ABCC3 Expressed by CD56dim CD16+ NK Cells Predicts Response in Glioblastoma Patients Treated with Combined Chemotherapy and Dendritic Cell Immunotherapy

 ,
,  and
and
Abstract
:1. Introduction
2. Results
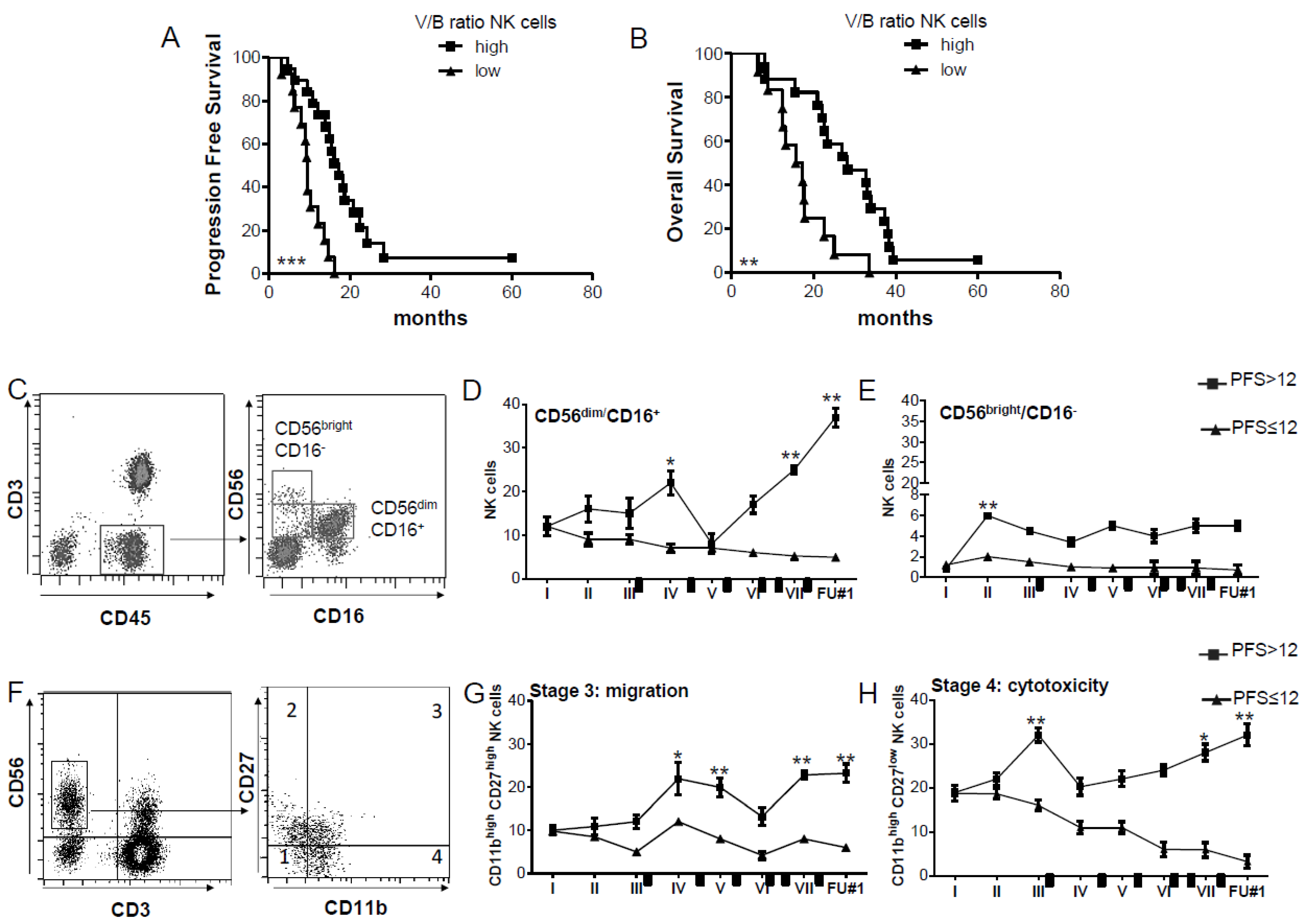
2.1. CD56dim CD16+ NK Cells with Cytolytic Ability Increase in DENDR1 PFS > 12 Patients
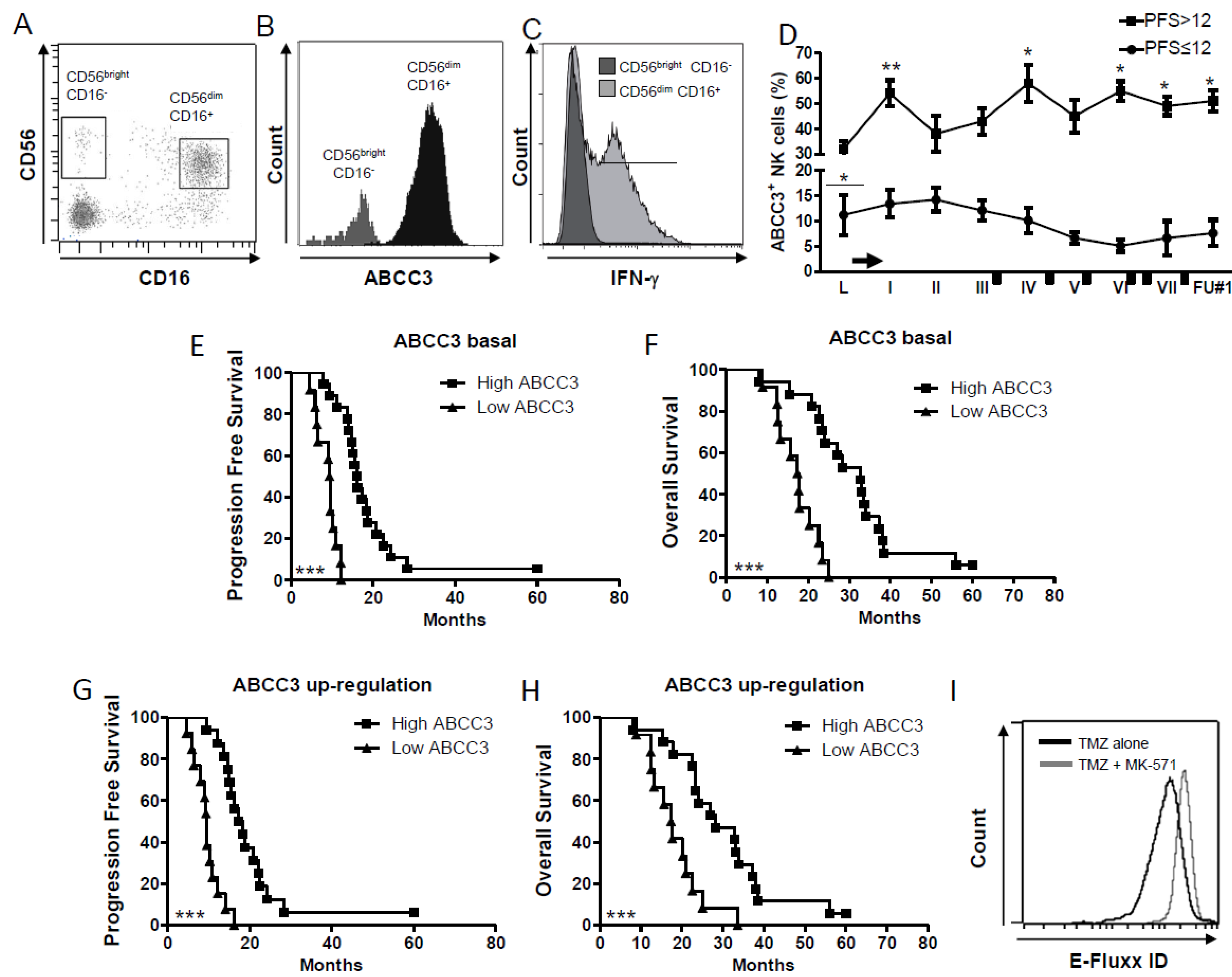
2.2. The Multidrug Resistance ABCC3 Expressed by CD56dim CD16+ NK Cells Is Active and Correlates with the Clinical Outcome
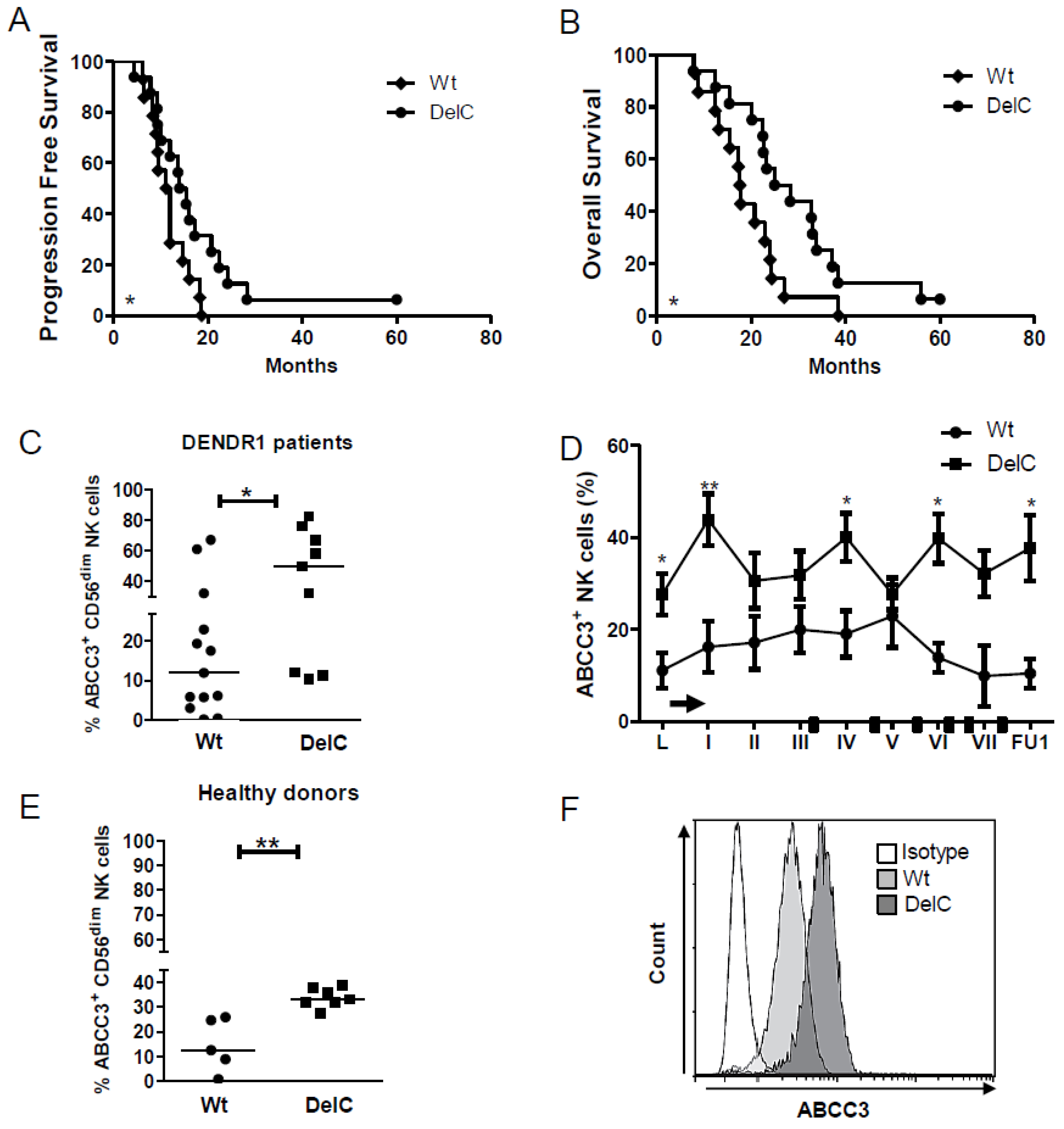
2.3. The ABCC3 SNP rs35467079 Correlates with A Prolonged Survival
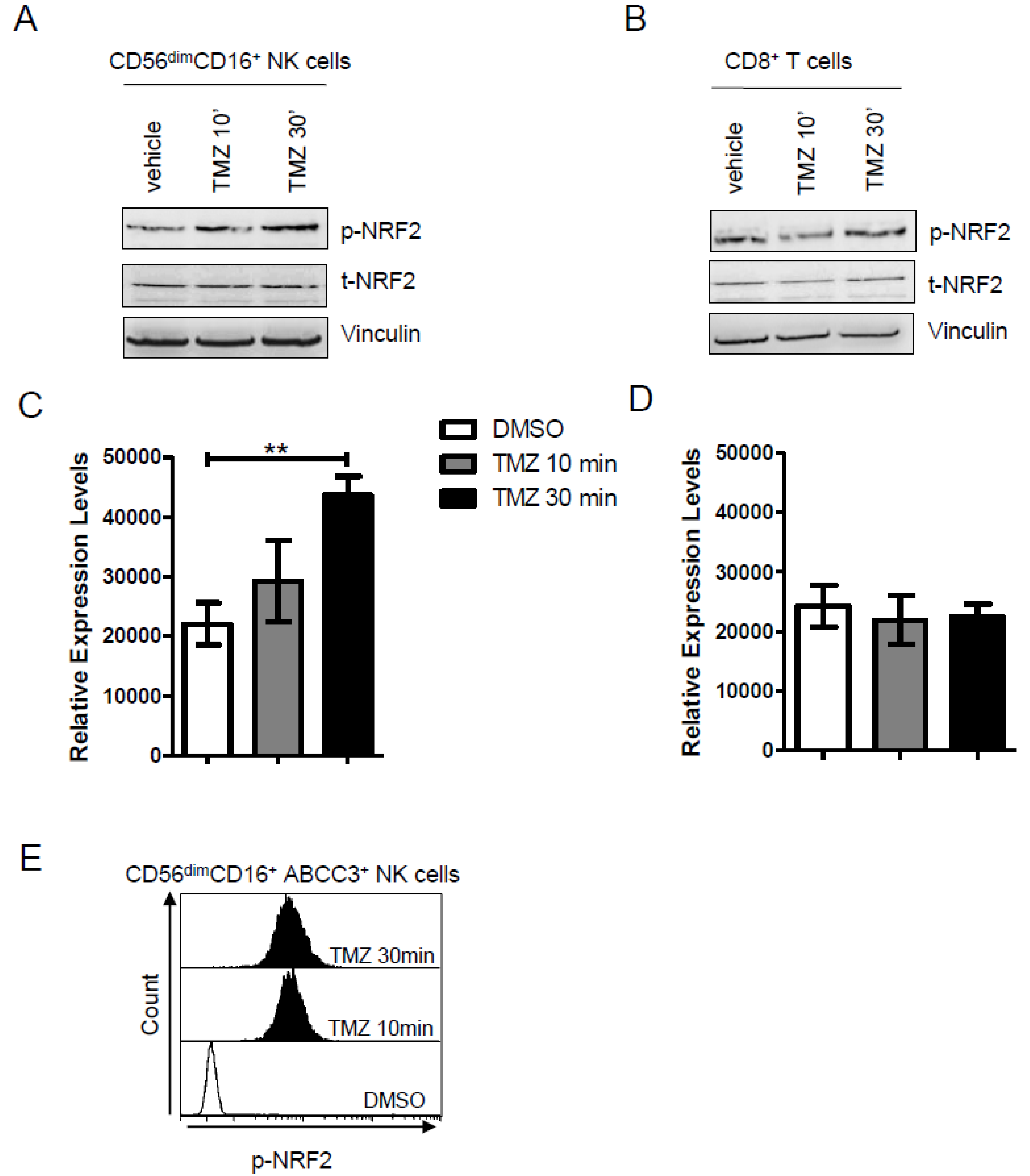
2.4. NRF2 Is Activated by TMZ Treatment in NK Cells Expressing ABCC3
3. Discussion
4. Materials and Methods
4.1. Patients and Treatment Protocol
4.2. Immunomonitoring
4.3. ABCC3 Transporter Activity
4.4. Western Blot
4.5. DNA Extraction and Genotyping
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Uribe, D.; Torres, Á.; Rocha, J.D.; Niechi, I.; Oyarzún, C.; Sobrevia, L.; San Martín, R.; Quezada, C. Multidrug resistance in glioblastoma stem-like cells: Role of the hypoxic microenvironment and adenosine signaling. Mol. Asp. Med. 2017, 55, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.B.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.-C.; Gorlia, T.; Hamou, M.-F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef]
- Auffinger, B.; Spencer, D.; Pytel, P.; Ahmed, A.U.; Lesniak, M.S. The role of glioma stem cells in chemotherapy resistance and glioblastoma multiforme recurrence. Expert Rev. Neurother. 2015, 15, 741–752. [Google Scholar] [CrossRef]
