Diagnostic Cytokines and Comparative Analysis Secreted from Exfoliated Deciduous Teeth, Dental Pulp, and Bone Marrow Derived Mesenchymal Stem Cells for Functional Cell-Based Therapy

Abstract
:1. Introduction
2. Results
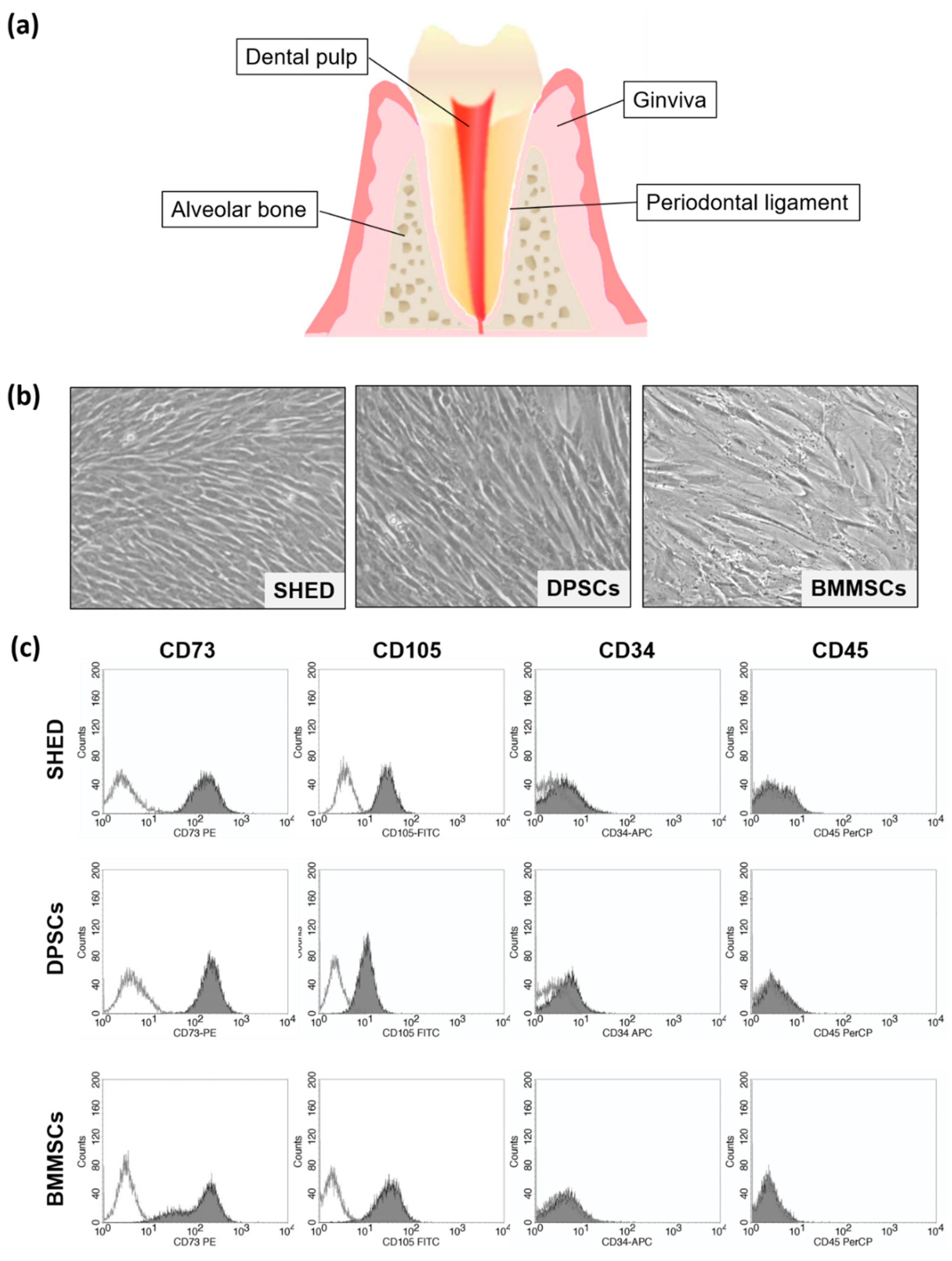
2.1. Characterization of SHED (Stem Cells from Exfoliated Deciduous Teeth), DPSCs (Dental Pulp Stem Cells), and BMMSCs (Bone Marrow-Derived Mesenchymal Stem Cells)
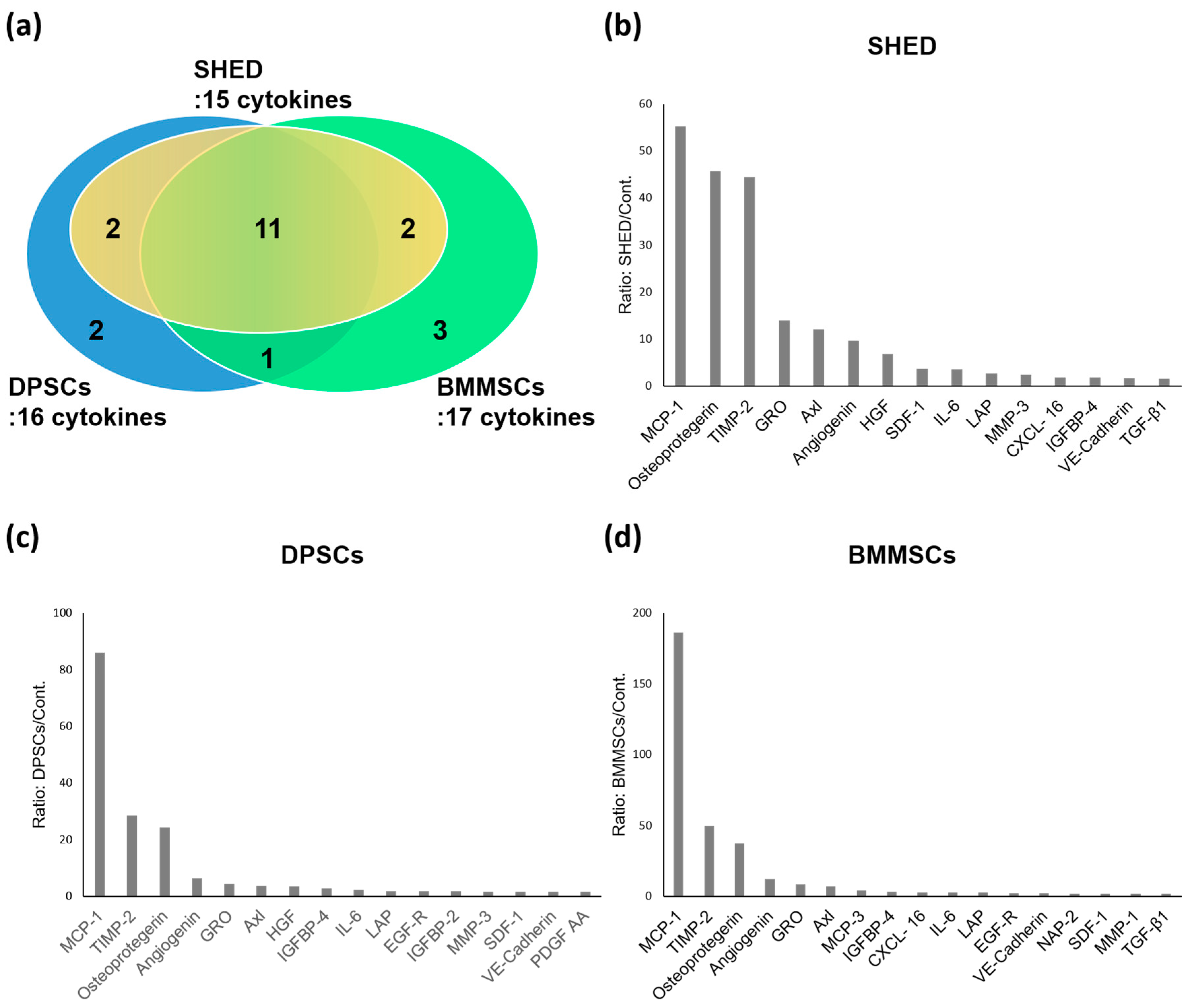
2.2. Cytokines Secreted from SHED, DPSCs, and BMMSCs
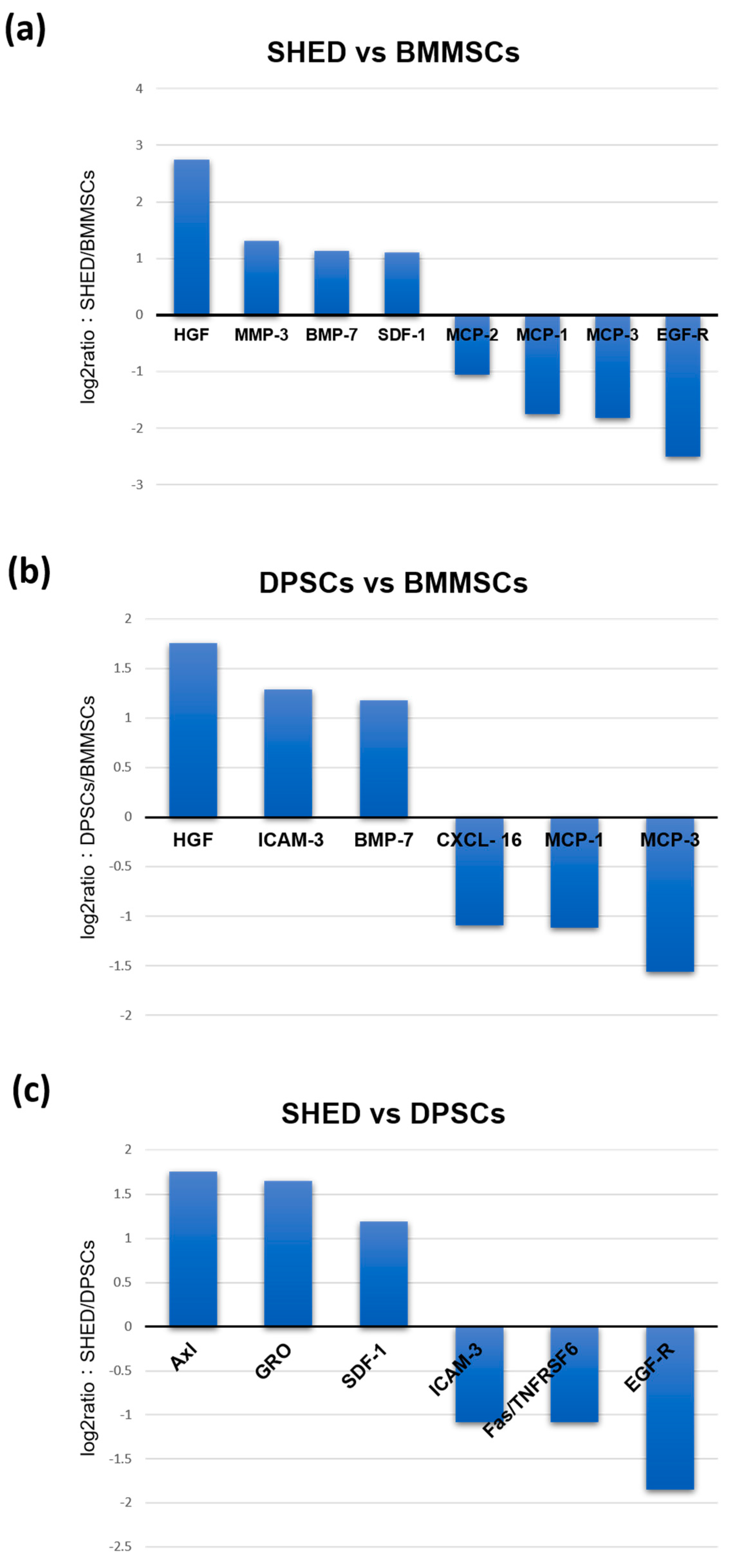
2.3. Comparison of Cytokines Expression Levels of SHED, DPSCs, and BMMSCs
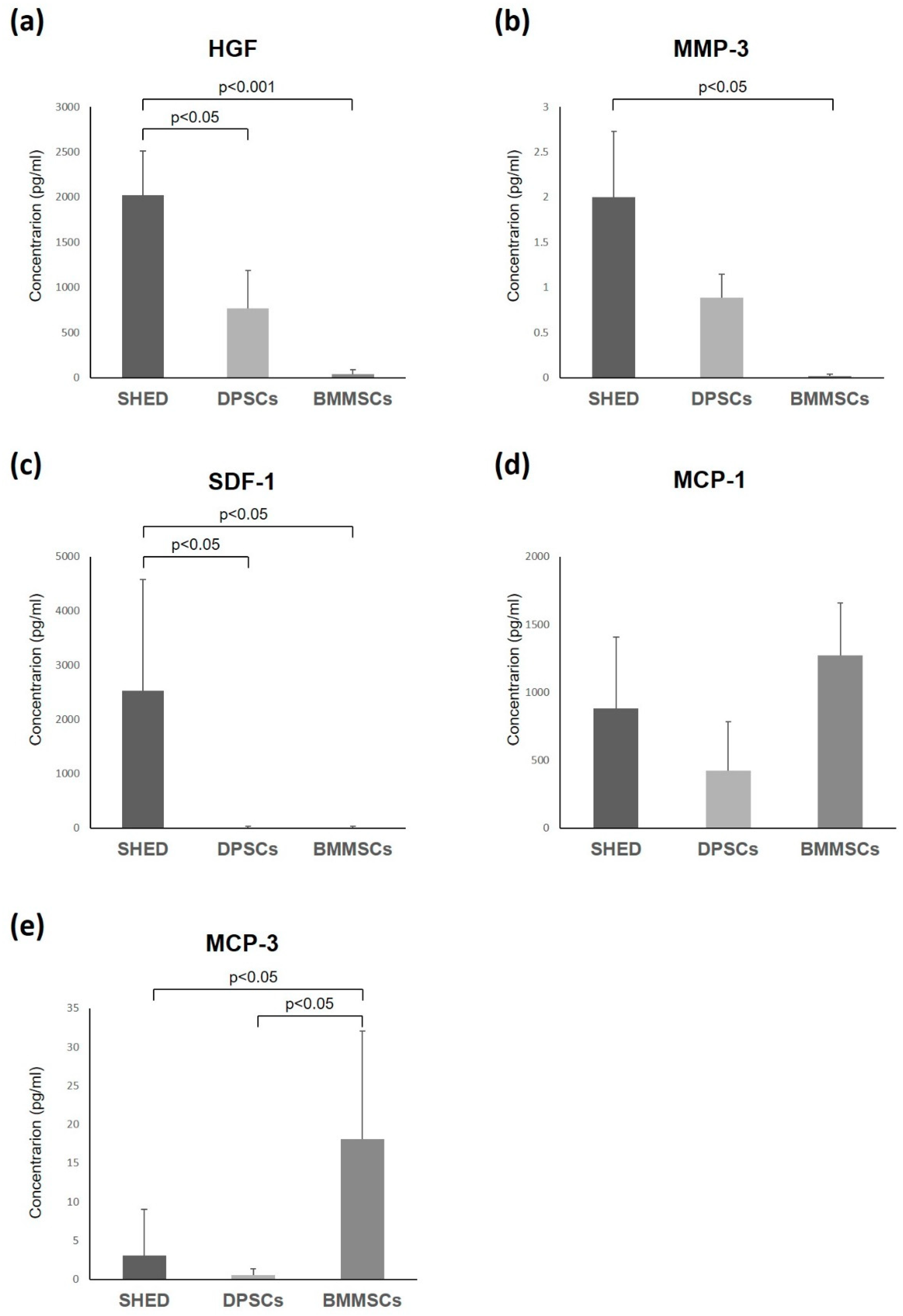
2.4. Enzyme-Linked Immunosorbent Assay
3. Discussion
4. Materials and Methods
4.1. Subjects and Cell Cultures
4.2. Flow Cytometry Analyses
4.3. Cytokine Antibody Array Analysis
4.4. Enzyme-Linked Immunosorbent Assay
4.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MSCs | mesenchymal stem/stromal cells |
| SHED | stem cells from exfoliated deciduous teeth |
| DPSCs | dental pulp stem cells |
| BMMSCs | bone marrow-derived mesenchymal stem cells |
| MCP-1 | monocyte chemoattractant protein-1 |
| TIMP-2 | tissue inhibitor of metalloproteinase-2 |
| GRO | growth regulated oncogene |
| SDF-1 | stromal cell derived factor 1 |
| IL-6 | interleukin-6 |
| LAP | latency associated peptide |
| IGFBP-4 | insulin-like growth factor binding protein-4 |
| VE-Cadherin | vascular endothelial cadherin |
| HGF | hepatocyte growth factor |
| MMP-3 | matrix metalloproteinase-3 |
| MCP-3 | monocyte chemoattractant protein-1 |
| NAP-2 | neutrophil activating protein-2 |
| MMP-1 | matrix metalloproteinase-1 |
| TGF-β1 | Transforming growth factor beta 1 |
| EGF-R | epidermal growth factor receptor |
| IGFBP-2 | insulin-like growth factor binding protein-2 |
