahya, M.; Xu, Y.J.; Liu, F.; Si, W.; Jiang, H.; Cheng, B. Plant defense mechanism and current understanding of salicylic acid and NPRs in activating SAR. Physiol. Mol. Plant Pathol. 2018, 104, 15–22. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Klessig, D.F. SOS–too many signals for systemic acquired resistance? Trends Plant Sci. 2012, 17, 538–545. [Google Scholar] [CrossRef]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Bernsdorff, F.; Döring, A.C.; Gruner, K.; Schuck, S.; Bräutigam, A.; Zeier, J. Pipecolic acid orchestrates plant systemic acquired resistance and defense priming via salicylic acid-dependent and-independent pathways. Plant Cell 2016, 28, 102–129. [Google Scholar] [CrossRef] [Green Version]
- Shah, J.; Chaturvedi, R.; Chowdhury, Z.; Venables, B.; Petros, R.A. Signaling by small metabolites in systemic acquired resistance. Plant J. 2014, 79, 645–658. [Google Scholar] [CrossRef]
- Hartmann, M.; Zeier, J. N-Hydroxypipecolic acid and salicylic acid: A metabolic duo for systemic acquired resistance. Curr. Opin. Plant Biol. 2019, 50, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Doherty, H.M.; Selvendran, R.R.; Bowles, D.J. The wound response of tomato plants can be inhibited by aspirin and related hydroxy-benzoic acids. Physiol. Mol. Plant Pathol. 1988, 33, 377–384. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Front. Plant Sci. 2015, 5, 804. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Basra, S.; Wahid, A.; Ahmad, N.; Saleem, B. Improving the drought tolerance in rice (Oryza sativa L.) by exogenous application of salicylic acid. J. Agron. Crop Sci. 2009, 195, 237–246. [Google Scholar] [CrossRef]
- Yang, C.; Hu, L.; Ali, B.; Islam, F.; Bai, Q.; Yun, X.; Yoneyama, K.; Zhou, W. Seed treatment with salicylic acid invokes defence mechanism of Helianthus annuus against Orobanche cumana. Ann. Appl. Biol. 2016, 169, 408–422. [Google Scholar] [CrossRef]
- Ansari, A.; Razmjoo, J.; Karimmojeni, H. Mycorrhizal colonization and seed treatment with salicylic acid to improve physiological traits and tolerance of flaxseed (Linum usitatissimum L.) plants grown under drought stress. Acta Physiol. Plant. 2016, 38, 34. [Google Scholar] [CrossRef]
- Smith, J.L.; De Moraes, C.M.; Mescher, M.C. Jasmonate-and salicylate-mediated plant defense responses to insect herbivores, pathogens and parasitic plants. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 497–503. [Google Scholar] [CrossRef]
- Mohamed, A.; Ellicott, A.; Housley, T.; Ejeta, G. Hypersensitive response to Striga infection in sorghum. Crop Sci. 2003, 43, 1320–1324. [Google Scholar] [CrossRef]
- Borsics, T.; Lados, M. Dodder infection induces the expression of a pathogenesis-related gene of the family PR-10 in alfalfa. J. Exp. Bot. 2002, 53, 1831–1832. [Google Scholar] [CrossRef] [Green Version]
- Runyon, J.B.; Mescher, M.C.; Felton, G.W.; De Moraes, C.M. Parasitism by Cuscuta pentagona sequentially induces JA and SA defence pathways in tomato. Plant Cell Environ. 2010, 33, 290–303. [Google Scholar] [CrossRef]
- Moran, P.J.; Thompson, G.A. Molecular responses to aphid feeding in Arabidopsis in relation to plant defense pathways. Plant Physiol. 2001, 125, 1074–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohase, L.; van der Westhuizen, A.J. Salicylic acid is involved in resistance responses in the Russian wheat aphid-wheat interaction. J. Plant Physiol. 2002, 159, 585–590. [Google Scholar] [CrossRef]
