The Effect of Hydroxamic Siderophores Structure on Acetylation of Histone H3 and Alpha Tubulin in Pinus sylvestris Root Cells

 , ,
, ,
Abstract
:1. Introduction
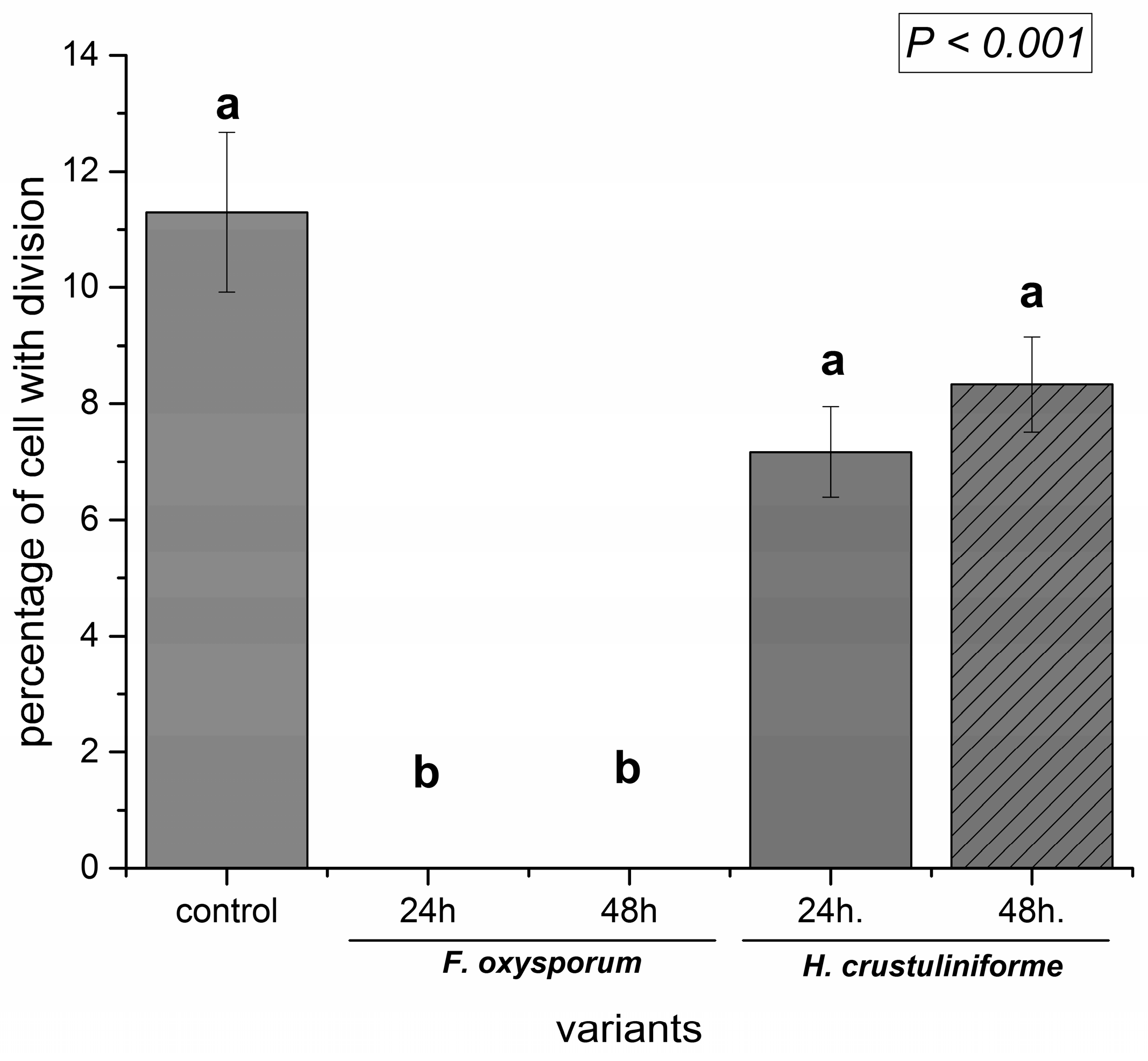
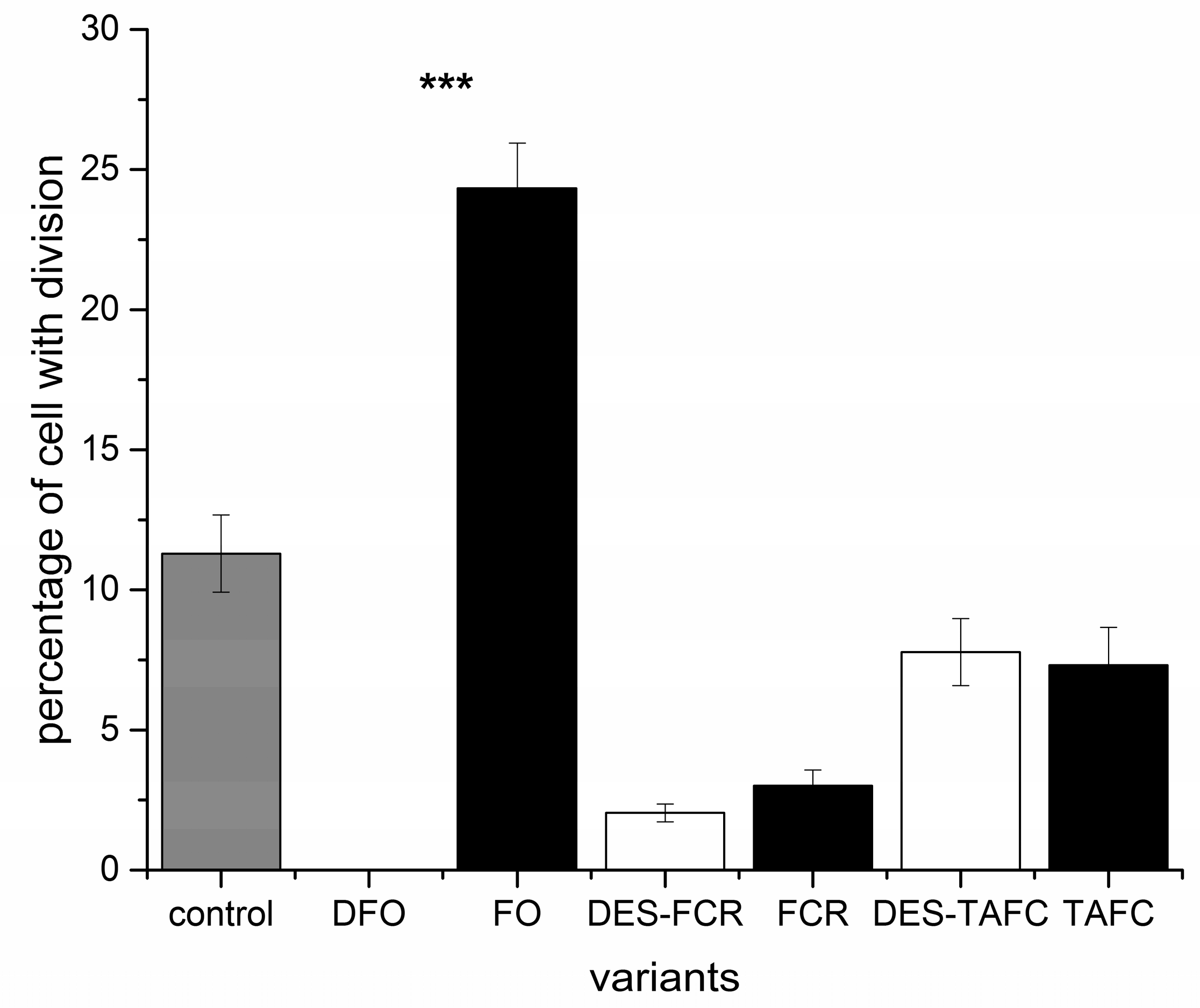
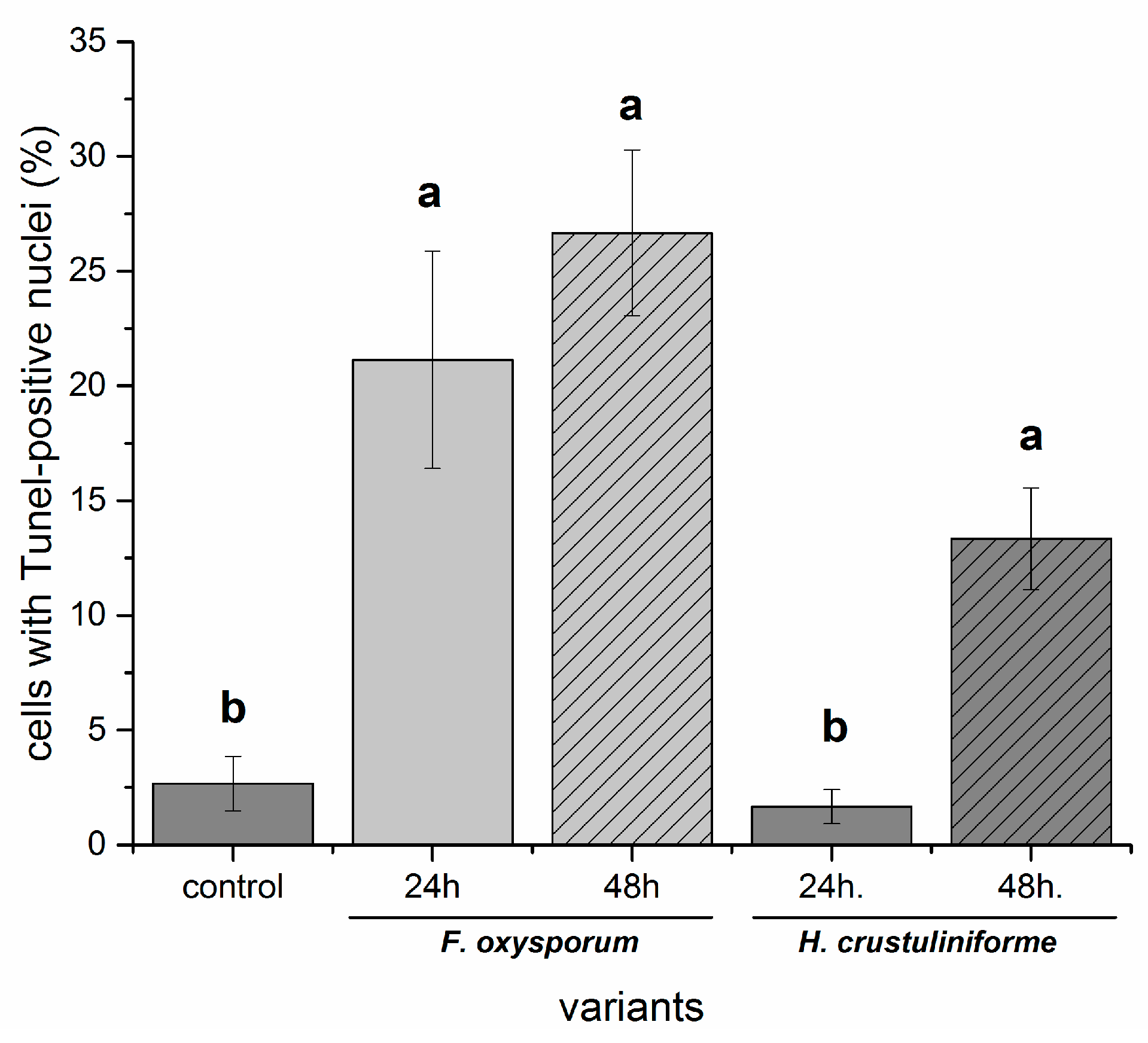
2. Results
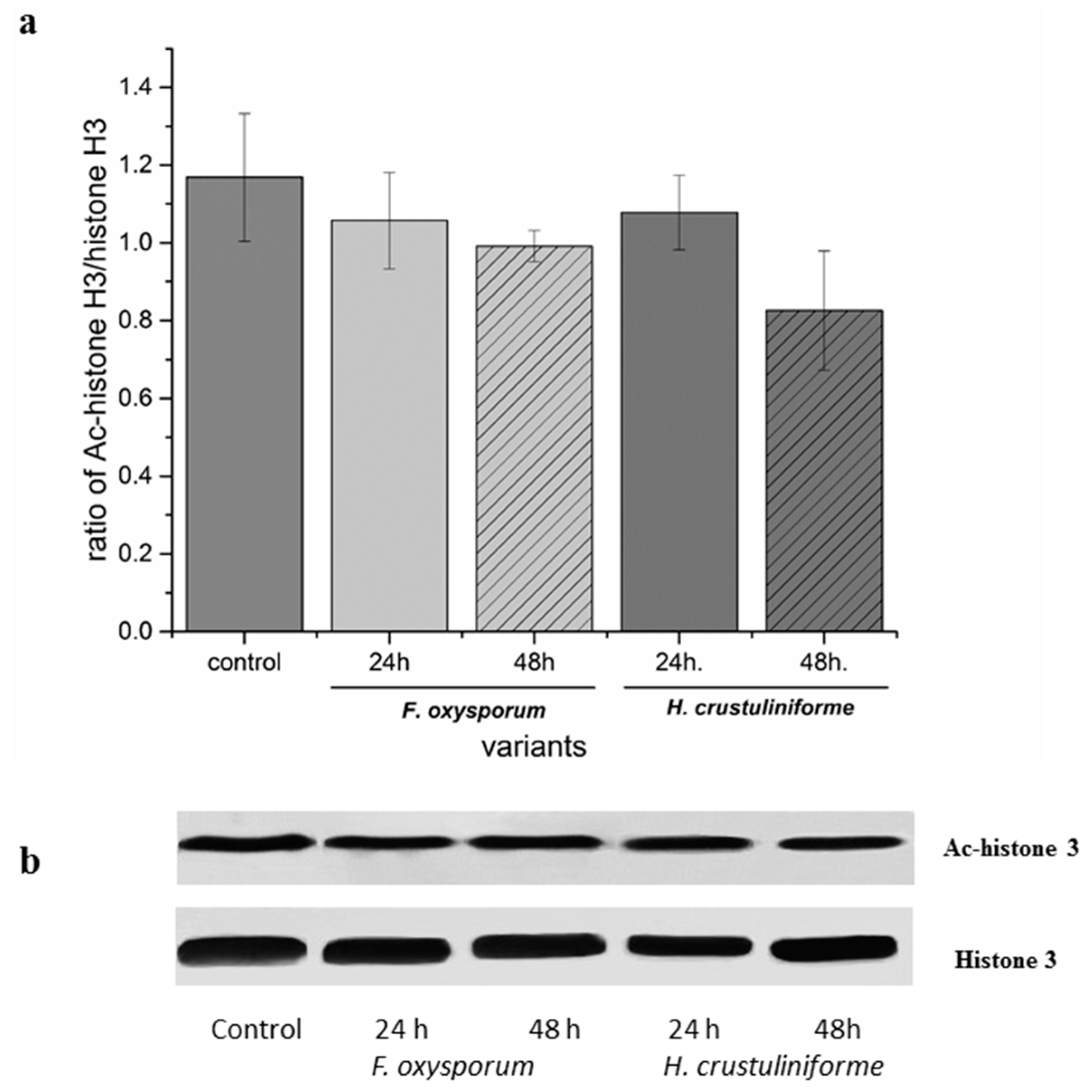
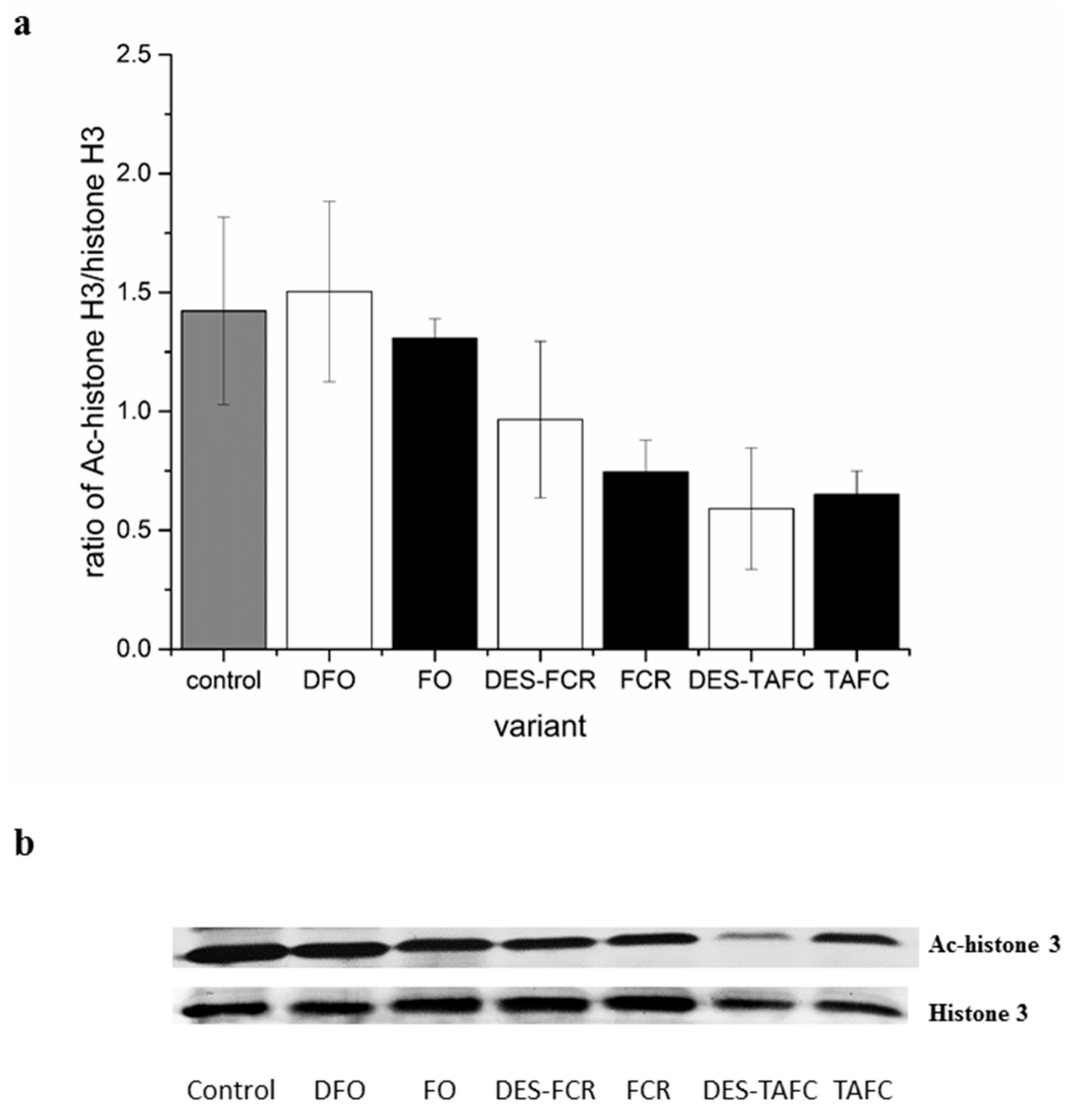
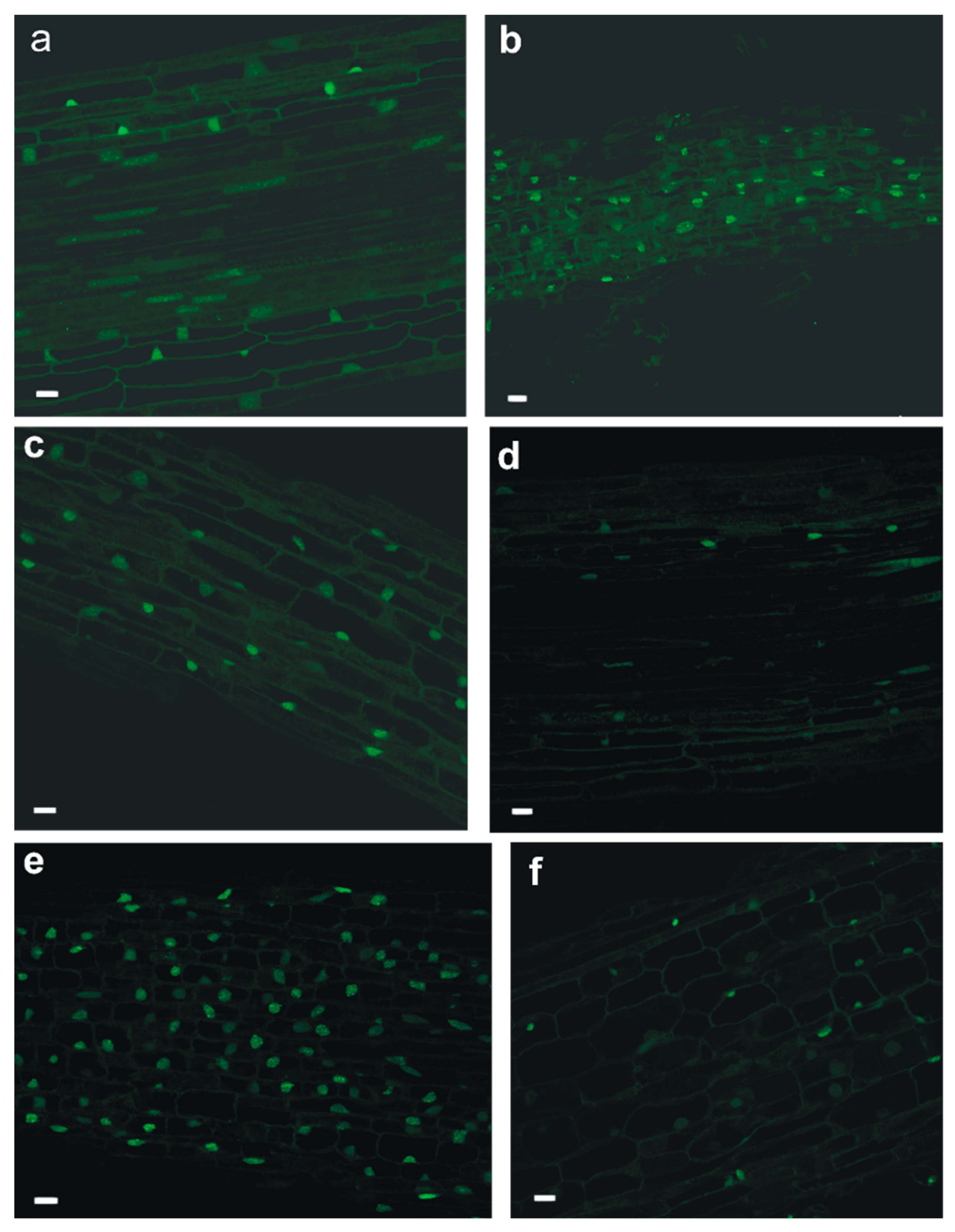
2.1. Assessment of Microtubule and Histone Acetylation
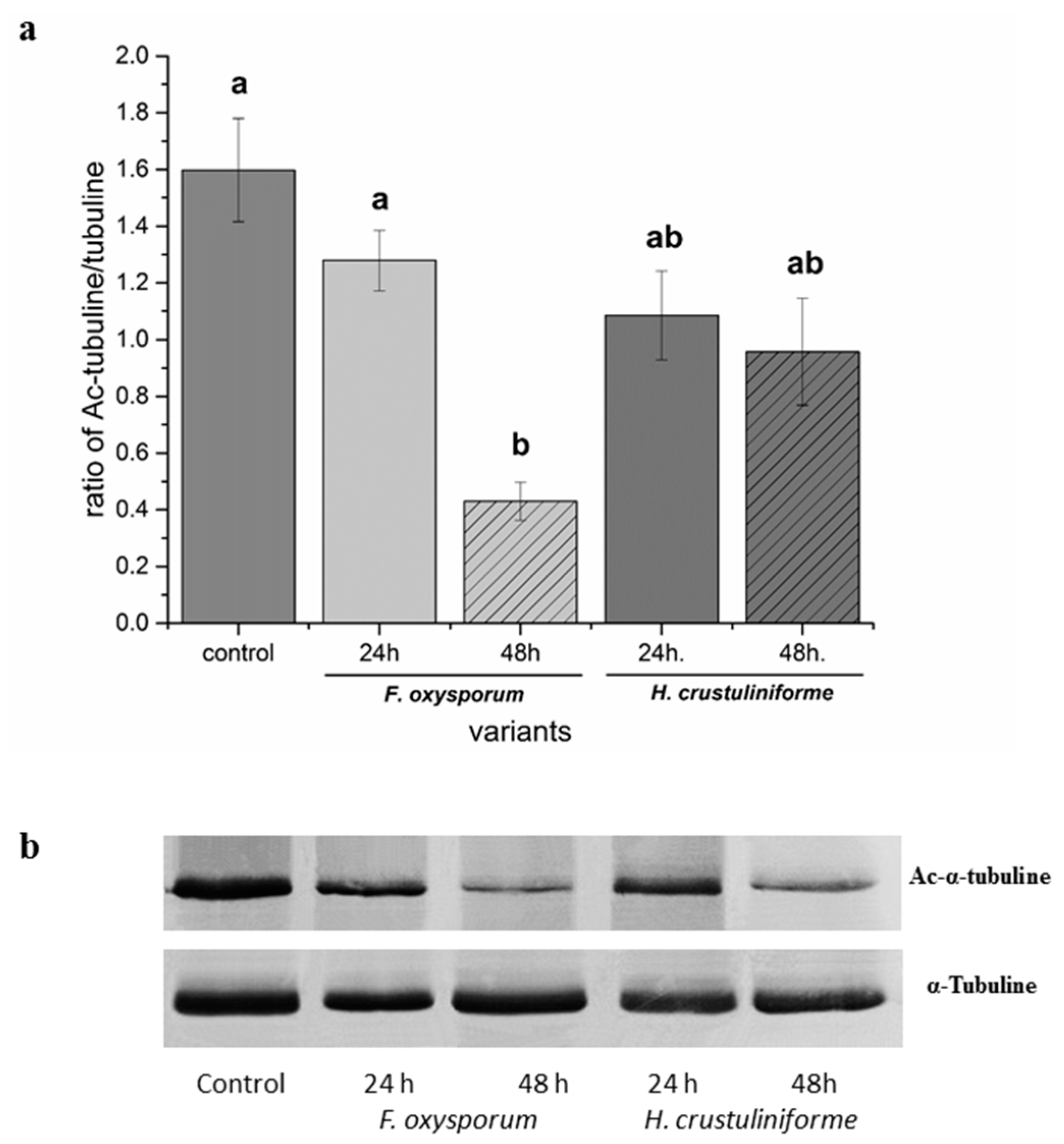
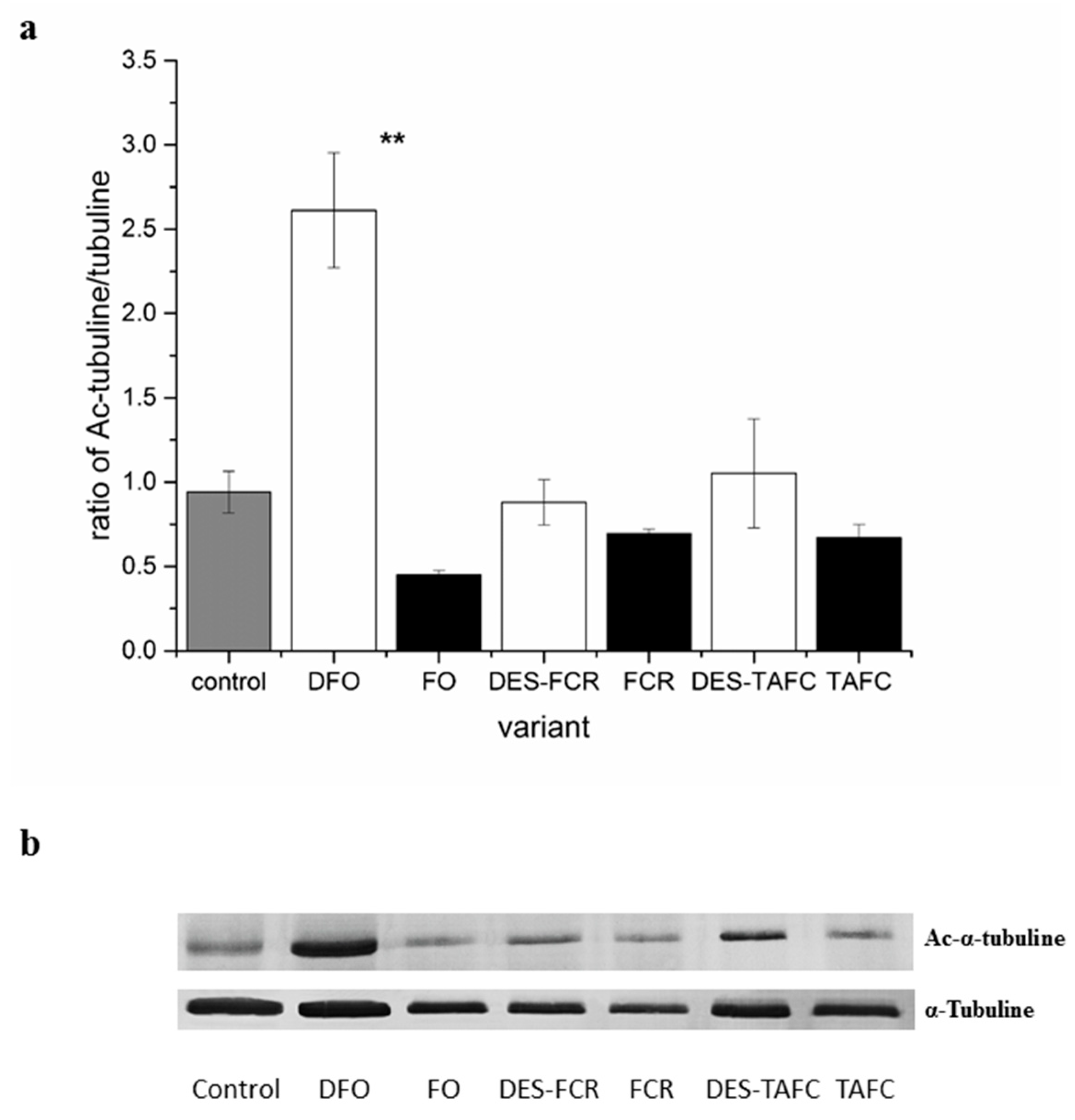
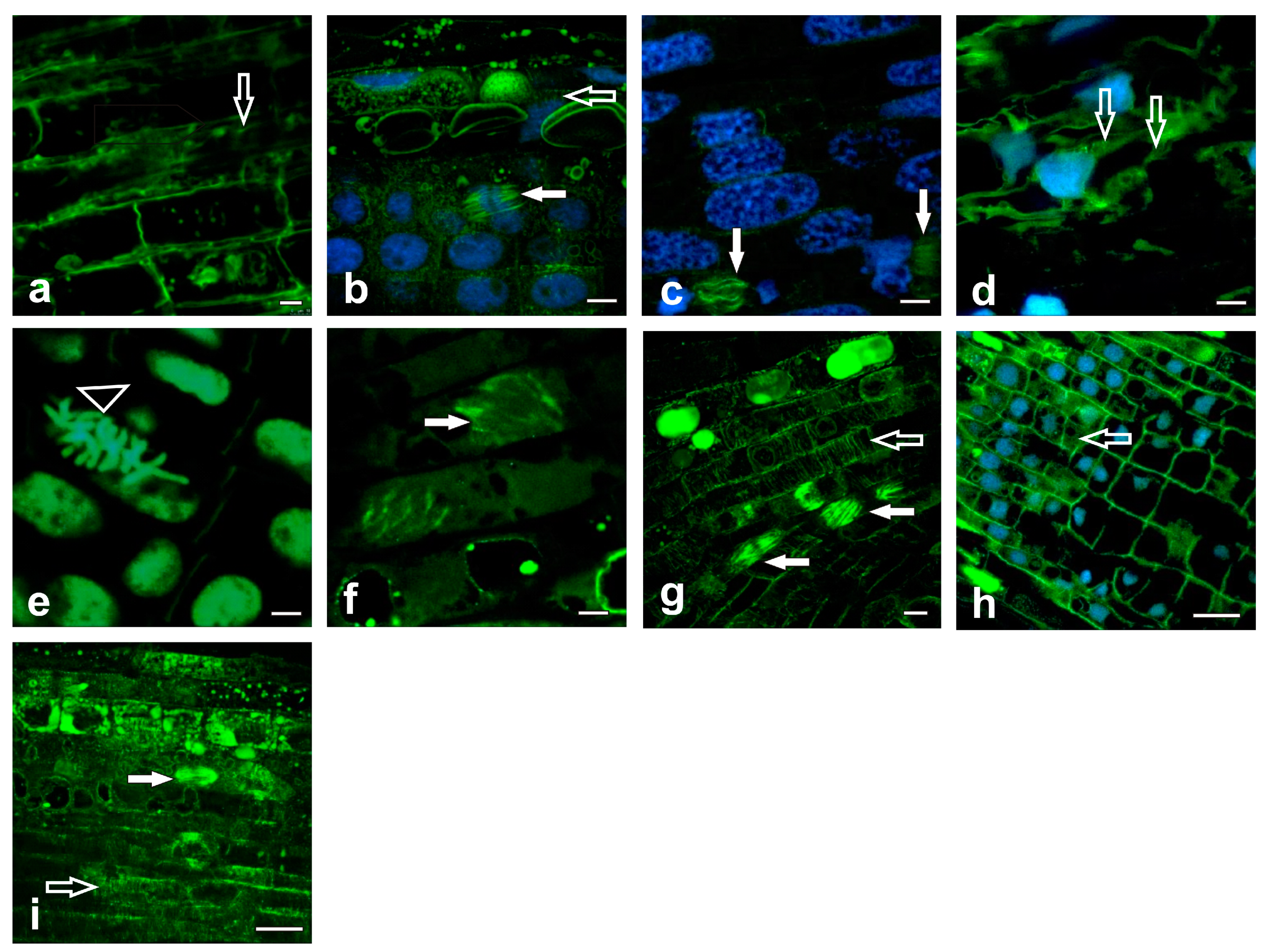
2.2. Analysis of the Tubulin Cytoskeleton
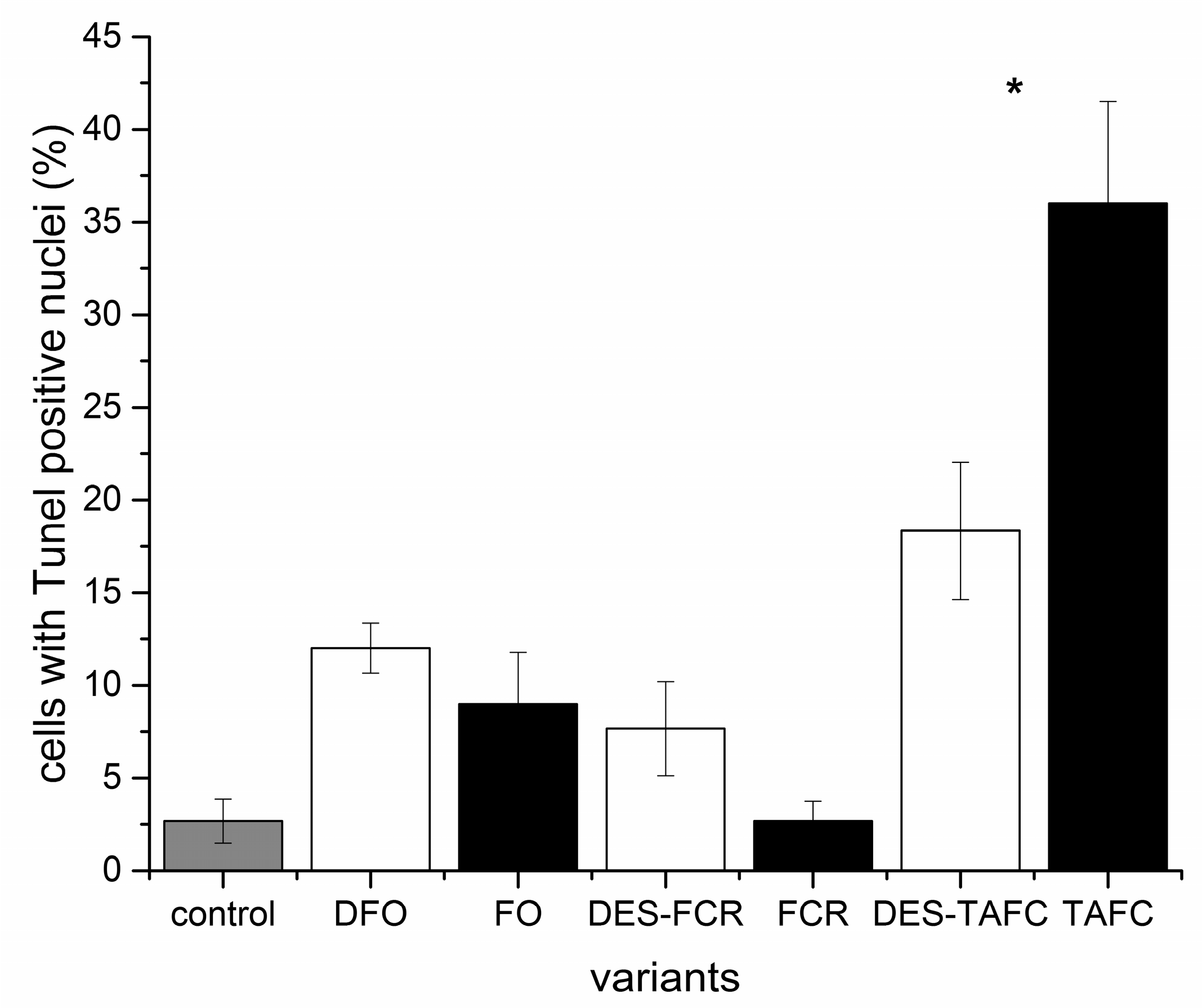
2.3. Cell Death Assessment (Nuclear DNA Fragmentation)
3. Discussion
4. Materials and Methods
4.1. Organisms and Growth Conditions
4.2. Assessment of the Acetylation of Microtubules and Histones
4.3. Analysis of Tubulin Cytoskeleton
4.4. Assessment of Cell Death (Nuclear DNA Fragmentation)
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aznar, A.; Chen, N.W.; Rigault, M.; Riache, N.; Joseph, D.; Desmaële, D.; Mouille, G.; Boutet, S.; Soubigou-Taconnat, L.; Renou, J.P.; et al. Scavenging iron: A novel mechanism of plant immunity activation by microbial siderophores. Plant Physiol. 2014, 164, 2167–2183. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Greenshields, D.L.; Sammynaiken, R.; Hirji, R.N.; Selvaraj, G.; Wei, Y. Targeted alterations in iron homeostasis underlie plant defense responses. J. Cell Sci. 2007, 120, 596–605. [Google Scholar] [CrossRef] [Green Version]
- Oide, S.; Moeder, W.; Krasnoff, S.; Gibson, D.; Haas, H.; Yoshioka, K.; Turgeon, B.G. NPS6, encoding a nonribosomal peptide synthetase involved in siderophore-mediated iron metabolism, is a conserved virulence determinant of plant pathogenic ascomycetes. Plant Cell 2006, 18, 2836–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucha, J.; Budka, A.; Kayzer, D.; Guzicka, M.; Łakomy, P.; Zadworny, M. The origin of reactive oxygen during interaction of Pinus sylvestris root and Heterobasidion annosum s.l.