- Rieger, L.; Rieger, J.; Winter, S.; Streffer, J.; Esser, P.; Dichgans, J.; Meyermann, R.; Weller, M. Evidence for a constitutive, verapamil-sensitive, non-P-glycoprotein multidrug resistance phenotype in malignant glioma that is unaltered by radiochemotherapy in vivo. Acta Neuropathol. 2000, 99, 555–562. [Google Scholar] [CrossRef]
- Mohri, M.; Nitta, H.; Yamashita, J. Expression of multidrug resistance-associated protein (MRP) in human gliomas. J. Neurooncol. 2000, 49, 105–115. [Google Scholar] [CrossRef]
- Bähr, O.; Rieger, J.; Duffner, F.; Meyermann, R.; Weller, M.; Wick, W. P-glycoprotein and multidrug resistance-associated protein mediate specific patterns of multidrug resistance in malignant glioma cell lines, but not in primary glioma cells. Brain Pathol. 2003, 13, 482–494. [Google Scholar] [CrossRef]
- Calatozzolo, C.; Gelati, M.; Ciusani, E.; Sciacca, F.L.; Pollo, B.; Cajola, L.; Marras, C.; Silvani, A.; Vitellaro-Zuccarello, L.; Croci, D.; et al. Expression of drug resistance proteins Pgp, MRP1, MRP3, MRP5 and GST-pi in human glioma. J. Neurooncol. 2005, 74, 113–121. [Google Scholar] [CrossRef]
- Kuan, C.-T.; Wakiya, K.; Herndon, J.E.; Lipp, E.S.; Pegram, C.N.; Riggins, G.J.; Rasheed, A.; Szafranski, S.E.; McLendon, R.E.; Wikstrand, C.J.; et al. MRP3: A molecular target for human glioblastoma multiforme immunotherapy. BMC Cancer 2010, 10, 468. [Google Scholar] [CrossRef]
- Hirrlinger, J.; König, J.; Dringen, R. Expression of mRNAs of multidrug resistance proteins (Mrps) in cultured rat astrocytes, oligodendrocytes, microglial cells and neurones. J. Neurochem. 2002, 82, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Oselin, K.; Mrozikiewicz, P.M.; Pähkla, R.; Roots, I. Quantitative determination of the human MRP1 and MRP2 mRNA expression in FACS-sorted peripheral blood CD4+, CD8+, CD19+, and CD56+ cells. Eur. J. Haematol. 