Preventive Effects of Pyungwi-san against Dextran Sulfate Sodium- and Clostridium difficile-Induced Inflammatory Bowel Disease in Mice


Abstract
:1. Introduction
2. Results
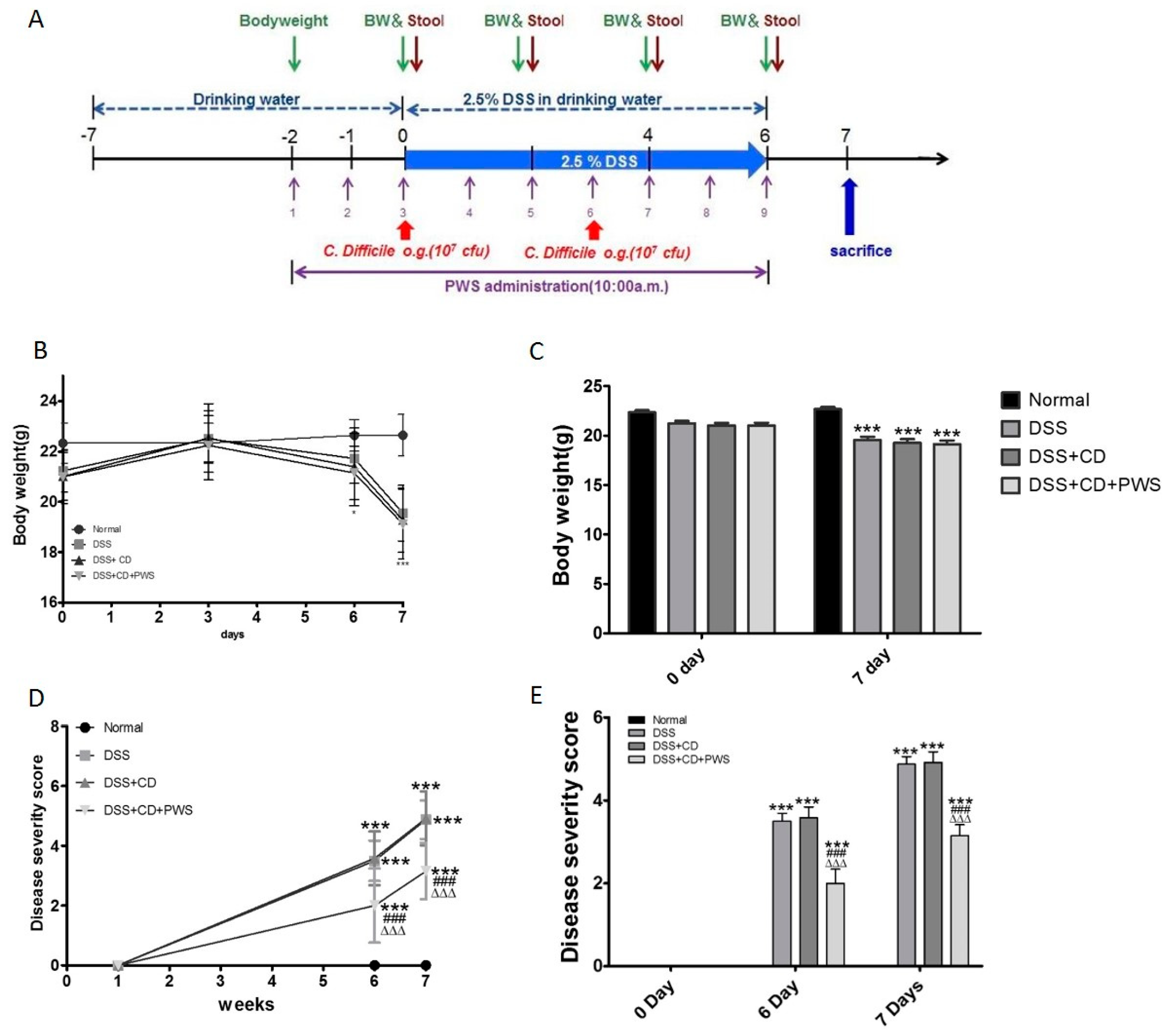