–the linkage with the iron. Eur. J. Plant Pathol. 2015, 143, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Mucha, J.; Guzicka, M.; Łakomy, P.; Zadworny, M. Iron and reactive oxygen responses in Pinus sylvestris root cortical cells infected with different species of Heterobasidion annosum sensu lato. Planta 2012, 236, 975–988. [Google Scholar] [CrossRef] [PubMed]
- Mucha, J.; Guzicka, M.; Łakomy, P.; Zadworny, M. Production of metal-chelating compounds by species of Heterobasidion annosum sensu lato. J. Phytopathol. 2013, 161, 791–799. [Google Scholar] [CrossRef]
- Mucha, J.; Guzicka, M.; Lakomy, P.; Zadworny, M. Accumulation of iron-binding compounds in root of Pinus sylvestris challenged by Heterobasidion annosum sensu lato. Dendrobiology 2015, 73. [Google Scholar] [CrossRef] [Green Version]
- Mucha, J.; Napierała-Filipiak, A.; Gabała, E.; Pawłowski, T.A.; Zadworny, M. Redistribution of iron and hydrogen peroxide in Pinus sylvestris roots in response to trophically diverse fungi. Eur. J. Plant Pathol. 2019, 153, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.J.; Koulman, A.; Christensen, M.; Lane, G.A.; Fraser, K.; Forester, N.; Johnson, R.D.; Bryan, G.T.; Rasmussen, S. An extracellular siderophore is required to maintain the mutualistic interaction of Epichloë festucae with Lolium perenne. PLoS Pathog. 2013, 9, e1003332. [Google Scholar] [CrossRef] [Green Version]
- De Vleesschauwer, D.; Djavaheri, M.; Bakker, P.A.H.M.; Höfte, M. Pseudomonas fluorescens WCS374r-induced systemic resistance in rice against Magnaporthe oryzae is based on pseudobactin-mediated priming for a salicylic acid-repressible multifaceted defense response. Plant Physiol. 2008, 148, 1996–2012. [Google Scholar] [CrossRef] [Green Version]