2003, 71, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Gray, J.; Kiosses, W.B.; Khurana, A.; Hitomi, K.; Crosby, C.M.; Chawla, A.; Fu, Z.; Zhao, M.; Veerapen, N.; et al. Mrp1 is involved in lipid presentation and iNKT cell activation by Streptococcus pneumoniae. Nat. Commun. 2018, 9, 4279. [Google Scholar] [CrossRef] [PubMed]
- Giannoudis, A.; Davies, A.; Harris, R.J.; Lucas, C.M.; Pirmohamed, M.; Clark, R.E. The clinical significance of ABCC3 as an imatinib transporter in chronic myeloid leukaemia. Leukemia 2014, 28, 1360–1363. [Google Scholar] [CrossRef]
- Yee, S.W.; Mefford, J.A.; Singh, N.; Percival, M.-E.; Stecula, A.; Yang, K.; Witte, J.S.; Takahashi, A.; Kubo, M.; Matsuda, K.; et al. Impact of polymorphisms in drug pathway genes on disease-free survival in adults with acute myeloid leukemia. J. Hum. Genet. 2013, 58, 353–361. [Google Scholar] [CrossRef]
- Pessina, S.; Cantini, G.; Kapetis, D.; Cazzato, E.; Di Ianni, N.; Finocchiaro, G.; Pellegatta, S. The multidrug-resistance transporter Abcc3 protects NK cells from chemotherapy in a murine model of malignant glioma. Oncoimmunology 2016, 5, e1108513. [Google Scholar] [CrossRef]
- Pellegatta, S.; Eoli, M.; Cuccarini, V.; Anghileri, E.; Pollo, B.; Pessina, S.; Frigerio, S.; Servida, M.; Cuppini, L.; Antozzi, C.; et al. Survival gain in glioblastoma patients treated with dendritic cell immunotherapy is associated with increased NK but not CD8+ T cell activation in the presence of adjuvant temozolomide. Oncoimmunology 2018, 7, e1412901. [Google Scholar] [CrossRef]