- Li, Q.; Xie, Q.G.; Smith-Becker, J.; Navarre, D.A.; Kaloshian, I. Mi-1-mediated aphid resistance involves salicylic acid and mitogen-activated protein kinase signaling cascades. Mol. Plant-Microbe Interact. 2006, 19, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarate, S.I.; Kempema, L.A.; Walling, L.L. Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Deng, X.; Jia, S.; Huang, J.; Miao, X.; Huang, Y. Role of salicylic acid in tomato defense against cotton bollworm, Helicoverpa armigera Hubner. Z. Für Naturforschung 2004, 59, 856–862. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [Green Version]
- Broekgaarden, C.; Caarls, L.; Vos, I.A.; Pieterse, C.M.; Van Wees, S.C. Ethylene: Traffic controller on hormonal crossroads to defense. Plant Physiol. 2015, 169, 2371–2379. [Google Scholar] [CrossRef] [Green Version]
- Karapetyan, S.; Dong, X. Redox and the circadian clock in plant immunity: A balancing act. Free Radic. Biol. Med. 2018, 119, 56–61. [Google Scholar] [CrossRef]
- Suárez-Vidal, E.; López-Goldar, X.; Sampedro, L.; Zas, R. Effect of light availability on the interaction between maritime pine and the pine weevil: Light drives insect feeding behavior but also the defensive capabilities of the host. Front. Plant Sci. 2017, 8, 1452. [Google Scholar] [CrossRef] [Green Version]
- Poór, P.; Takács, Z.; Bela, K.; Czékus, Z.; Szalai, G.; Tari, I. Prolonged dark period modulates the oxidative burst and enzymatic antioxidant systems in the leaves of salicylic acid-treated tomato. J. Plant Physiol. 2017, 213, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Stout, M.J.; Thaler, J.S.; Thomma, B.P. Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods. Annu. Rev. Entomol. 2006, 51, 663–689. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Shah, J. Salicylic acid in plant disease resistance. In Salicylic Acid: A Plant Hormone; Springer: Berlin/Heidelberg, Germany, 2007; pp. 335–370. [Google Scholar]
- Yang, L.; Li, B.; Zheng, X.y.; Li, J.; Yang, M.; Dong, X.; He, G.; An, C.; Deng, X.W. Salicylic acid biosynthesis is enhanced and contributes to increased biotrophic pathogen resistance in Arabidopsis hybrids. Nat. Commun. 2015, 6, 7309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coquoz, J.; Buchala, A.; Meuwly, P.; Metraux, J. Arachidonic acid induces local but not systemic synthesis of salicylic acid and confers systemic resistance in potato plants to Phytophthora infestans and Alternaria solani. Phytopathology 1995, 85, 1219–1224. [Google Scholar] [CrossRef]
- Silverman, P.; Seskar, M.; Kanter, D.; Schweizer, P.; Metraux, J.P.; Raskin, I. Salicylic acid in rice (biosynthesis, conjugation, and possible role). Plant Physiol. 1995, 108, 633–639. [Google Scholar] [CrossRef] [Green Version]
- Halim, V.A.; Eschen-Lippold, L.; Altmann, S.; Birschwilks, M.; Scheel, D.; Rosahl, S. Salicylic acid is important for basal defense of Solanum tuberosum against Phytophthora infestans. Mol. Plant Microbe Interact. 2007, 20, 1346–1352. [Google Scholar] [CrossRef] [Green Version]
- Hein, J.W.; Wolfe, G.V.; Blee, K.A. Comparison of rhizosphere bacterial communities in Arabidopsis thaliana mutants for systemic acquired resistance. Microb. Ecol. 2008, 55, 333–343. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; Von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “talking trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, I.T.; Kessler, A.; Halitschke, R. Volatile signaling in plant–plant–herbivore interactions: What is ewblock? Curr. Opin. Plant Biol. 2002, 5, 351–354. [Google Scholar] [CrossRef]