- Meziane, H.; van der Sluis, I.; van Loon, L.C.; Höfte, M.; Bakker, P.A. Determinants of Pseudomonas putida WCS358 involved in inducing systemic resistance in plants. Mol. Plant Pathol. 2005, 6, 177–185. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.C.; Bakker, P.A.; van der Heijdt, W.H.; Wendehenne, D.; Pugin, A. Early responses of tobacco suspension cells to rhizobacterial elicitors of induced systemic resistance. Mol. Plant Microbe Interact. 2008, 21, 1609–1621. [Google Scholar] [CrossRef] [Green Version]
- Mucha, J.; Gabała, E.; Zadworny, M. The effects of structurally different siderophores on the organelles of Pinus sylvestris root cells. Planta 2019, 249, 1747–1760. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (cu, Zn, Mn, Fe, Ni, Mo, B, cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Mucha, J.; Guzicka, M.; Ratajczak, E.; Zadworny, M. Strategies utilized by trophically diverse fungal species for Pinus sylvestris root colonization. Tree Physiol. 2014, 34, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Vorontsova, M.; Shaw, P.; Reader, S.; Moore, G. Effect of 5-azacytidine and trichostatin A on somatic centromere association in wheat. Genome 2004, 47, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Lechner, T.; Lusser, A.; Brosch, G.; Eberharter, A.; Goralik-Schramel, M.; Loidl, P. A comparative study of histone deacetylases of plant, fungal and vertebrate cells. BBA Protein Struct. Mol. Enzymol. 1996, 1296, 181–188. [Google Scholar] [CrossRef]
- Gantt, S.L.; Gattis, S.G.; Fierke, C.A. Catalytic activity and inhibition of human histone deacetylase 8 is dependent on the identity of the active site metal ion. Biochemistry 2006, 45, 6170–6178. [Google Scholar] [CrossRef]