- Moretta, L. Dissecting CD56dim human NK cells. Blood 2010, 116, 3689–3691. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Turner, S.C.; Chen, K.S.; Ghaheri, B.A.; Ghayur, T.; Carson, W.E.; Caligiuri, M.A. Human natural killer cells: A unique innate immunoregulatory role for the CD56bright subset. Blood 2001, 97, 3146–3151. [Google Scholar] [CrossRef]
- Chiossone, L.; Chaix, J.; Fuseri, N.; Roth, C.; Vivier, E.; Walzer, T. Maturation of mouse NK cells is a 4-stage developmental program. Blood 2009, 113, 5488–5496. [Google Scholar] [CrossRef]
- Fu, B.; Wang, F.; Sun, R.; Ling, B.; Tian, Z.; Wei, H. CD11b and CD27 reflect distinct population and functional specialization in human natural killer cells. Immunology 2011, 133, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Hirota, T.; Ryokai, Y.; Kobayashi, D.; Kimura, M.; Irie, S.; Higuchi, S.; Ieiri, I. Systematic screening of human ABCC3 polymorphisms and their effects on MRP3 expression and function. Drug Metab. Pharm. 2011, 26, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Hitzl, M.; Burk, O.; Mornhinweg, E.; Keil, A.; Kerb, R.; Klein, K.; Zanger, U.M.; Eichelbaum, M.; Fromm, M.F. Genetic polymorphisms in the multidrug resistance-associated protein 3 (ABCC3, MRP3) gene and relationship to its mRNA and protein expression in human liver. Pharmacogenetics 2004, 14, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Canet, M.J.; Merrell, M.D.; Harder, B.G.; Maher, J.M.; Wu, T.; Lickteig, A.J.; Jackson, J.P.; Zhang, D.D.; Yamamoto, M.; Cherrington, N.J. Identification of a functional antioxidant response element within the eighth intron of the human ABCC3 gene. Drug Metab. Dispos. 2015, 43, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Souza-Fonseca-Guimaraes, F.; Cursons, J.; Huntington, N.D. The emergence of natural killer cells as a major target in cancer immunotherapy. Trends Immunol. 