| PDGF AA | platelet derived growth factor AA |
| CXCL-16 | C-X-C motif ligand-16 |
| ELISA | enzyme-linked immunosorbent assay |
| SCI | spinal cord injury |
| MMPs | matrix metalloproteinases |
| ECM | extracellular matrix |
| CXCR4 | CXC receptor 4 |
| DMEM | Dulbecco’s modified Eagle medium |
| PBS | phosphate-buffered saline |
References
- Pittenger, M.F.; Mackay, A.M.; Beck, C.B.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craiq, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Nakamura-Yamada, S.; Kusano, K.; Baba, S. Clinical Potential and Current Progress of Dental Pulp Stem Cells for Various Systemic Diseases in Regenerative Medicine: A Concise Review. Int. J. Mol. Sci. 2019, 20, 1132. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Young, H.E.; Steele, T.A.; Bray, R.A.; Hudson, J.; Floyd, J.A.; Hawkins, K.; Thomas, K.; Austin, T.; Edwards, C.; Cuzzourt, J.; et al. Human reserve pluripotent mesenchymal stem cells are present in the connective tissues of skeletal muscle and dermis derived from fetal, adult, and geriatric donors. Anat. Rec. 2001, 264, 51–62. [Google Scholar] [CrossRef]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef]
- Yamada, Y.; Nakamura, S.; Ito, K.; Sugito, T.; Yoshimi, R.; Nagasaka, T.; Ueda, M. A feasibility of useful cell-based therapy by bone regeneration with deciduous tooth stem cells, dental pulp stem cells, or bone-marrow-derived mesenchymal stem cells for clinical study using tissue engineering technology. Tissue Eng. Part A 2010, 16, 1891–1900. [Google Scholar] [CrossRef]
- Yamada, Y.; Ito, K.; Nakamura, S.; Ueda, M.; Nagasaka, T. Promising cell-based therapy for bone regeneration using stem cells from deciduous teeth, dental pulp, and bone marrow. Cell Transplant. 2011, 20, 1003–1013. [Google Scholar] [CrossRef]