- Choi, S.M.; Song, H.R.; Han, S.K.; Han, M.; Kim, C.Y.; Park, J.; Lee, Y.H.; Jeon, J.S.; Noh, Y.S.; Noh, B. HDA19 is required for the repression of salicylic acid biosynthesis and salicylic acid-mediated defense responses in Arabidopsis. Plant J. 2012, 71, 135–146. [Google Scholar] [CrossRef]
- Cuperlovic-Culf, M.; Culf, A. Protein acetylation as an integral part of metabolism in cancer development and progression. Am. J. Cancer Rev. 2014, 2, 6–28. [Google Scholar]
- Kim, K.C.; Lai, Z.; Fan, B.; Chen, Z. Arabidopsis WRKY38 and WRKY62 transcription factors interact with histone deacetylase 19 in basal defence. Plant Cell 2008, 20, 2357–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Zhang, L.; Duan, J.; Miki, B.; Wu, K. HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling of pathogen response in Arabidopsis. Plant Cell 2005, 17, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ransom, R.F.; Walton, J.D. Histone hyperacetylation in maize in response to treatment with HC-toxin or infection by the filamentous fungus Cochliobolus carbonum. Plant Physiol. 1997, 115, 1021–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, G.; Walley, J.W. Dynamic protein acetylation in plant–pathogen interactions. Front. Plant Sci. 2016, 7, 421. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant immunity: From signaling to epigenetic control of defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Walley, J.W.; Shen, Z.; McReynolds, M.R.; Schmelz, E.A.; Briggs, S.P. Fungal-induced protein hyperacetylation in maize identified by acetylome profiling. Proc. Natl. Acad. Sci. USA 2018, 115, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Finkemeier, I.; Laxa, M.; Miguet, L.; Howden, A.J.; Sweetlove, L.J. Proteins of diverse function and subcellular location are lysine acetylated in Arabidopsis. Plant Physiol. 