- Pearse, I.S.; Hughes, K.; Shiojiri, K.; Ishizaki, S.; Karban, R. Interplant volatile signaling in willows: Revisiting the original talking trees. Oecologia 2013, 172, 869–875. [Google Scholar] [CrossRef]
- Karban, R.; Huntzinger, M.; McCall, A.C. The specificity of eavesdropping on sagebrush by other plants. Ecology 2004, 85, 1846–1852. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Blande, J.D. Molecular plant volatile communication. In Sensing in Nature; Springer: Berlin/Heidelberg, Germany, 2012; pp. 17–31. [Google Scholar]
- You, L.X.; Wang, P.; Kong, C.H. The levels of jasmonic acid and salicylic acid in a rice-barnyardgrass coexistence system and their relation to rice allelochemicals. Biochem. Syst. Ecol. 2011, 39, 491–497. [Google Scholar] [CrossRef]
- Schreiner, M.; Krumbein, A.; Knorr, D.; Smetanska, I. Enhanced glucosinolates in root exudates of Brassica rapa ssp. rapa mediated by salicylic acid and methyl jasmonate. J. Agric. Food Chem. 2011, 59, 1400–1405. [Google Scholar] [CrossRef]
- Kong, C.H.; Zhang, S.Z.; Li, Y.H.; Xia, Z.C.; Yang, X.F.; Meiners, S.J.; Wang, P. Plant neighbor detection and allelochemical response are driven by root-secreted signaling chemicals. Nat. Commun. 2018, 9, 3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasteels, J.M.; Rowell-Rahier, M. The chemical ecology of herbivory on willows. Proc. R. Soc. 1992, 98, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Ruuhola, T. Dynamics of Salicylates in Willows and Its Relation to Herbivory; University of Joensuu: Joensuu, Finland, 2001. [Google Scholar]
- Ollerstam, O.; Larsson, S. Salicylic acid mediates resistance in the willow Salix viminalis against the gall midge Dasineura marginemtorquens. J. Chem. Ecol. 2003, 29, 163–174. [Google Scholar] [CrossRef]
- Lortzing, V.; Oberländer, J.; Lortzing, T.; Tohge, T.; Steppuhn, A.; Kunze, R.; Hilker, M. Insect egg deposition renders plant defence against hatching larvae more effective in a salicylic acid-dependent manner. Plant Cell Environ. 2019, 42, 1019–1032. [Google Scholar] [CrossRef]
- Bruessow, F.; Gouhier-Darimont, C.; Buchala, A.; Metraux, J.P.; Reymond, P. Insect eggs suppress plant defence against chewing herbivores. Plant J. 2010, 62, 876–885. [Google Scholar] [CrossRef] [Green Version]
- Groux, R.; Hilfiker, O.; Gouhier-Darimont, C.; Peñaflor, M.F.G.V.; Erb, M.; Reymond, P. Role of methyl salicylate on oviposition deterrence in Arabidopsis thaliana. J. Chem. Ecol. 2014, 40, 754–759. [Google Scholar] [CrossRef]
- Ulland, S.; Ian, E.; Mozuraitis, R.; Borg-Karlson, A.K.; Meadow, R.; Mustaparta, H. Methyl salicylate, identified as primary odorant of a specific receptor neuron type, inhibits oviposition by the moth Mamestra brassicae L. (Lepidoptera, Noctuidae). Chem. Senses 2007, 33, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Favaro, R.; Resende, J.T.V.; Gabriel, A.; Zeist, A.R.; Cordeiro, E.C.N.; Favaro Júnior, J.L. Salicylic acid: Resistance inducer to two-spotted spider mite in strawberry crop. Hortic. Bras. 2019, 37, 60–64. [Google Scholar] [CrossRef]
- Felton, G.; Korth, K.; Bi, J.; Wesley, S.; Huhman, D.; Mathews, M.; Murphy, J.; Lamb, C.; Dixon, R. Inverse relationship between systemic resistance of plants to microorganisms and to insect herbivory. Curr. Biol. 1999, 9, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Felton, G.W.; Korth, K.L. Trade-offs between pathogen and herbivore resistance. Curr. Opin. Plant Biol. 2000, 3, 309–314. [Google Scholar] [CrossRef]