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Eleuteri, S.; Fierabracci, A. Insights into the Secretome of Mesenchymal Stem Cells and Its Potential Applications. Int. J. Mol. Sci. 2019, 20, 4597. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Fujimoto, A.; Ito, A.; Yoshimi, R.; Ueda, M. Cluster analysis and gene expression profiles: A cDNA microarray system-based comparison between human dental pulp stem cells (hDPSCs) and human mesenchymal stem cells (hMSCs) for tissue engineering cell therapy. Biomaterials 2006, 27, 3766–3781. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Yamada, Y.; Katagiri, W.; Sugito, T.; Ito, K.; Ueda, M. Stem cell proliferation pathways comparison between human exfoliated deciduous teeth and dental pulp stem cells by gene expression profile from promising dental pulp. J. Endod. 2009, 35, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Yamada, Y.; Nakamura, S.; Umemura, E.; Ito, K.; Ueda, M. Potential characteristics of stem cells from human exfoliated deciduous teeth compared with bone marrow-derived mesenchymal stem cells for mineralized tissue-forming cell biology. J. Endod. 2011, 37, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Nakaumura, S.; Ito, K.; Umemura, E.; Hara, K.; Nagasaka, T.; Abe, A.; Baba, S.; Furuichi, Y.; Izumi, Y.; et al. Injectable Bone Tissue Engineering Using Expanded Mesenchymal Stem Cells. Stem Cells 2013, 31, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC—Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.; Dunnill, P. A brief definition of regenerative medicine. Regen. Med. 2008, 3, 1–5. [Google Scholar] [CrossRef]
- Su, X.; Paris, M.; Gi, Y.J.; Tsai, K.Y.; Cho, M.S.; Lin, Y.L.; Biernaskie, J.A.; Sinha, S.; Prives, C.; Pevny, L.H.; et al. TAp63 prevents premature aging by promoting adult stem cell maintenance. Cell Stem Cell 2009, 5, 64–75. [Google Scholar] [CrossRef]
