On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
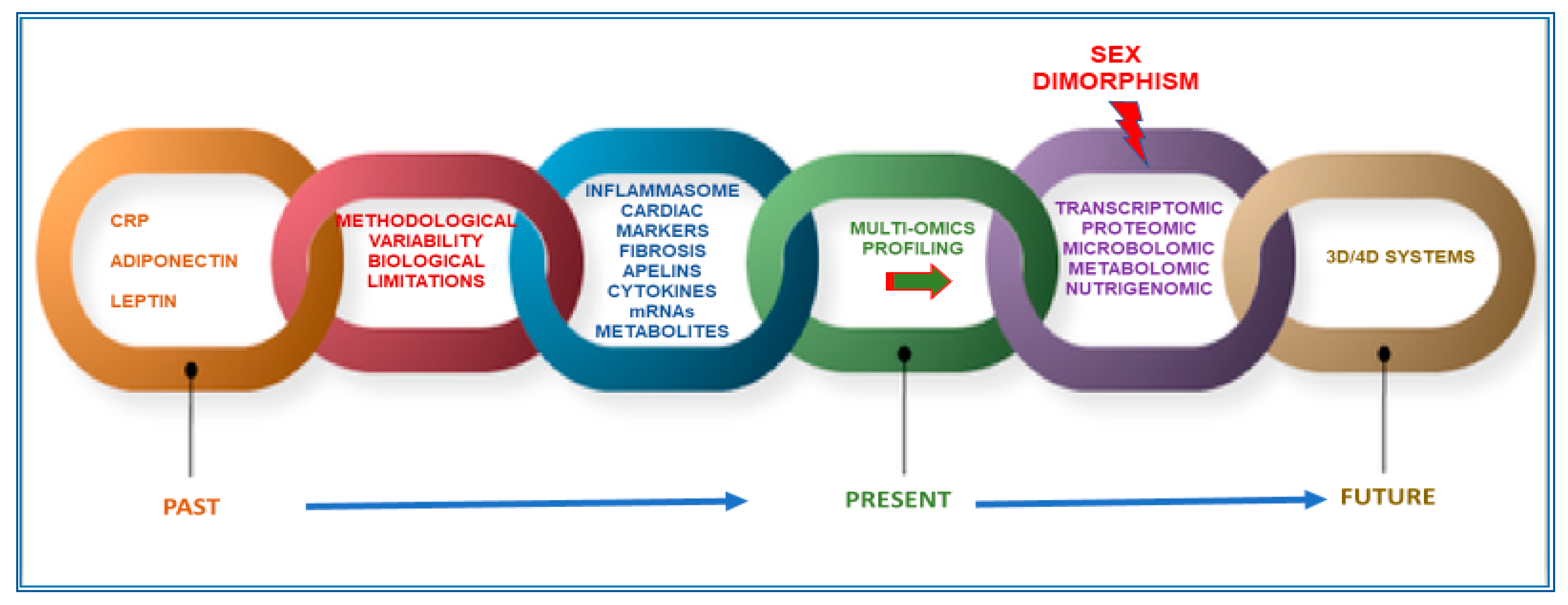
:1. Challenges and Drawbacks in the Research of Appropriate Biomarkers for the Management of Cardiometabolic Diseases
2. Multi-Omics Approaches, 3D and 4D Systems, and Innovative Techniques to Search for More Appropriate CMD Biomarkers: Reasons, Gaps, and Limitations
2.1. A Focus on Metabolomics and Metabolite Profiling and Examples of Related CMD Biomarkers
2.2. Microbiomic Profiling and Related Promising CMB Predictive Biomarker
2.3. Nutrigenomics and Nutrigenetics Approaches in the Research of CMD Biomarkers
2.4. The Difficult Challenge of Integration and Interpretation of Multidimensional Data from Multi-Omics Analyses
3. Adding Another Important Layer: Sex Dimorphism in CMD and its Relevance in the Research of Accurate Sex or Gender Biomarkers
4. Age and Medication: Other Factors to Consider in the Search of CMD Biomarkers and Strategies for Reducing their Effects
5. Another Revolutionary Approach to Consider in CMD Biomarkers Research: Extracellular Vesicles
6. Conclusions and Perspectives
Author Contributions
Disclaimer
Funding
Conflicts of Interest
Abbreviations
| BCAA | branched-chain amino acids |
| CMD | cardiometabolic diseases |
| CRISPR/Cas9 | clustered regularly interspaced short palindromic repeats -associated protein 9 |
| GWAS | genome-wide association studies |
| MVs | microvesicles |
| NO | nitric oxide |
| PI3K/AKT/mTOR | phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin |
| TLR | toll-like receptor |
| TMA | trimethylamine |
| TMAO | trimethylamine N-oxide |
References
- Oda, E. Historical perspectives of the metabolic syndrome. Clin. Derm. 2018, 36, 3–8. [Google Scholar] [CrossRef] [PubMed]
- McCracken, E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the metabolic syndrome. Clin. Derm. 2018, 36, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Kaptoge, S.; Di Angelantonio, E.; Lowe, G.; Pepys, M.B.; Thompson, S.G.; Collins, R.; Danesh, J. C-reactive protein concentration and risk of coronary heart disease, stroke, and mortality: An individual participant meta-analysis. Lancet 2010, 375, 132–140. [Google Scholar] [PubMed]
- Sattar, N.; Murray, H.M.; McConnachie, A.; Blauw, G.J.; Bollen, E.L.; Buckley, B.M.; Cobbe, S.M.; Ford, I.; Gaw, A.; Hyland, M.; et al. C-reactive protein and prediction of coronary heart disease and global vascular events in the Prospective Study of Pravastatin in the Elderly at Risk (PROSPER). Circulation 2007, 115, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Wensley, F.; Gao, P.; Burgess, S.; Kaptoge, S.; Di Angelantonio, E.; Shah, T.; Engert, J.C.; Clarke, R.; Davey-Smith, G.; Nordestgaard, B.G.; et al. Association between C reactive protein and coronary heart disease: Mendelian randomisation analysis based on individual participant data. BMJ 2011, 342, d548. [Google Scholar]
- Kanhai, D.A.; Kranendonk, M.E.; Uiterwaal, C.S.; van der Graaf, Y.; Kappelle, L.J.; Visseren, F.L. Adiponectin and incident coronary heart disease and stroke. A systematic review and meta-analysis of prospective studies. Obes. Rev. 2013, 14, 555–567. [Google Scholar] [CrossRef]