2019, 40, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Wennerberg, E.; Galluzzi, L. Born to kill: NK cells go to war against cancer. Trends Cancer 2019, 5, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- De Maria, A.; Bozzano, F.; Cantoni, C.; Moretta, L. Revisiting human natural killer cell subset function revealed cytolytic CD56dimCD16+ NK cells as rapid producers of abundant IFN-gamma on activation. Proc. Natl. Acad. Sci. USA 2011, 108, 728–732. [Google Scholar] [CrossRef]
- Jacobs, R.; Hintzen, G.; Kemper, A.; Beul, K.; Kempf, S.; Behrens, G.; Sykora, K.W.; Schmidt, R.E. CD56bright cells differ in their KIR repertoire and cytotoxic features from CD56dim NK cells. Eur. J. Immunol. 2001, 31, 3121–3127. [Google Scholar] [CrossRef]
- Chai, J.; He, Y.; Cai, S.-Y.; Jiang, Z.; Wang, H.; Li, Q.; Chen, L.; Peng, Z.; He, X.; Wu, X.; et al. Elevated hepatic multidrug resistance-associated protein 3/ATP-binding cassette subfamily C 3 expression in human obstructive cholestasis is mediated through tumor necrosis factor alpha and c-Jun NH2-terminal kinase/stress-activated protein kinase-signaling pathway. Hepatology 2012, 55, 1485–1494. [Google Scholar]
- Scheffer, G.L.; Kool, M.; de Haas, M.; de Vree, J.M.L.; Pijnenborg, A.C.L.M.; Bosman, D.K.; Elferink, R.P.J.O.; van der Valk, P.; Borst, P.; Scheper, R.J. Tissue distribution and induction of human multidrug resistant protein 3. Lab. Investig. 2002, 82, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Takechi, T.; Hirota, T.; Sakai, T.; Maeda, N.; Kobayashi, D.; Ieiri, I. Interindividual differences in the expression of ATP-binding cassette and solute carrier family transporters in human skin: DNA methylation regulates transcriptional activity of the human ABCC3 gene. Drug Metab. Dispos. 