- Miller, F.D.; Kaplan, D.R. Mobilizing endogenous stem cells for repair and regeneration: Are we there yet? Cell Stem Cell 2012, 10, 650–652. [Google Scholar] [CrossRef]
- Russell, W.E.; McGowan, J.A.; Bucher, N.L. Partial characterization of a hepatocyte growth factor from rat platelets. J. Cell. Physiol. 1984, 119, 183–192. [Google Scholar] [CrossRef]
- Graziani, A.; Gramaglia, D.; dalla Zonca, P.; Comoglio, P.M. Hepatocyte growth factor/scatter factor stimulates the Ras-guanine nucleotide exchanger. J. Biol. Chem. 1993, 268, 9165–9168. [Google Scholar] [PubMed]
- Zarnegar, R.; Michalopoulos, G.K. The many faces of hepatocyte growth factor: From hepatopoiesis to hematopoiesis. J. Cell Biol. 1995, 129, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Shibata, R.; Yamamoto, N.; Nishikawa, M.; Hibi, H.; Tanigawa, T.; Ueda, M.; Murohara, T.; Yamamoto, A. Dental pulp-derived stem cell conditioned medium reduces cardiac injury following ischemia-reperfusion. Sci. Rep. 2015, 6, 16295. [Google Scholar] [CrossRef] [PubMed]
- Hirata, M.; Ishigami, M.; Matsushita, Y.; Ito, T.; Hattori, H.; Hibi, H.; Goto, H.; Ueda, M.; Yamamoto, A. Multifaceted Therapeutic Benefits of Factors Derived From Dental Pulp Stem Cells for Mouse Liver Fibrosis. Stem Cells Transl. Med. 2016, 5, 1416–1424. [Google Scholar] [CrossRef]
- Kitamura, K.; Nagoshi, N.; Tsuji, O.; Matsumoto, M.; Okano, H.; Nakamura, M. Application of Hepatocyte growth factor for acute spinal cord injury: The road from basic studies to human treatnment. Int. J. Mol. Sci. 2019, 20, 1054. [Google Scholar] [CrossRef]
- Kitamura, K.; Iwanami, A.; Nakamura, M.; Yamane, J.; Watanabe, K.; Suzuki, Y.; Miyazawa, D.; Shibata, S.; Funakoshi, H.; Miyatake, S.; et al. Hepatocyte growth factor promotes endogenous repair and functional recovery after spinal cord injury. J. Neurosci. Res. 2007, 85, 2332–2342. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar]