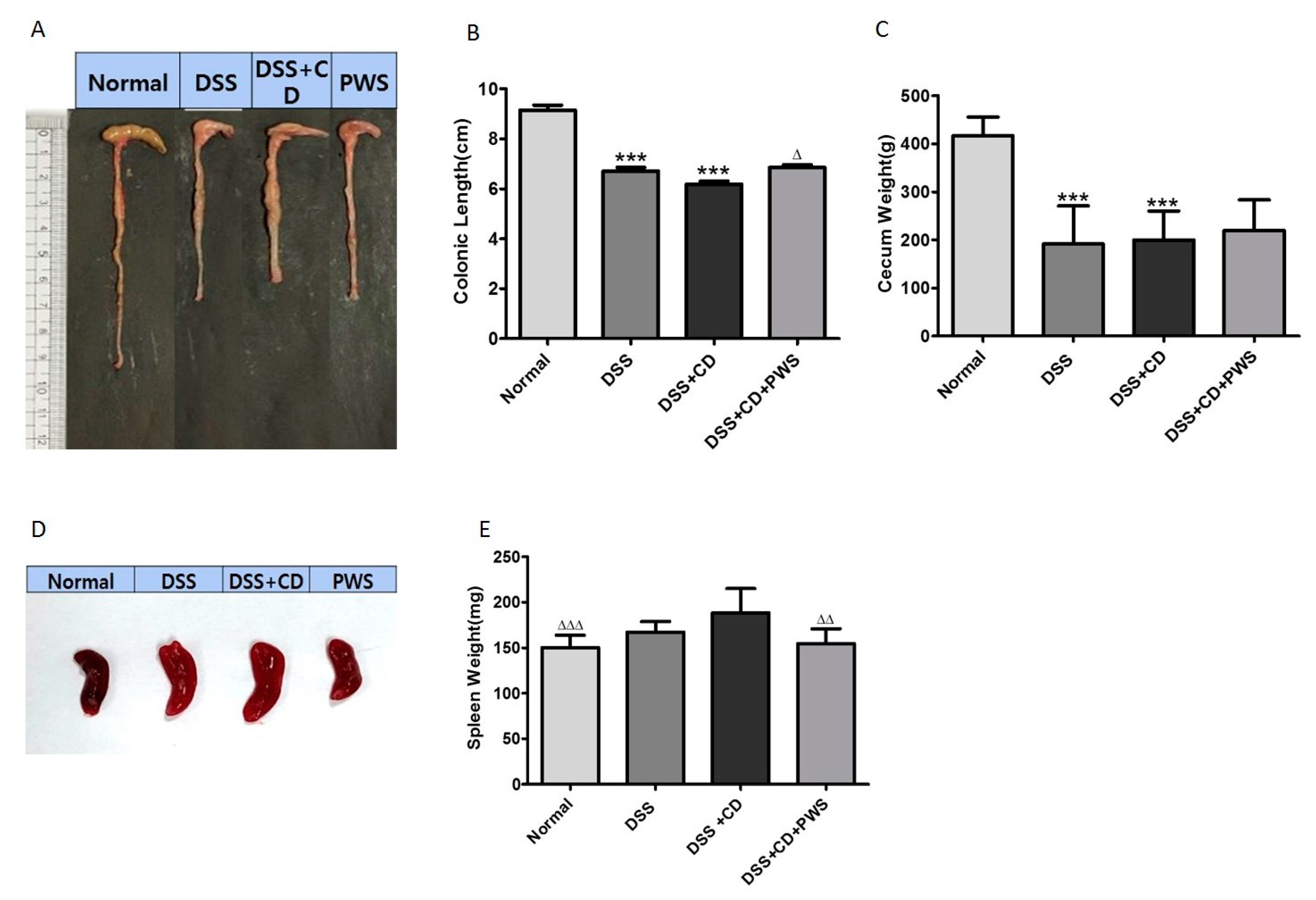
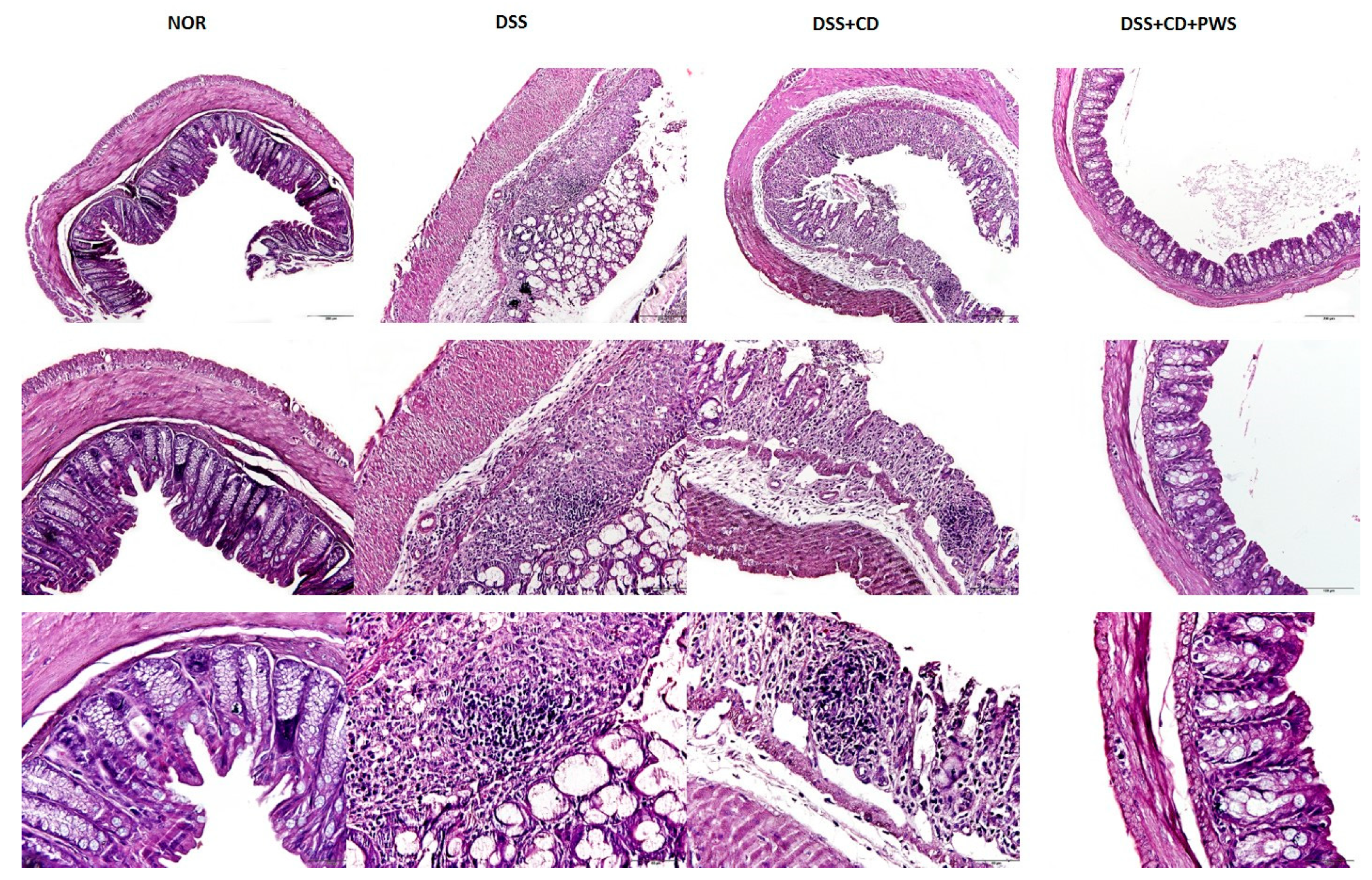
2.1. The Effect of PWS Treatment on