- Sook Lee, E.; Park, S.S.; Kim, E.; Sook Yoon, Y.; Ahn, H.Y.; Park, C.Y.; Ho Yun, Y.; Woo Oh, S. Association between adiponectin levels and coronary heart disease and mortality: A systematic review and meta-analysis. Int. J. Epidemiol. 2013, 42, 1029–1039. [Google Scholar] [CrossRef]
- Borges, M.C.; Lawlor, D.A.; de Oliveira, C.; White, J.; Horta, B.L.; Barros, A.J. Role of Adiponectin in coronary heart disease risk: A Mendelian randomization study. Circ. Res. 2016, 22, 119491–119499. [Google Scholar] [CrossRef]
- Roujeau, C.; Jockers, R.; Dam, J. New pharmacological perspectives for the leptin receptor in the treatment of obesity. Front. Endocrinol. 2014, 5, 167. [Google Scholar] [CrossRef]
- Zeng, R.; Xu, C.H.; Xu, Y.N.; Wang, Y.L.; Wang, M. Association of leptin levels with pathogenetic risk of coronary heart disease and stroke: A meta-analysis. Arq. Bras. Endocrinol. Metabol. 2014, 58, 817–823. [Google Scholar] [CrossRef]
- Yang, H.; Guo, W.; Li, J.; Cao, S.; Zhang, J.; Pan, J.; Wang, Z.; Wen, P.; Sh, X.; Zhang, S. Leptin concentration and risk of coronary heart disease and stroke: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0166360. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.B.; Sun, F.; Nie, X.L.; Wang, J. Leptin and coronary heart disease: A systematic review and meta-analysis. Atherosclerosis 2014, 233, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Sun, D. Leptin receptor gene polymorphism and the risk of cardiovascular disease: A systemic review and meta-analysis. Int. J. Environ. Res. Public Health 2017, 14, 375. [Google Scholar] [CrossRef] [PubMed]
- Janket, S.J.; Javaheri, H.; Ackerson, L.K.; Ayilavarapu, S.; Meurman, J.H. Oral infections, metabolic inflammation, genetics, and cardiometabolic diseases. J. Dent. Res. 2015, 94 (Suppl. 9), 119S–127S. [Google Scholar] [CrossRef]
- Bansal, A.; Henao-Mejia, J.; Simmons, R.A. Immune system: An emerging player in mediating effects of endocrine disruptors on metabolic health. Endocrinology 2018, 159, 32–45. [Google Scholar] [CrossRef]
- Hayden, M.R.; Chowdhury, N.; Govindarajan, G.; Karuparthi, P.R.; Habibi, J.; Sowers, J.R. Myocardial myocyte remodeling and fibrosis in the cardiometabolic syndrome. J. Cardiometab. Syndr. 2006, 1, 326–333. [Google Scholar] [CrossRef]
- Guo, R.; Rogers, O.; Nair, S. Targeting apelinergic system in cardiometabolic disease. Curr. Drug Targets 2017, 18, 1785–1791. [Google Scholar] [CrossRef]
- Newgard, C.B. Metabolomics and metabolic diseases: Where do we stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef]
- Njock, M.S.; Fish, J.E. Endothelial miRNAs as cellular messengers in cardiometabolic diseases. Trends Endocrinol. Metab. 2017, 28, 237–246. [Google Scholar] [CrossRef]
- Rotllan, N.; Price, N.; Pati, P.; Goedeke, L.; Fernández-Hernando, C. microRNAs in lipoprotein metabolism and cardiometabolic disorders. Atherosclerosis 2016, 246, 352–360. [Google Scholar] [CrossRef]
- Assmann, T.S.; Milagro, F.I.; Martínez, J.A. Crosstalk between microRNAs, the putative target genes and the lncRNA network in metabolic diseases. Mol. Med. Rep. 2019, 20, 3543–3554. [Google Scholar] [CrossRef] [PubMed]