2018, 46, 628–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Ven, R.; Oerlemans, R.; van der Heijden, J.W.; Scheffer, G.L.; de Gruijl, T.D.; Jansen, G.; Scheper, R.J. ABC drug transporters and immunity: Novel therapeutic targets in autoimmunity and cancer. J. Leukoc. Biol. 2009, 86, 1075–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.H.; Denton, M.D.; Alexander, S.I.; Khoury, S.J.; Sayegh, M.H.; Briscoe, D.M. Specific MDR1 P-glycoprotein blockade inhibits human alloimmune T cell activation in vitro. J. Immunol. 2001, 166, 2451–2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köck, K.; Grube, M.; Jedlitschky, G.; Oevermann, L.; Siegmund, W.; Ritter, C.A.; Kroemer, H.K. Expression of adenosine triphosphate-binding cassette (ABC) drug transporters in peripheral blood cells: Relevance for physiology and pharmacotherapy. Clin. Pharm. 2007, 46, 449–470. [Google Scholar]
- Borg, C.; Terme, M.; Taïeb, J.; Ménard, C.; Flament, C.; Robert, C.; Maruyama, K.; Wakasugi, H.; Angevin, E.; Thielemans, K.; et al. Novel mode of action of c-kit tyrosine kinase inhibitors leading to NK cell-dependent antitumor effects. J. Clin. Investig. 2004, 114, 379–388. [Google Scholar] [CrossRef]
- Mahadevan, D.; Riley, C.; Simons, D.; Della Croce, K.; Wisner, L.; Iorio, M.; Garewal, H.; Bearss, D. Mechanisms of gleevec resistance in GIST and potential therapeutic interventions. Am. Assoc. Cancer Res. 2005, 65, 1197–1198. [Google Scholar]