- Stout, M.; Fidantsef, A.; Duffey, S.; Bostock, R. Signal interactions in pathogen and insect attack: Systemic plant-mediated interactions between pathogens and herbivores of the tomato, Lycopersicon esculentum. Physiol. Mol. Plant Pathol. 1999, 54, 115–130. [Google Scholar] [CrossRef]
- Johnson, S.N.; Clark, K.E.; Hartley, S.E.; Jones, T.H.; McKenzie, S.W.; Koricheva, J. Aboveground–belowground herbivore interactions: A meta-analysis. Ecology 2012, 93, 2208–2215. [Google Scholar] [CrossRef]
- Wang, M.; Biere, A.; Van der Putten, W.H.; Bezemer, T.M. Sequential effects of root and foliar herbivory on aboveground and belowground induced plant defense responses and insect performance. Oecologia 2014, 175, 187–198. [Google Scholar] [CrossRef]
- Erb, M.; Robert, C.A.; Hibbard, B.E.; Turlings, T.C. Sequence of arrival determines plant-mediated interactions between herbivores. J. Ecol. 2011, 99, 7–15. [Google Scholar] [CrossRef]
- Huang, W.; Robert, C.A.; Hervé, M.R.; Hu, L.; Bont, Z.; Erb, M. A mechanism for sequence specificity in plant-mediated interactions between herbivores. New Phytol. 2017, 214, 169–179. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant immunity: From signaling to epigenetic control of defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Espinas, N.A.; Saze, H.; Saijo, Y. Epigenetic control of defense signaling and priming in plants. Front. Plant Sci. 2016, 7, 1201. [Google Scholar] [CrossRef]
- Luna, E.; Bruce, T.J.; Roberts, M.R.; Flors, V.; Ton, J. Next-generation systemic acquired resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaughter, A.; Daniel, X.; Flors, V.; Luna, E.; Hohn, B.; Mauch-Mani, B. Descendants of primed Arabidopsis plants exhibit resistance to biotic stress. Plant Physiol. 2012, 158, 835–843. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Carrasco, G.; Martínez-Aguilar, K.; Alvarez-Venegas, R. Transgenerational defense priming for crop protection against plant pathogens: A hypothesis. Front. Plant Sci. 2017, 8, 696. [Google Scholar] [CrossRef] [Green Version]
- De Moraes, C.M.; Lewis, W.; Pare, P.; Alborn, H.; Tumlinson, J. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570. [Google Scholar] [CrossRef]
- Turlings, T.C.; Wäckers, F.L.; Vet, L.E.; Lewis, W.J.; Tumlinson, J.H. Learning of host-finding cues by hymenopterous parasitoids. In Insect Learning; Springer: Berlin/Heidelberg, Germany, 1993; pp. 51–78. [Google Scholar]
- Vet, L.E.; Lewis, W.J.; Carde, R.T. Parasitoid foraging and learning. In Chemical Ecology of Insects 2; Springer: Berlin/Heidelberg, Germany, 1995; pp. 65–101. [Google Scholar]
- Vlot, A.C.; Klessig, D.F.; Park, S.W. Systemic acquired resistance: The elusive signals. Curr. Opin. Plant Biol. 2008, 11, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; Sabelis, M.W.; Takabayashi, J.; Bruin, J.; Posthumus, M.A. Plant strategies of manipulating predatorprey interactions through allelochemicals: Prospects for application in pest control. J. Chem. Ecol. 1990, 16, 3091–3118. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, R.; Shimoda, T.; Kawaguchi, M.; Arimura, G.i.; Horiuchi, J.i.; Nishioka, T.; Takabayashi, J. Lotus japonicus infested with herbivorous mites emits volatile compounds that attract predatory mites. J. Plant Res. 2000, 113, 427–433. [Google Scholar] [CrossRef]