- Dihazi, H.; Asif, A.R.; Beißbarth, T.; Bohrer, R.; Feussner, K.; Feussner, I.; Jahn, O.; Lenz, C.; Majcherczyk, A.; Schmidt, B.; et al. Integrative omics—From data to biology. Expert Rev. Proteomics 2018, 15, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems biology and multi-omics integration: Viewpoints from the metabolomics research community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.F.; Allayee, H.; Gerszten, R.E.; Ideraabdullah, F.; Kris-Etherton, P.M.; Ordovás, J.M.; Rimm, E.B.; Wang, T.J.; Bennett, B.J. American heart association council on functional genomics and translational biology, council on epidemiology and prevention, and stroke council. nutrigenomics, the microbiome, and gene-environment interactions: New directions in cardiovascular disease research, prevention, and treatment: A scientific statement from the American Heart Association. Circ. Cardiovasc. Genet. 2016, 9, 291–313. [Google Scholar] [PubMed]
- Taleb, S. Tryptophan dietary impacts gut barrier and metabolic diseases. Front. Immunol. 2019, 10, 2113. [Google Scholar] [CrossRef]
- The Health Consequences of Smoking: 50 Years of Progress: A Report of the Surgeon General. US Department of Health and Human Services, Centers for Disease Control and Prevention: Atlanta, GA, USA. Available online: http://www.surgeongeneral.gov/library/reports/50-years-of-progress/index.html (accessed on 31 October 2014).
- Sotos-Prieto, M.; Mattei, J. Mediterranean diet and cardiometabolic diseases in racial/ethnic minority populations in the United States. Nutrients 2018, 10, 352. [Google Scholar] [CrossRef]
- Balistreri, C.R. Anti-inflamm-ageing and/or anti-age-related disease emerging treatments: A historical alchemy or revolutionary effective procedures? Mediat. Inflamm. 2018, 2018, 3705389. [Google Scholar] [CrossRef]
- Galbete, C.; Schwingshackl, L.; Schwedhelm, C.; Boeing, H.; Schulze, M.B. Evaluating Mediterranean diet and risk of chronic disease in cohort studies: An umbrella review of meta-analyses. Eur. J. Epidemiol. 2018, 33, 909–931. [Google Scholar] [CrossRef]
- Dinu, M.; Pagliai, G.; Casini, A.; Sofi, F. Mediterranean diet and multiple health outcomes: An umbrella review of meta-analyses of observational studies and randomised trials. Eur. J. Clin. Nutr. 2018, 72, 30–43. [Google Scholar] [CrossRef]
- Peters, R.; Ee, N.; Peters, J.; Beckett, N.; Booth, A.; Rockwood, K.; Anstey, K.J. Common risk factors for major noncommunicable disease, a systematic overview of reviews and commentary: The implied potential for targeted risk reduction. Ther. Adv. Chronic Dis. 2019, 10, 2040622319880392. [Google Scholar] [CrossRef]
- Franquesa, M.; Pujol-Busquets, G.; García-Fernández, E.; Rico, L.; Shamirian-Pulido, L.; Aguilar-Martínez, A.; Medina, F.X.; Serra-Majem, L.; Bach-Faig, A. Mediterranean diet and cardiodiabesity: A systematic review through evidence-based answers to key clinical questions. Nutrients 2019, 11, 655. [Google Scholar] [CrossRef] [PubMed]