- Rundhaug, J.E. Matrix metalloproteinases and angiogenesis. J. Cell Mol. Med. 2005, 9, 267–285. [Google Scholar] [CrossRef]
- Nagase, H.; Woessner, J.F., Jr. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef]
- Fini, M.E.; Cook, J.R.; Mohan, R.; Brinckerhoft, C.E. Matrix Metalloproteinases; Parks, W.C., Mecham, R.P., Eds.; Academic Press: San Diego, CA, USA, 1998; pp. 299–356. [Google Scholar]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Kubota, S.; Kawata, K.; Mukudai, Y.; Uehara, J.; Ohgawara, T.; Ibaragi, S.; Sasaki, A.; Kuboki, T.; Takigawa, M. Novel transcription-factor-like function of human matrix metalloproteinase 3 regulating the CTGF/CCN2 gene. Mol. Cell. Biol. 2008, 28, 2391–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haro, H.; Crawford, H.C.; Fingleton, B.; MacDougall, J.R.; Shinomiya, K.; Spengler, D.M.; Matrisian, L.M. Matrix metalloproteinase-3-dependent generation of a macrophage chemoattractant in a model of herniated disc resorption. J. Clin. Investig. 2000, 105, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, K.; Kawashima, N.; Suzuki, N.; Koizumi, Y.; Yamamoto, M.; Nakashima, M.; Suda, H. Down-regulation of inflammatory mediator synthesis and infiltration of inflammatory cells by MMP-3 in experimentally induced rat pulpitis. J. Endod. 2014, 40, 1404–1409. [Google Scholar] [CrossRef]
- Zheng, L.; Amano, K.; Iohara, K.; Ito, M.; Imabayashi, K.; Into, T.; Matsushita, K.; Nakamura, H.; Nakashima, M. Matrix metalloproteinase-3 accelerates wound healing following dental pulp injury. Am. J. Pathol. 2009, 175, 1905–1914. [Google Scholar] [CrossRef] [Green Version]