2011, 155, 1779–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.T.; Nimick, M.; Uhrig, G.; Templeton, G.; Morrice, N.; Gourlay, R.; Delong, A.; Moorhead, G.B. Arabidopsis thaliana histone deacetylase 14 (Hda14) is an alpha-tubulin deacetylase that associates with Pp2a and enriches in the microtubule fraction with the putative histone acetyltransferase Elp3. Plant J. 2012, 71, 263–272. [Google Scholar] [CrossRef]
- Wu, X.; Oh, M.H.; Schwarz, E.M.; Larue, C.T.; Sivaguru, M.; Imai, B.S.; Yau, P.M.; Ort, D.R.; Huber, S.C. Lysine acetylation is a widespread protein modification for diverse proteins in Arabidopsis. Plant Physiol 2011, 155, 1769–1778. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Lv, S.; Zhang, C.; Yang, C. Histone deacetylases and their functions in plants. Plant Cell Rep. 2013, 32, 465–478. [Google Scholar] [CrossRef]
- Cahill, D.; Rookes, J.; Michalczyk, A.; McDonald, K.; Drake, A. Microtubule dynamics in compatible and incompatible interactions of soybean hypocotyl cells with Phytophthora sojae. Plant Pathol. 2002, 51, 629–640. [Google Scholar] [CrossRef]
- Dellagi, A.; Segond, D.; Rigault, M.; Fagard, M.; Simon, C.; Saindrenan, P.; Expert, D. Microbial siderophores exert a subtle role in Arabidopsis during infection by manipulating the immune response and the iron status. Plant Physiol. 2009, 150, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Zadworny, M.; Guzicka, M.; Łakomy, P.; Samardakiewicz, S.; Smoliński, D.J.; Mucha, J. Analysis of microtubule and microfilament distribution in Pinus sylvestris roots following infection by Heterobasidion species. Forest Pathol. 2013, 43, 222–231. [Google Scholar] [CrossRef]