- Martini, D. Health benefits of Mediterranean diet. Nutrients. 2019, 11, 1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.H.; Kitai, T.; Hazen, S.L. Gut microbiota in cardiovascular health and disease. Circ. Res. 2017, 31, 1183–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasselman, L.J.; Vernice, N.A.; DeLeon, J.; Reiss, A.B. The gut microbiome and elevated cardiovascular risk in obesity and autoimmunity. Atherosclerosis 2018, 271, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Rotroff, D.M.; Motsinger-Reif, A.A. Embracing integrative multiomics approaches. Int. J. Genom. 2016, 2016, 1715985. [Google Scholar] [CrossRef] [Green Version]
- Veldhuizen, J.; Migrino, R.Q.; Nikkhah, M. Three-dimensional microengineered models of human cardiac diseases. J. Biol. Eng. 2019, 13, 29. [Google Scholar] [CrossRef] [Green Version]
- Park, S.B.; Lee, S.Y.; Jung, W.H.; Lee, J.; Jeong, H.G.; Hong, J.; Kang, D.; Kim, K.Y. Development of in vitro three-dimensional co-culture system for metabolic syndrome therapeutic agents. Diabetes Obes. Metab. 2019, 21, 1146–1157. [Google Scholar] [CrossRef]
- Zaragoza, C.; Gomez-Guerrero, C.; Martin-Ventura, J.L.; Blanco-Colio, L.; Lavin, B.; Mallavia, B.; Tarin, C.; Mas, S.; Ortiz, A.; Egido, J. Animal models of cardiovascular diseases. J. Biomed. Biotechnol. 2011, 2011, 497841. [Google Scholar] [CrossRef]
- Von Scheidt, M.; Zhao, Y.; Kurt, Z.; Pan, C.; Zeng, L.; Yang, X.; Schunkert, H.; Lusis, A.J. Applications and limitations of mouse models for understanding human atherosclerosis. Cell Metab. 2017, 25, 248–261. [Google Scholar] [CrossRef] [Green Version]
- Riehle, C.; Bauersachs, J. Of mice and men: Models and mechanisms of diabetic cardiomyopathy. Basic Res. Cardiol. 2018, 114, 2. [Google Scholar] [CrossRef] [Green Version]
- Améen, C.; Strehl, R.; Björquist, P.; Lindahl, A.; Hyllner, J.; Sartipy, P. Human embryonic stem cells: Current technologies and emerging industrial applications. Crit. Rev. Oncol. Hematol. 2008, 65, 54–80. [Google Scholar] [CrossRef] [PubMed]
- Sayed, N.; Wu, J.C. Towards cardio-precision medicine. Eur. Heart J. 2017, 38, 1014–1016. [Google Scholar] [CrossRef] [PubMed]
- Balistreri, C.R.; Garagnani, P.; Madonna, R.; Vaiserman, A.; Melino, G. Developmental programming of adult haematopoiesis system. Ageing Res. Rev. 2019, 54, 100918. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Mead, B.E.; Safaee, H.; Langer, R.; Karp, J.M.; Levy, O. Engineering stem cell organoids. Cell Stem Cell. 2016, 18, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.N.; Ingber, D.E. Microfluidic organs-on-chips. Nat. Biotechnol. 2014, 32, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Miri, A.K.; Mostafavi, E.; Khorsandi, D.; Hu, S.K.; Malpica, M.; Khademhosseini, A. Bioprinters for organs-on-chips. Biofabrication 2019, 11, 042002. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Choudhury, D. Microfluidic bioprinting for organ-on-a-chip models. Drug Discov. Today 2019, 24, 1248–1257. [Google Scholar] [CrossRef]