- Dicke, M.; Takabayashi, J.; Posthumus, M.A.; Schütte, C.; Krips, O.E. Plant—Phytoseiid interactions mediated by herbivore-induced plant volatiles: Variation in production of cues and in responses of predatory mites. Exp. Appl. Acarol. 1998, 22, 311–333. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Janssen, A.; Bruin, J.; Posthumus, M.A.; Sabelis, M.W. An ecological cost of plant defence: Attractiveness of bitter cucumber plants to natural enemies of herbivores. Ecol. Lett. 2002, 5, 377–385. [Google Scholar] [CrossRef]
- Geervliet, J.B.; Posthumus, M.A.; Vet, L.E.; Dicke, M. Comparative analysis of headspace volatiles from different caterpillar-infested or uninfested food plants of Pieris species. J. Chem. Ecol. 1997, 23, 2935–2954. [Google Scholar] [CrossRef]
- Scutareanu, P.; Drukker, B.; Bruin, J.; Posthumus, M.A.; Sabelis, M.W. Volatiles from Psylla-infested pear trees and their possible involvement in attraction of anthocorid predators. J. Chem. Ecol. 1997, 23, 2241–2260. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.; Pettersson, J.; Pickett, J.; Wadhams, L.; Woodcock, C. Spring migration of damson-hop aphid, Phorodon humuli (Homoptera, Aphididae), and summer host plant-derived semiochemicals released on feeding. J. Chem. Ecol. 1993, 19, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Glinwood, R.; Pettersson, J. Host choice and host leaving in Rhopalosiphum padi (Hemiptera: Aphididae) emigrants and repellency of aphid colonies on the winter host. Bull. Entomol. Res. 2000, 90, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Snoeren, T.A.; Mumm, R.; Poelman, E.H.; Yang, Y.; Pichersky, E.; Dicke, M. The herbivore-induced plant volatile methyl salicylate negatively affects attraction of the parasitoid Diadegma semiclausum. J. Chem. Ecol. 2010, 36, 479–489. [Google Scholar] [CrossRef] [Green Version]
- James, D.G. Field evaluation of herbivore-induced plant volatiles as attractants for beneficial insects: Methyl salicylate and the green lacewing, Chrysopa nigricornis. J. Chem. Ecol. 2003, 29, 1601–1609. [Google Scholar] [CrossRef]
- De Boer, J.G.; Dicke, M. The role of methyl salicylate in prey searching behavior of the predatory mite Phytoseiulus persimilis. J. Chem. Ecol. 2004, 30, 255–271. [Google Scholar] [CrossRef]
- Mallinger, R.E.; Hogg, D.B.; Gratton, C. Methyl salicylate attracts natural enemies and reduces populations of soybean aphids (Hemiptera: Aphididae) in soybean agroecosystems. J. Econ. Entomol. 2011, 104, 115–124. [Google Scholar] [CrossRef]
- James, D.G. Further field evaluation of synthetic herbivore-induced plan volatiles as attractants for beneficial insects. J. Chem. Ecol. 2005, 31, 481–495. [Google Scholar] [CrossRef]
- James, D.G.; Price, T.S. Field-testing of methyl salicylate for recruitment and retention of beneficial insects in grapes and hops. J. Chem. Ecol. 2004, 30, 1613–1628. [Google Scholar] [CrossRef]
- Lee, J.C. Effect of methyl salicylate-based lures on beneficial and pest arthropods in strawberry. Environ. Entomol. 2010, 39, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Filgueiras, C.C.; Willett, D.S.; Pereira, R.V.; Junior, A.M.; Pareja, M.; Duncan, L.W. Eliciting maize defense pathways aboveground attracts belowground biocontrol agents. Sci. Rep. 2016, 6, 36484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filgueiras, C.C.; Willett, D.S.; Junior, A.M.; Pareja, M.; El Borai, F.; Dickson, D.W.; Stelinski, L.L.; Duncan, L.W. Stimulation of the salicylic acid pathway aboveground recruits entomopathogenic nematodes belowground. PLoS ONE 2016, 11, e0154712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dam, N.M. Belowground herbivory and plant defenses. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 373–391. [Google Scholar] [CrossRef]