- Trapet, P.; Avoscan, L.; Klinguer, A.; Pateyron, S.; Citerne, S.; Chervin, C.; Mazurier, S.; Lemanceau, P.; Wendehenne, D.; Besson-Bard, A. The Pseudomonas fluorescens siderophore pyoverdine weakens Arabidopsis thaliana defense in favor of growth in iron-deficient conditions. Plant Physiol. 2016, 171, 675–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.H.; Hurley, B.; Felsensteiner, C.; Yea, C.; Ckurshumova, W.; Bartetzko, V.; Wang, P.W.; Quach, V.; Lewis, J.D.; Liu, Y.C.; et al. A bacterial acetyltransferase destroys plant microtubule networks and blocks secretion. PLoS Pathog. 2012, 8, e1002523. [Google Scholar] [CrossRef] [Green Version]
- Gilmer, S.; Clay, P.; MacRae, T.H.; Fowke, L.C. Acetylated tubulin is found in all microtubule arrays of two species of pine. Protoplasma 1999, 207, 174–185. [Google Scholar] [CrossRef]
- Nam, H.J.; Kang, J.K.; Kim, S.K.; Ahn, K.J.; Seok, H.; Park, S.J.; Chang, J.S.; Pothoulakis, C.; Lamont, J.T.; Kim, H. Clostridium difficile toxin A decreases acetylation of tubulin, leading to microtubule depolymerization through activation of histone deacetylase 6, and this mediates acute inflammation. J. Biol. Chem. 2010, 285, 32888–32896. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Durán, R.; Zipfel, C. Trade-off between growth and immunity: Role of brassinosteroids. Trends Plant Sci. 2015, 20, 12–19. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, Z.; Zhang, Y.; Yong, S.; Salas-Burgos, A.; Koomen, J.; Olashaw, N.; Parsons, J.T.; Yang, X.J.; Dent, S.R.; et al. HDAC6 modulates cell motility by altering the acetylation level of cortactin. Mol. Cell 2007, 27, 197–213. [Google Scholar]
- Schmelzer, E. Cell polarization, a crucial process in fungal defence. Trends Plant Sci. 2002, 7, 411–415. [Google Scholar] [CrossRef]
- Cheng, Q.; Balzer, E.; Miller, S.M.; Yoshida, M.; Wong, J.C. Effect of histone deacetylase inhibitors on tubulin acetylation and development in Volvox carteri (Volvocales) 1. J. Phycol. 2006, 42, 417–422. [Google Scholar] [CrossRef]