- Maschmeyer, I.; Lorenz, A.K.; Schimek, K.; Hasenberg, T.; Ramme, A.P.; Hübner, J.; Lindner, M.; Drewell, C.; Bauer, S.; Thomas, A.; et al. A four-organ-chip for interconnected long-term co-culture of human intestine, liver, skin and kidney equivalents. Lab Chip 2015, 15, 2688–2699. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shi, J.; Lu, H.S.; Zhang, H. Functional genomics and CRISPR applied to cardiovascular research and medicine. Arterioscler. Thromb. Vasc. Biol. 2019, 39, e188–e194. [Google Scholar] [CrossRef]
- Da Silva Xavier, G.; Hodson, D.J. Mouse models of peripheral metabolic disease. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 299–315. [Google Scholar] [CrossRef]
- Ben Jehuda, R.; Shemer, Y.; Binah, O. Genome editing in induced pluripotent stem cells using CRISPR/Cas9. Stem Cell Rev. Rep. 2018, 14, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, G.; Liu, M.L. Microvesicles as emerging biomarkers and therapeutic targets in cardiometabolic diseases. Genomics Proteomics Bioinform. 2018, 16, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Gerszten, R.E. Toward new biomarkers of cardiometabolic diseases. Cell Metab. 2013, 18, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siomkajło, M.; Daroszewski, J. Branched chain amino acids: Passive biomarkers or the key to the pathogenesis of cardiometabolic diseases? Adv. Clin. Exp. Med. 2019. [Google Scholar] [CrossRef]
- Krzystek-Korpacka, M.; Wiśniewski, J.; Fleszar, M.G.; Bednarz-Misa, I.; Bronowicka-Szydełko, A.; Gacka, M.; Masłowski, L.; Kędzior, K.; Witkiewicz, W.; Gamian, A. Metabolites of the nitric oxide (NO) pathway are altered and indicative of reduced NO and arginine bioavailability in patients with cardiometabolic diseases complicated with chronic wounds of lower extremities: Targeted metabolomics approach (LC-MS/MS). Oxid. Med. Cell Longev. 2019, 2019, 5965721. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.Y.; Cheng, X.L.; Lin, R.C. Lipidomics applications for discovering biomarkers of diseases in clinical chemistry. Int. Rev. Cell Mol. Biol. 2014, 313, 1–26. [Google Scholar] [CrossRef]
- Stevens, K.G.; Bader, C.A.; Sorvina, A.; Brooks, D.A.; Plush, S.E.; Morrison, J.L. Imaging and lipidomics methods for lipid analysis in metabolic and cardiovascular disease. J. Dev. Orig. Health Dis. 2017, 8, 566–574. [Google Scholar] [CrossRef]
- Heianza, Y.; Ma, W.; Manson, J.E.; Rexrode, K.M.; Qi, L. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: A systematic review and meta-analysis of prospective studies. J. Am. Heart Assoc. 2017, 6, e004947. [Google Scholar] [CrossRef]
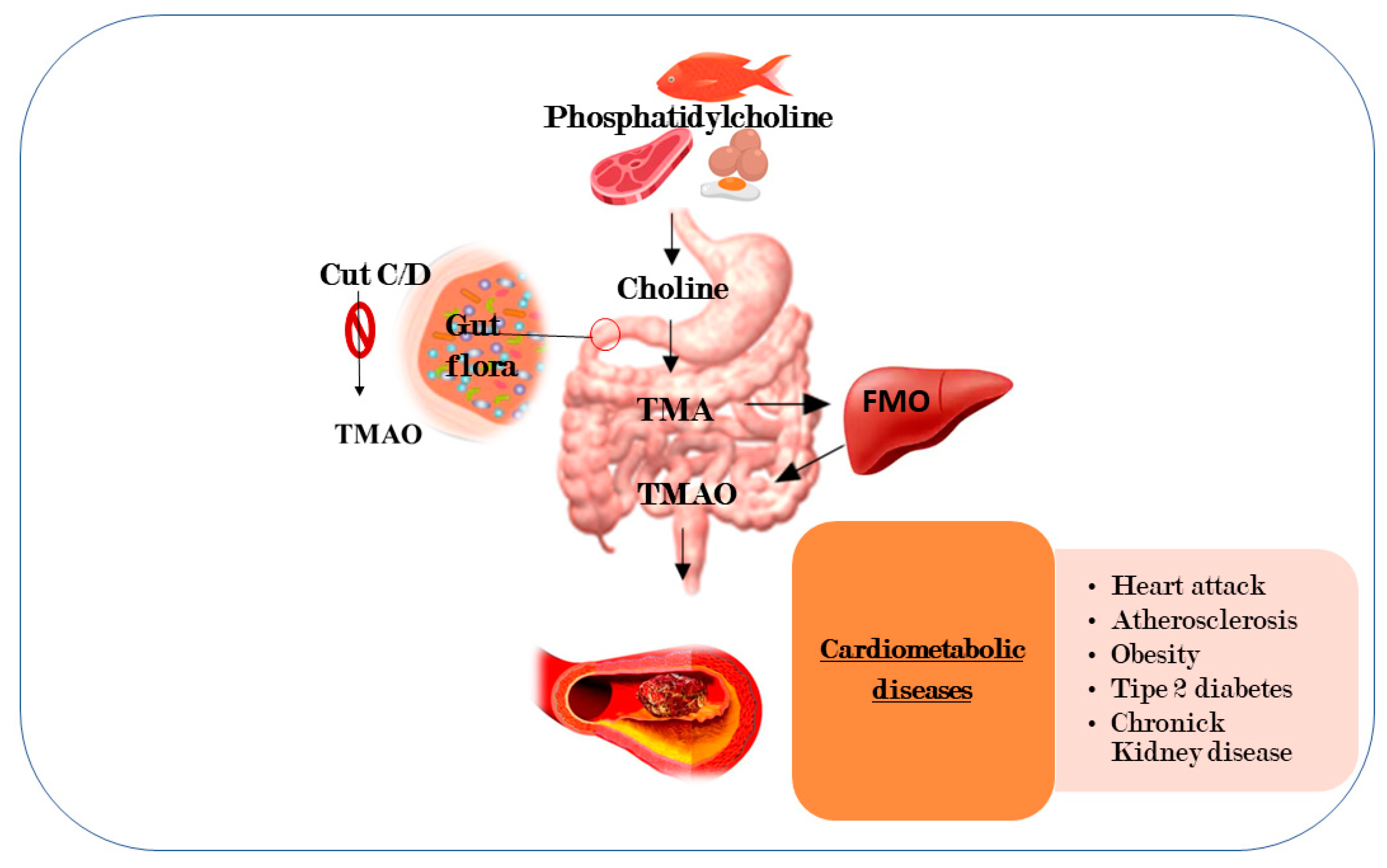
- Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; Wu, Y.; Schauer, P.; Smith, J.D.; Allayee, H.; Tang, W.H.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar]
- Tang, W.H.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Dou, P.; Gao, M.; Kong, X.; Li, C.; Liu, Z.; Huang, T. Assessment of causal direction between gut microbiota-dependent metabolites and cardiometabolic health: A bidirectional mendelian randomization analysis. Diabetes 2019, 68, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, R.; Ge, X.; Han, L.; Yu, P.; Gong, X.; Meng, Q.; Zhang, Y.; Fan, H.; Zheng, L.; Liu, Z.; et al. Gut microbe-generated metabolite trimethylamine N-oxide