- Eoli, M.; Menghi, F.; Bruzzone, M.G.; De Simone, T.; Valletta, L.; Pollo, B.; Bissola, L.; Silvani, A.; Bianchessi, D.; D’Incerti, L.; et al. Methylation of O6-methylguanine DNA methyltransferase and loss of heterozygosity on 19q and/or 17p are overlapping features of secondary glioblastomas with prolonged survival. Clin. Cancer Res. 2007, 13, 2606–2613. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients (n = 30) | Age/Gender | MGMT (Met ≥ 0.1) | NK Cell Response § | SNP | PFS (months) | OS (months) |
|---|---|---|---|---|---|---|
| 1 | 55/F | U (0.07) | NO | DelC | 13.7 | 22.5 |
| 2 | 62/F | U (0.01) | NO | Wt | 12.0 | 24.4 |
| 3 | 66/M | U (0.04) | YES | DelC | 15.4 | 15.4 |
| 4 | 70/F | U (0.00) | NO | Wt | 14.7 | 17.8 |
| 5 | 49/M | U (0.00) | NO | DelC | 10.2 | 12.5 |
| 6 | 65/F | M (0.71) | YES | DelC | 20.8 | 33.9 |
| 7 | 60/M | U (0.01) | YES | DelC | 9.3 | 25.0 |
| 8 | 58/M | U (0.00) | YES | DelC | 9.4 | 22.6 |
| 9 | 50/M | U (0.00) | YES | DelC | 16.1 | 33.0 |
| 10 | 48/M | M (2.38) | YES | DelC | 4.4 | 7.8 |
| 11 | 23/F | U (0.003) | NO | Wt | 3.1 | 6.4 |
| 12 | 44/M | U (0.03) | YES | DelC | 24.2 | 38.4 |
| 14 | 62/M | M (0.46) | NO | DelC | 7.9 | 20.2 |
| 16 | 70/M | M (1.50) | YES | DelC | 17.2 | 32.8 |
| 19 | 56/M | U (0.00) | NO | Wt | 3.2 | 6.9 |
| 20 | 48/M | U (0.00) | NO | Wt | 9.0 | 12.4 |
| 21 | 53/F | M (0.47) | YES | DelC | 28.3 | 56.0 |
| 22 | 63/M | U (0.02) | YES | Wt | 6.5 | 8.1 |
| 23 | 45/M | M (0.74) | YES | DelC | >60.0 | >60.0 |
| 24 | 55/F | U (0.00) | YES | DelC | 14.0 | 28.3 |
| 25 | M/58 | M (0.18) | NO | DelC | 22.4 | 37.2 |
| 26 | F/45 | U (0.07) | NO | Wt | 12.0 | 17.6 |
| 27 | M/49 | M (0.38) | NO | Wt | 9.5 | 13.2 |
| 28 | F/54 | U (0.00) | NO | Wt | 12.0 | 15.5 |
| 29 | M/43 | M (0.56) | NO | Wt | 9.4 | 17.3 |