- Piperno, G.; LeDizet, M.; Chang, X.-J. Microtubules containing acetylated ~-tubulin in mammalian cells in culture. J. Cell Biol. 1987, 104, 289–302. [Google Scholar] [CrossRef]
- Bánréti, Á.; Sass, M.; Graba, Y. The emerging role of acetylation in the regulation of autophagy. Autophagy 2013, 9, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Ingestad, T. Mineral nutrient requirements of Pinus sylvestris and Picea abies seedlings. Physiol Plantarum 1979, 45, 373–380. [Google Scholar] [CrossRef]
- Staszak, A.M.; Pawłowski, T.A. Forest tree research in post genomic era. Introduction to systems biology of broadleaves. Dendrobiology 2012, 68, 113–123. [Google Scholar]
- Staszak, A.M.; Guzicka, M.; Pawłowski, T.A. Signalling regulators of abscisic and gibberellic acid pathways are involved in dormancy breaking of Norway maple (Acer platanoides L.) seeds. Acta Physiol Plant 2017, 39, 251. [Google Scholar] [CrossRef] [Green Version]
- Staszak, A.M.; Rewers, M.; Sliwinska, E.; Klupczyńska, E.A.; Pawłowski, T.A. DNA synthesis pattern, proteome, and ABA and GA signalling in developing seeds of Norway maple (Acer platanoides). Funct. Plant Biol. 2019, 46, 152–164. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fischer, R.; Timberlake, W.E. Aspergillus nidulans apsA (anucleate primary sterigmata) encodes a coiled-coil protein required for nuclear positioning and completion of asexual development. J. Cell Biol. 1995, 128, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Vitha, S.; Baluška, F.; Mews, M.; Volkmann, D. Immunofluorescence detection of F-actin on low melting point wax sections from plant tissues. J. Histochem. Cytochem. 1997, 45, 89–95. [Google Scholar] [CrossRef] [Green Version]