and the risk of diabetes: A systematic review and dose-response meta-analysis. Obes. Rev. 2019, 20, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Descamps, H.C.; Herrmann, B.; Wiredu, D.; Thaiss, C.A. The path toward using microbial metabolites as therapies. EBioMedicine 2019, 44, 747–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estruch, R.; Ros, E.; Salas-Salvado, J.; Covas, M.I.; Corella, D.; Aros, F.; Gomez-Gracia, E.; Ruiz-Gutierrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a mediterranean diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Leon-Mimila, P.; Wang, J.; Huertas-Vazquez, A. Relevance of multi-omics studies in cardiovascular diseases. Front Cardiovasc. Med. 2019, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Balistreri, C.R.; Candore, G.; Accardi, G.; Bova, M.; Buffa, S.; Bulati, M.; Forte, G.I.; Listì, F.; Martorana, A.; Palmeri, M.; et al. Genetics of longevity. Data from the studies on Sicilian centenarians. Immun. Ageing 2012, 9, 8. [Google Scholar] [CrossRef]
- Candore, G.; Balistreri, C.R.; Colonna-Romano, G.; Lio, D.; Listì, F.; Vasto, S.; Caruso, C. Gender-related immune-inflammatory factors, age-related diseases, and longevity. Rejuvenation Res. 2010, 13, 292–297. [Google Scholar] [CrossRef] [Green Version]
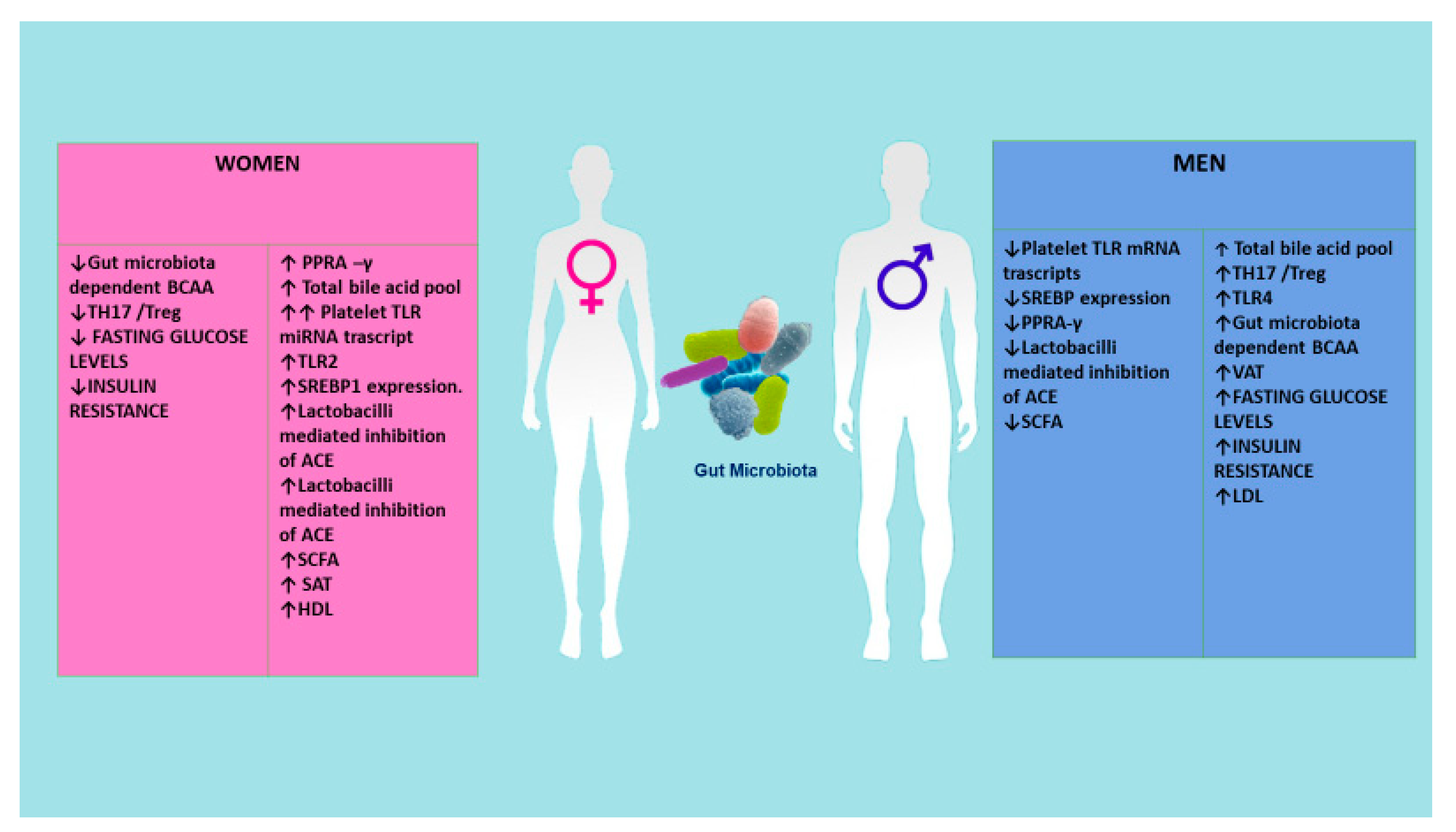
- Gerdts, E.; Regitz-Zagrosek, V. Sex differences in cardiometabolic disorders. Nat. Med. 2019, 25, 1657–1666. [Google Scholar] [CrossRef]
- Kuznetsova, T. Sex differences in epidemiology of cardiac and vascular disease. Adv. Exp. Med. Biol. 2018, 1065, 61–70. [Google Scholar] [CrossRef]
- Chella Krishnan, K.; Mehrabian, M.; Lusis, A.J. Sex differences in metabolism and cardiometabolic disorders. Curr. Opin. Lipidol. 2018, 29, 404–410. [Google Scholar] [CrossRef]
- Madonna, R.; Balistreri, C.R.; De Rosa, S.; Muscoli, S.; Selvaggio, S.; Selvaggio, G.; Ferdinandy, P.; De Caterina, R. Impact of sex differences and diabetes on coronary atherosclerosis and ischemic heart disease. J. Clin. Med. 2019, 8, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razavi, A.C.; Potts, K.S.; Kelly, T.N.; Bazzano, L.A. Sex, gut microbiome, and cardiovascular disease risk. Biol. Sex Differ. 2019, 10, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannenbaum, C.; Ellis, R.P.; Eyssel, F.; Zou, J.; Schiebinger, L. Sex and gender analysis improves science and engineering. Nature 2019, 575, 137–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.Y. Observational studies: A review of study designs, challenges an strategies to reduce confounding. Int. J. Clin. Pract. 2009, 63, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Wilke, I.; Borosch, J.; Pecha, S.; Papmeyer, S.; Behrens, S.; von Kodolitsch, Y.; Aydin, A. Age-dependent clinical and echocardiographic manifestations of aortic stenosis in an unselected, non-biased cohort. Cardiovasc. Diagn. Ther. 2019, 9 (Suppl. 2), S238–S246. [Google Scholar] [CrossRef]
- Balistreri, C.R.; Colonna-Romano, G.; Lio, D.; Candore, G.; Caruso, C. TLR4 polymorphisms and ageing: Implications for the pathophysiology of age-related diseases. J Clin Immunol. 2009, 29, 406–415. [Google Scholar] [CrossRef]