| 30 | M/65 | M (0.26) | NO | Wt | 18.7 | 27.0 |
| 31 | F/60 | M (1.78) | YES | Wt | 10.7 | 22.9 |
| 32 | M/62 | M (0.21) | NO | Wt | 16.1 | 33.5 |
| 33 | M/53 | U (0.04) | YES | Wt | 18.3 | 38.5 |
| 34 | M/49 | M (2.39) | YES | DelC | 12.0 | 23.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pellegatta, S.; Di Ianni, N.; Pessina, S.; Paterra, R.; Anghileri, E.; Eoli, M.; Finocchiaro, G. ABCC3 Expressed by CD56dim CD16+ NK Cells Predicts Response in Glioblastoma Patients Treated with Combined Chemotherapy and Dendritic Cell Immunotherapy. Int. J. Mol. Sci. 2019, 20, 5886. https://doi.org/10.3390/ijms20235886
Pellegatta S, Di Ianni N, Pessina S, Paterra R, Anghileri E, Eoli M, Finocchiaro G. ABCC3 Expressed by CD56dim CD16+ NK Cells Predicts Response in Glioblastoma Patients Treated with Combined Chemotherapy and Dendritic Cell Immunotherapy. International Journal of Molecular Sciences. 2019; 20(23):5886. https://doi.org/10.3390/ijms20235886
Chicago/Turabian StylePellegatta, Serena, Natalia Di Ianni, Sara Pessina, Rosina Paterra, Elena Anghileri, Marica Eoli, and Gaetano Finocchiaro. 2019. "ABCC3 Expressed by CD56dim CD16+ NK Cells Predicts Response in Glioblastoma Patients Treated with Combined Chemotherapy and Dendritic Cell Immunotherapy" International Journal of Molecular Sciences 20, no. 23: 5886. https://doi.org/10.3390/ijms20235886
APA StylePellegatta, S., Di Ianni, N., Pessina, S., Paterra, R., Anghileri, E., Eoli, M., & Finocchiaro, G. (2019). ABCC3 Expressed by CD56dim CD16+ NK Cells Predicts Response in Glioblastoma Patients Treated with Combined Chemotherapy and Dendritic Cell Immunotherapy. International Journal of Molecular Sciences, 20(23), 5886. https://doi.org/10.3390/ijms20235886