- Rasmann, S.; Hiltpold, I.; Ali, J. The role of root-produced volatile secondary metabolites in mediating soil interactions. In Advances in Selected Plant Physiology Aspects; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar]
- Van Dam, N.M.; Bouwmeester, H.J. Metabolomics in the rhizosphere: Tapping into belowground chemical communication. Trends Plant Sci. 2016, 21, 256–265. [Google Scholar] [CrossRef]
- Aratchige, N.; Lesna, I.; Sabelis, M. Below-ground plant parts emit herbivore-induced volatiles: Olfactory responses of a predatory mite to tulip bulbs infested by rust mites. Exp. Appl. Acarol. 2004, 33, 21–30. [Google Scholar] [CrossRef]
- Ali, J.G.; Alborn, H.T.; Stelinski, L.L. Subterranean herbivore-induced volatiles released by citrus roots upon feeding by Diaprepes abbreviatus recruit entomopathogenic nematodes. J. Chem. Ecol. 2010, 36, 361–368. [Google Scholar] [CrossRef]
- Van Tol, R.W.; Van Der Sommen, A.T.; Boff, M.I.; Van Bezooijen, J.; Sabelis, M.W.; Smits, P.H. Plants protect their roots by alerting the enemies of grubs. Ecol. Lett. 2001, 4, 292–294. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732. [Google Scholar] [CrossRef]
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Annu. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Ali, J.G.; Alborn, H.T.; Campos-Herrera, R.; Kaplan, F.; Duncan, L.W.; Rodriguez-Saona, C.; Koppenhöfer, A.M.; Stelinski, L.L. Subterranean, herbivore-induced plant volatile increases biological control activity of multiple beneficial nematode species in distinct habitats. PLoS ONE 2012, 7, e38146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenhardt, J.; Hiltpold, I.; Köllner, T.G.; Frey, M.; Gierl, A.; Gershenzon, J.; Hibbard, B.E.; Ellersieck, M.R.; Turlings, T.C. Restoring a maize root signal that attracts insect-killing nematodes to control a major pest. Proc. Natl. Acad. Sci. USA 2009, 106, 13213–13218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willett, D.S.; Alborn, H.T.; Stelinski, L.L. Multitrophic effects of belowground parasitoid learning. Sci. Rep. 2017, 7, 2067. [Google Scholar] [CrossRef] [PubMed]
- Filgueiras, C.C.; Willett, D.S.; Pereira, R.V.; Sabino, P.H.d.S.; Junior, A.M.; Pareja, M.; Dickson, D.W. Parameters affecting plant defense pathway mediated recruitment of entomopathogenic nematodes. Biocontrol Sci. Technol. 2017, 27, 833–843. [Google Scholar] [CrossRef]
- Neveu, N.; Grandgirard, J.; Nenon, J.; Cortesero, A. Systemic release of herbivore-induced plant volatiles by turnips infested by concealed root-feeding larvae Delia radicum L. J. Chem. Ecol. 2002, 28, 1717–1732. [Google Scholar] [CrossRef]
- Faize, L.; Faize, M. Functional analogues of salicylic acid and their use in crop protection. Agronomy 2018, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance-salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Song, G.C.; Ryu, C.M. Evidence for Volatile Memory in Plants: Boosting Defence Priming through the Recurrent Application of Plant Volatiles. Mol. Cells 2018, 41, 724. [Google Scholar]
- Wenig, M.; Ghirardo, A.; Sales, J.H.; Pabst, E.S.; Breitenbach, H.H.; Antritter, F.; Weber, B.; Lange, B.; Lenk, M.; Cameron, R.K.; et al. Systemic acquired resistance networks amplify airborne defense cues. Nat. Commun. 2019, 10, 3813. [Google Scholar] [CrossRef] [Green Version]