- Nishino, Y.; Yamada, Y.; Ebisawa, K.; Nakamura, S.; Okabe, K.; Umemura, E.; Hara, K.; Ueda, M. Stem cells from human exfoliated deciduous teeth (SHED) enhance wound healing and the possibility of novel cell therapy. Cytotherapy 2011, 13, 598–605. [Google Scholar] [CrossRef]
- Liekens, S.; Schols, D.; Hatse, S. CXCL12-CXCR4 axis in angiogenesis, metastasis and stem cell mobilization. Curr. Pharm. Des. 2010, 16, 3903–3920. [Google Scholar] [CrossRef]
- Song, M.; Jang, H.; Lee, J.; Kim, J.H.; Kim, S.H.; Sun, K.; Park, Y. Regeneration of chronic myocardial infarction by injectable hydrogels containing stem cell homing factor SDF-1 and angiogenic peptide Ac-SDKP. Biomaterials 2014, 35, 2436–2445. [Google Scholar] [CrossRef]
- Ding, B.S.; Cao, Z.; Lis, R.; Nolan, D.J.; Guo, P.; Simons, M.; Penfold, M.E.; Shido, K.; Rabbany, S.Y.; Rafii, S. Divergent angiocrine signals from vascular niche balance liver regeneration and fibrosis. Nature 2014, 505, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Heskamp, A.; Leibinger, M.; Andreadaki, A.; Gobrecht, P.; Diekmann, H.; Fischer, D. CXCL12/SDF-1 facilitates optic nerve regeneration. Neurobiol. Dis. 2013, 55, 76–86. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.; Tao, J.; Jiang, Y.; Hu, C.; Huang, L.; Ji, J.; Ouyang, H.W. The use of type 1 collagen scaffold containing stromal cell-derived factor-1 to create a matrix environment conducive to partial-thickness cartilage defects repair. Biomaterials 2013, 34, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Yang, F.; Ma, J.; Bouma, M.J.; Boerman, O.C.; Chen, Z.; van den Beucken, J.J.; Jansen, J.A. Incorporation of stromal cell-derived factor-1alpha in PCL/gelatin electrospun membranes for guided bone regeneration. Biomaterials 2013, 34, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, Y.S.; Bae, W.J.; Lee, H.J.; Chang, S.W.; Kim, W.S.; Kim, E.C. The role of SDF-1 and CXCR4 on odontoblastic differentiation in human dental pulp cells. Int. Endod. J. 