- De Vleesschauwer, D.; Höfte, M. Bacterial determinants and host defense responses underpinning rhizobacteria-mediated systemic resistance in rice. In Advances in Genetics, Genomics and Control of Rice Blast Disease; Wang, G.L., Valent, B., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 191–206. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variable | df | acH3/H3 | acTub/Tub | Percentage of Cell Division | Cell Death | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| Siderophores ability to bind iron (DES) | 1 | 0.36 | 0.559 | 30.22 | 0.001 | 28.99 | 0.001 | 0.001 | 0.975 |
| Siderophores compounds (SIDER) | 2 | 5.55 | 0.020 | 8.15 | 0.006 | 95.59 | 0.001 | 22.77 | 0.001 |
| SIDER×DES | 2 | 0.21 | 0.817 | 14.44 | 0.001 | 98.11 | 0.001 | 6.42 | 0.003 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mucha, J.; Pawłowski, T.A.; Klupczyńska, E.A.; Guzicka, M.; Zadworny, M. The Effect of Hydroxamic Siderophores Structure on Acetylation of Histone H3 and Alpha Tubulin in Pinus sylvestris Root Cells. Int. J. Mol. Sci. 2019, 20, 6099. https://doi.org/10.3390/ijms20236099
Mucha J, Pawłowski TA, Klupczyńska EA, Guzicka M, Zadworny M. The Effect of Hydroxamic Siderophores Structure on Acetylation of Histone H3 and Alpha Tubulin in Pinus sylvestris Root Cells. International Journal of Molecular Sciences. 2019; 20(23):6099. https://doi.org/10.3390/ijms20236099
Chicago/Turabian StyleMucha, Joanna, Tomasz A. Pawłowski, Ewelina A. Klupczyńska, Marzenna Guzicka, and Marcin Zadworny. 2019. "The Effect of Hydroxamic Siderophores Structure on Acetylation of Histone H3 and Alpha Tubulin in Pinus sylvestris Root Cells" International Journal of Molecular Sciences 20, no. 23: 6099. https://doi.org/10.3390/ijms20236099
APA StyleMucha, J., Pawłowski, T. A., Klupczyńska, E. A., Guzicka, M., & Zadworny, M. (2019). The Effect of Hydroxamic Siderophores Structure on Acetylation of Histone H3 and Alpha Tubulin in Pinus sylvestris Root Cells. International Journal of Molecular Sciences, 20(23), 6099. https://doi.org/10.3390/ijms20236099