- Jaskiewicz, M.; Conrath, U.; Peterhänsel, C. Chromatin modification acts as a memory for systemic acquired resistance in the plant stress response. EMBO Rep. 2011, 12, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Ishii, H.; Tomita, Y.; Horio, T.; Narusaka, Y.; Nakazawa, Y.; Nishimura, K.; Iwamoto, S. Induced resistance of acibenzolar-S-methyl (CGA 245704) to cucumber and Japanese pear diseases. Eur. J. Plant Pathol. 1999, 105, 77–85. [Google Scholar] [CrossRef]
- Zhang, S.; Reddy, M.; Kokalis-Burelle, N.; Wells, L.W.; Nightengale, S.P.; Kloepper, J.W. Lack of induced systemic resistance in peanut to late leaf spot disease by plant growth-promoting rhizobacteria and chemical elicitors. Plant Dis. 2001, 85, 879–884. [Google Scholar] [CrossRef]
- Kouzai, Y.; Noutoshi, Y.; Inoue, K.; Shimizu, M.; Onda, Y.; Mochida, K. Benzothiadiazole, a plant defense inducer, negatively regulates sheath blight resistance in Brachypodium distachyon. Sci. Rep. 2018, 8, 17358. [Google Scholar] [CrossRef]
- Thaler, J.S.; Fidantsef, A.L.; Duffey, S.S.; Bostock, R.M. Trade-offs in plant defense against pathogens and herbivores: A field demonstration of chemical elicitors of induced resistance. J. Chem. Ecol. 1999, 25, 1597–1609. [Google Scholar] [CrossRef]
- Ning, Y.; Liu, W.; Wang, G.L. Balancing immunity and yield in crop plants. Trends Plant Sci. 2017, 22, 1069–1079. [Google Scholar] [CrossRef]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant domestication through an ecological lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Li, Y.; Huang, F.; Lu, Y.; Shi, Y.; Zhang, M.; Fan, J.; Wang, W. Mechanism of plant–microbe interaction and its utilization in disease-resistance breeding for modern agriculture. Physiol. Mol. Plant Pathol. 2013, 83, 51–58. [Google Scholar] [CrossRef]
- Xu, G.; Greene, G.H.; Yoo, H.; Liu, L.; Marqués, J.; Motley, J.; Dong, X. Global translational reprogramming is a fundamental layer of immune regulation in plants. Nature 2017, 545, 487. [Google Scholar] [CrossRef]
- Xu, G.; Yuan, M.; Ai, C.; Liu, L.; Zhuang, E.; Karapetyan, S.; Wang, S.; Dong, X. uORF-mediated translation allows engineered plant disease resistance without fitness costs. Nature 2017, 545, 491. [Google Scholar] [CrossRef]
- Hajek, A.E.; Eilenberg, J. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]


{kind=link}
| Plant | Herbivore | |||
|---|---|---|---|---|
| Family | Species | Common Name | Species | Citation |
| Fabacae | Phaseolus lunatus | Spider Mite | Tetranychus urticae Koch | [116] |
| Fabacae | Phaseolus lunatus | Spider Mite | Tetranychus urticae Koch | [117] |
| Solanaceae | Solanum lycopersicum | Spider Mite | Tetranychus urticae Koch | [118] |
| Cucurbitaceae | Cucumis sativus | Spider Mite | Tetranychus urticae Koch | [119] |
| Brassicaceae | Brassica oleracea capitata L. | Garden Whites | Pieris spp. | [120] |
| Rosaceae | Pyrus communis | Pear Psyllid | Psylla pyricola Forster | [121] |
| Cannabaceae | Humulus iupulus L. | Damson Hop Aphid | Phorodon humuli Forster | [122] |
| Rosaceae | Prunus padus | Bird Cherry–Oat Aphid | Rhopalosiphum padi L. | [123] |
| Solanaceae | Nicotiana attenuata Torr. ex Wats | Five-Spotted Hawkmoth | Manduca quinquemaculata | [124] |
| Solanaceae | Nicotiana attenuata Torr. ex Wats | Leaf Bug | Dicyphus minimus | [124] |
| Solanaceae | Nicotiana attenuata Torr. ex Wats | Tobacco Flea Beetle | Epitrix hirtipennis | [124] |
| Fabaceae | Lotus japonicus | Spider Mite | Tetranychus urticae | [117] |
| Plant | Herbivore | Natural Enemy | ||||||
|---|---|---|---|---|---|---|---|---|
| Family | Species | Common | Species | Common | Species | Impact on Herbivore | Setting | Citation |