2014, 47, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Iohara, K.; Imabayashi, K.; Ishizaka, R.; Watanabe, A.; Nabekura, J.; Ito, M.; Matsushita, K.; Nakamura, H.; Nakashima, M. Complete pulp regeneration after pulpectomy by transplantation of CD105+ stem cells with stromal cell-derived factor-1. Tissue Eng. Part A 2011, 17, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Tsou, C.L.; Peters, W.; Si, Y.; Slaymaker, S.; Aslanian, A.M.; Weisberg, S.P.; Mack, M.; Charo, I.F. Critical roles for CCR2 and MCP-3 in monocyte mobilization from bone marrow and recruitment to inflammatory sites. J. Clin. Investig. 2007, 117, 902–909. [Google Scholar] [CrossRef] [Green Version]
- Schenk, S.; Mal, N.; Finan, A.; Zhang, M.; Kiedrowski, M.; Popovic, Z.; McCarthy, P.M.; Penn, M.S. Monocyte chemotactic protein-3 is a myocardial mesenchymal stem cell homing factor. Stem Cells 2007, 25, 245–251. [Google Scholar] [CrossRef]
- Menten, P.; Wuyts, A.; Van Damme, J. Monocyte chemotactic protein-3. Eur. Cytokine Netw. 2001, 12, 554–560. [Google Scholar]
- Shinohara, K.; Greenfield, S.; Pan, H.; Vasanji, A.; Kumagai, K.; Midura, R.J.; Kiedrowski, M.; Penn, M.S.; Muschler, G.F. Stromal cell-derived factor-1 and monocyte chemotactic protein-3 improve recruitment of osteogenic cells into sites of musculoskeletal repair. J. Orthop. Res. 2011, 29, 1064–1069. [Google Scholar] [CrossRef]
- Axelrod, H.; Pienta, K.J. Axl as a mediator of cellular growth and survival. Oncotarget 2014, 5, 8818–8852. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokines | SHEDs | DPSCs | BMMSCs | |
|---|---|---|---|---|
| Growth factors | Angiogenin | ++ | ++ | ++ |
| HGF | ++ | ++ | ― | |
| TGF-β1 | + | ― | + | |
| EGF-R | ― | + | ++ | |
| IGFBP-2 | ― | + | ― | |
| IGFBP-4 | + | ++ | ++ | |
| PDGF AA | ― | + | ― | |
| Chemokines | MCP-1 | ++ | ++ | ++ |