- De Diego, I.; Peleg, S.; Fuchs, B. The role of lipids in aging-related metabolic changes. Chem Phys Lipids. 2019, 222, 59–69. [Google Scholar] [CrossRef]
- Kildey, K.; Rooks, K.; Weier, S.; Flower, R.L.; Dean, M.M. Effect of age, gender and mannose-binding lectin (MBL) status on the inflammatory profile in peripheral blood plasma of Australian blood donors. Hum. Immunol. 2014, 75, 973–979. [Google Scholar] [CrossRef]
- Cevenini, E.; Caruso, C.; Candore, G.; Capri, M.; Nuzzo, D.; Duro, G.; Rizzo, C.; Colonna-Romano, G.; Lio, D.; Di Carlo, D.; et al. Age-related inflammation: The contribution of different organs, tissues and systems. How to face it for therapeutic approaches. Curr. Pharm. Des. 2010, 16, 609–618. [Google Scholar] [CrossRef]
- Böhm, M.; Schumacher, H.; Laufs, U.; Sleight, P.; Schmieder, R.; Unger, T.; Teo, K.; Yusuf, S. Effects of nonpersistence with medication on outcomes in high-risk patients with cardiovascular disease. Am. Heart J. 2013, 166, 306–314. [Google Scholar] [CrossRef]
- Dieterlen, M.T.; John, K.; Haase, S.; Garbade, J.; Tarnok, A.; Mohr, F.W.; Bittner, H.B.; Barten, M.J. Effect of confounding factors on a phospho-flow assay of ribosomal S6 protein for therapeutic drug monitoring of the mTOR-inhibitor everolimus in heart transplanted patients. Biomarkers 2017, 22, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Lafeber, M.; Spiering, W.; van der Graaf, Y.; Nathoe, H.; Bots, M.L.; Grobbee, D.E.; Visseren, F.L. The combined use of aspirin, a statin, and blood pressure-lowering agents (polypill components) and the risk of vascular morbidity and mortality in patients with coronary artery disease. Am. Heart J. 2013, 166, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Howards, P.P. An overview of confounding. Part 2: How to identify it and special situations. Acta Obstet. Gynecol. Scand. 2018, 97, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Howards, P.P. An overview of confounding. Part 1: The concept and how to address it. Acta Obstet. Gynecol. Scand. 2018, 97, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Iraci, N.; Leonardi, T.; Gessler, F.; Vega, B.; Pluchino, S. Focus on Extracellular Vesicles: Physiological Role and Signalling Properties of Extracellular Membrane Vesicles. Int. J. Mol. Sci. 2016, 17, 171. [Google Scholar] [CrossRef]
- Bodega, G.; Alique, M.; Puebla, L.; Carracedo, J.; Ramírez, R.M. Microvesicles: ROS scavengers and ROS producers. J. Extracell. Vesicles 2019, 8, 1626654. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, V.C.; Timmers, L.; Schoneveld, A.H.; Wang, J.W.; van de Weg, S.M.; Sze, S.K.; van Keulen, J.K.; Hoes, A.W.; den Ruijter, H.M.; de Kleijn, D.P.; et al. Serum extracellular vesicle protein levels are associated with acute coronary syndrome. Eur. Heart J. Acute Cardiovasc. Care 2013, 2, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Jeanneteau, J.; Hibert, P.; Martinez, M.C.; Tual-Chalot, S.; Tamareille, S.; Furber, A.; Andriantsitohaina, R.; Prunier, F. Microparticle release in remote ischemic conditioning mechanism. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H871–H877. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.; Sorensson, P.; Saleh, N.; Arheden, H.; Ryden, L.; Pernow, J. Circulating endothelial and platelet derived microparticles reflect the size of myocardium at risk in patients with ST-elevation myocardial infarction. Atherosclerosis 2012, 221, 226–231. [Google Scholar] [CrossRef]