| Cannabaceae | Humulus lupulus | Damson-Hop aphid | Phorodon humuli [122] | Green Lacewing | Chrysopa nigricornis | Not Reported | Field | [126] |
| Fabaceae | Phaseolus lunatus | Spider Mite | Tetranychus urticae | Phytoseiulus persimilis | Not Reported | Lab | [127] | |
| Fabaceae | Glycine max L. | Soybean Aphid | Aphis glycines | Green Lacewing | Chrysopa nigricornis | Reduced | Field/Lab | [128] |
| Fabaceae | Glycine max L. | Soybean Aphid | Aphis glycines | Syrphid Flies | Reduced | Field/Lab | [128] | |
| Cannabaceae | Humulus lupulus | Ladybeetle | Stethorus punctum picipes | Not Reported | Field | [129] | ||
| Cannabaceae | Humulus lupulus | Minute Pirate Bug | Orius tristicolor | Not Reported | Field | [129] | ||
| Cannabaceae | Humulus lupulus | Bigeyed Bug | Geocoris pallens | Not Reported | Field | [129] | ||
| Cannabaceae | Humulus lupulus | Syrphidae | Not Reported | Field | [129] | |||
| Cannabaceae | Humulus lupulus | Empididae | Not Reported | Field | [129] | |||
| Cannabaceae | Humulus lupulus | Sarcophagidae | Not Reported | Field | [129] | |||
| Cannabaceae | Humulus lupulus | Agromyzidae | Not Reported | Field | [129] | |||
| Cannabaceae | Humulus lupulus | Micro-hymenoptera | Not Reported | Field | [129] | |||
| Vitaceae | Vitis labrusca | Green Lacewing | Chrysopa nigricornis | Not Reported | Field | [130] | ||
| Vitaceae | Vitis labrusca | Lacewing | Hemerobius sp. | Not Reported | Field | [130] | ||
| Vitaceae | Vitis labrusca | Deraeocoris brevis | Not Reported | Field | [130] | |||
| Vitaceae | Vitis labrusca | Stethorus punctum picipes | Not Reported | Field | [130] | |||
| Vitaceae | Vitis labrusca | Minute Pirate Bug | Orius tristicolor | Not Reported | Field | [130] | ||
| Rosaceae | Fragaria sp. | Aphididae, Thripidae, Cicadellidae | Minute Pirate Bug | Orius tristicolor | No Effect 1 | Field | [131] | |
| Rosaceae | Fragaria sp. | Aphididae, Thripidae, Cicadellidae | Chrysopidae | No Effect 1 | Field | [131] | ||
| Poaceae | Zea mays | Cucurbit Beetle | Diabrotica speciosa | Entomopathogenic Nematode | Heterorhabditis amazonensis | Not Reported | Lab | [132] |
| Rutaceae | Citrus paradisi × Poncirus trifoliata | Citrus Root Weevil | Diaprepes abbreviatus | Entomopathogenic Nematode | Steinernema diaprepesi | Not Reported | Lab | [133] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filgueiras, C.C.; Martins, A.D.; Pereira, R.V.; Willett, D.S. The Ecology of Salicylic Acid Signaling: Primary, Secondary and Tertiary Effects with Applications in Agriculture. Int. J. Mol. Sci. 2019, 20, 5851. https://doi.org/10.3390/ijms20235851
Filgueiras CC, Martins AD, Pereira RV, Willett DS. The Ecology of Salicylic Acid Signaling: Primary, Secondary and Tertiary Effects with Applications in Agriculture. International Journal of Molecular Sciences. 2019; 20(23):5851. https://doi.org/10.3390/ijms20235851
Chicago/Turabian StyleFilgueiras, Camila C., Adalvan D. Martins, Ramom V. Pereira, and Denis S. Willett. 2019. "The Ecology of Salicylic Acid Signaling: Primary, Secondary and Tertiary Effects with Applications in Agriculture" International Journal of Molecular Sciences 20, no. 23: 5851. https://doi.org/10.3390/ijms20235851
APA StyleFilgueiras, C. C., Martins, A. D., Pereira, R. V., & Willett, D. S. (2019). The Ecology of Salicylic Acid Signaling: Primary, Secondary and Tertiary Effects with Applications in Agriculture. International Journal of Molecular Sciences, 20(23), 5851. https://doi.org/10.3390/ijms20235851