| MCP-3 | ― | ― | ++ | |
| GRO | ++ | ++ | ++ | |
| SDF-1 | ++ | + | + | |
| CXCL- 16 | + | ― | ++ | |
| NAP-2 | ― | ― | + | |
| Inflammatory cytokines | Osteoprotegerin | ++ | ++ | ++ |
| TIMP-2 | ++ | ++ | ++ | |
| IL-6 | ++ | ++ | ++ | |
| LAP | ++ | + | ++ | |
| Other cytokines | Axl | ++ | ++ | ++ |
| VE-Cadherin | + | + | + | |
| MMP-1 | ― | ― | + | |
| MMP-3 | ++ | + | ― | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamada, Y.; Nakamura-Yamada, S.; Umemura-Kubota, E.; Baba, S. Diagnostic Cytokines and Comparative Analysis Secreted from Exfoliated Deciduous Teeth, Dental Pulp, and Bone Marrow Derived Mesenchymal Stem Cells for Functional Cell-Based Therapy. Int. J. Mol. Sci. 2019, 20, 5900. https://doi.org/10.3390/ijms20235900
Yamada Y, Nakamura-Yamada S, Umemura-Kubota E, Baba S. Diagnostic Cytokines and Comparative Analysis Secreted from Exfoliated Deciduous Teeth, Dental Pulp, and Bone Marrow Derived Mesenchymal Stem Cells for Functional Cell-Based Therapy. International Journal of Molecular Sciences. 2019; 20(23):5900. https://doi.org/10.3390/ijms20235900
Chicago/Turabian StyleYamada, Yoichi, Sayaka Nakamura-Yamada, Eri Umemura-Kubota, and Shunsuke Baba. 2019. "Diagnostic Cytokines and Comparative Analysis Secreted from Exfoliated Deciduous Teeth, Dental Pulp, and Bone Marrow Derived Mesenchymal Stem Cells for Functional Cell-Based Therapy" International Journal of Molecular Sciences 20, no. 23: 5900. https://doi.org/10.3390/ijms20235900
APA StyleYamada, Y., Nakamura-Yamada, S., Umemura-Kubota, E., & Baba, S. (2019). Diagnostic Cytokines and Comparative Analysis Secreted from Exfoliated Deciduous Teeth, Dental Pulp, and Bone Marrow Derived Mesenchymal Stem Cells for Functional Cell-Based Therapy. International Journal of Molecular Sciences, 20(23), 5900. https://doi.org/10.3390/ijms20235900