- Borges, F.T.; Melo, S.A.; Özdemir, B.C.; Kato, N.; Revuelta, I.; Miller, C.A.; Gattone, V.H.; LeBleu, V.S.; Kalluri, R. TGF-b1-containing exosomes from injured epithelial cells activate fibroblasts to initiate tissue regenerative responses and fibrosis. J. Am. Soc. Nephrol. 2013, 24, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Sabatier, F.; Darmon, P.; Hugel, B.; Combes, V.; Sanmarco, M.; Velut, J.G.; Arnoux, D.; Charpiot, P.; Freyssinet, J.M.; Oliver, C.; et al. Type 1 and type 2 diabetic patients display different patterns of cellular microparticles. Diabetes 2002, 51, 2840–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, H.; Sugiyama, S.; Kugiyama, K.; Watanabe, K.; Fukushima, H.; Tanaka, T.; Sakamoto, T.; Yoshimura, M.; Jinnouchi, H.; Ogawa, H. Elevated levels of VE-cadherin-positive endothelial microparticles in patients with type 2 diabetes mellitus and coronary artery disease. J. Am. Coll. Cardiol. 2005, 45, 1622–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, N.; Imaizumi, M.; Nomura, S.; Shozu, A.; Arichi, M.; Matsuoka, M.; Matsumura, M. Increased levels of platelet-derived microparticles in patients with diabetic retinopathy. Diabetes Res. Clin. Pract. 2005, 68, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Tsimerman, G.; Roguin, A.; Bachar, A.; Melamed, E.; Brenner, B.; Aharon, A. Involvement of microparticles in diabetic vascular complications. Thromb. Haemost. 2011, 106, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Valencia, K.; Lecanda, F. Microvesicles: Isolation, characterization for in vitro and in vivo procedures. Methods Mol. Biol. 2016, 1372, 181–192. [Google Scholar] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scola, L.; Giarratana, R.M.; Torre, S.; Argano, V.; Lio, D.; Balistreri, C.R. On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches. Int. J. Mol. Sci. 2019, 20, 6015. https://doi.org/10.3390/ijms20236015
Scola L, Giarratana RM, Torre S, Argano V, Lio D, Balistreri CR. On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches. International Journal of Molecular Sciences. 2019; 20(23):6015. https://doi.org/10.3390/ijms20236015
Chicago/Turabian StyleScola, Letizia, Rosa Maria Giarratana, Salvatore Torre, Vincenzo Argano, Domenico Lio, and Carmela Rita Balistreri. 2019. "On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches" International Journal of Molecular Sciences 20, no. 23: 6015. https://doi.org/10.3390/ijms20236015
APA StyleScola, L., Giarratana, R. M., Torre, S., Argano, V., Lio, D., & Balistreri, C. R. (2019). On the Road to Accurate Biomarkers for Cardiometabolic Diseases by Integrating Precision and Gender Medicine Approaches. International Journal of Molecular Sciences, 20(23), 6015. https://doi.org/10.3390/ijms20236015