Clinical and Tissue Parameters in DSS and DSS+ C. difficile-Induced Colitis
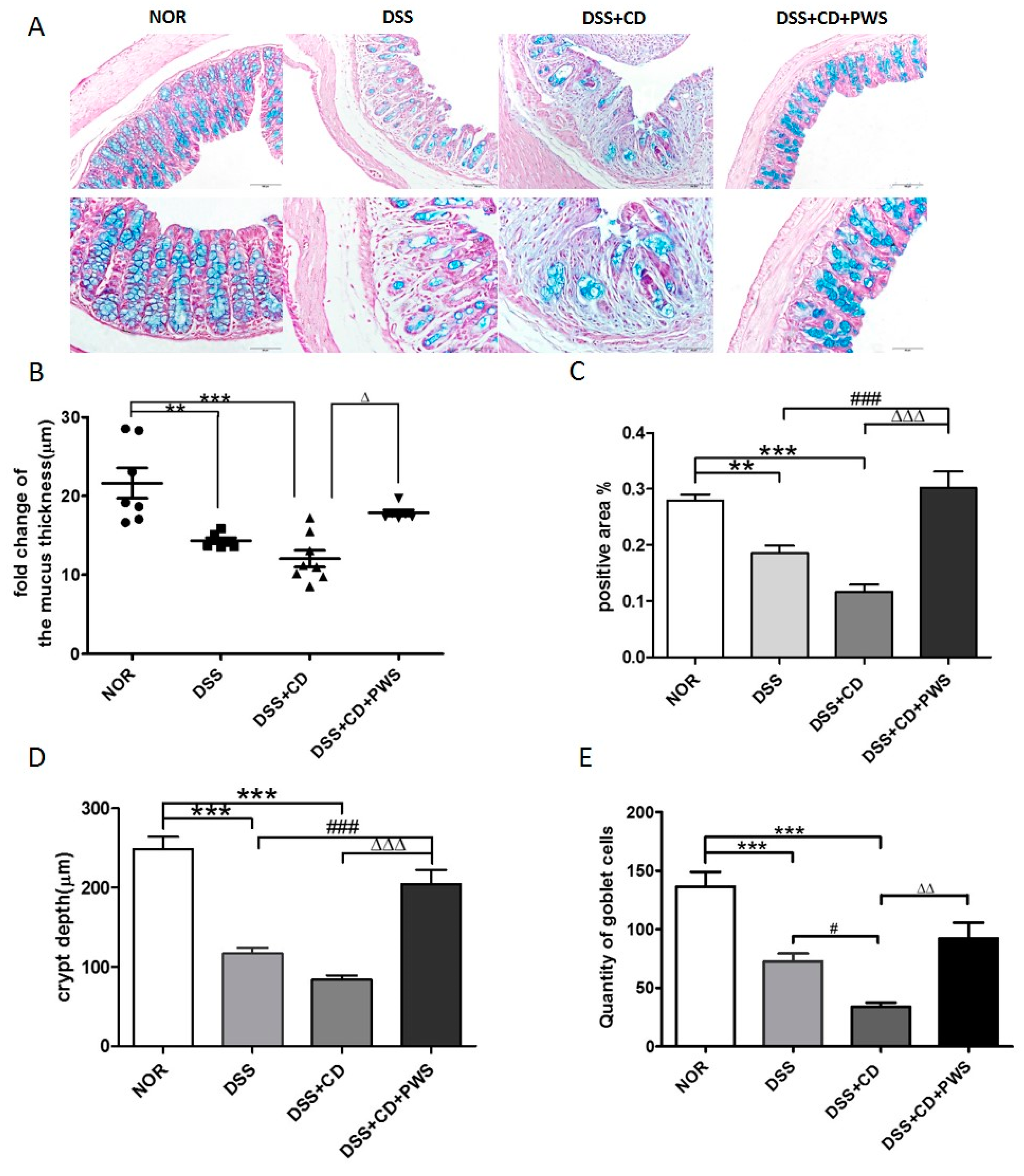
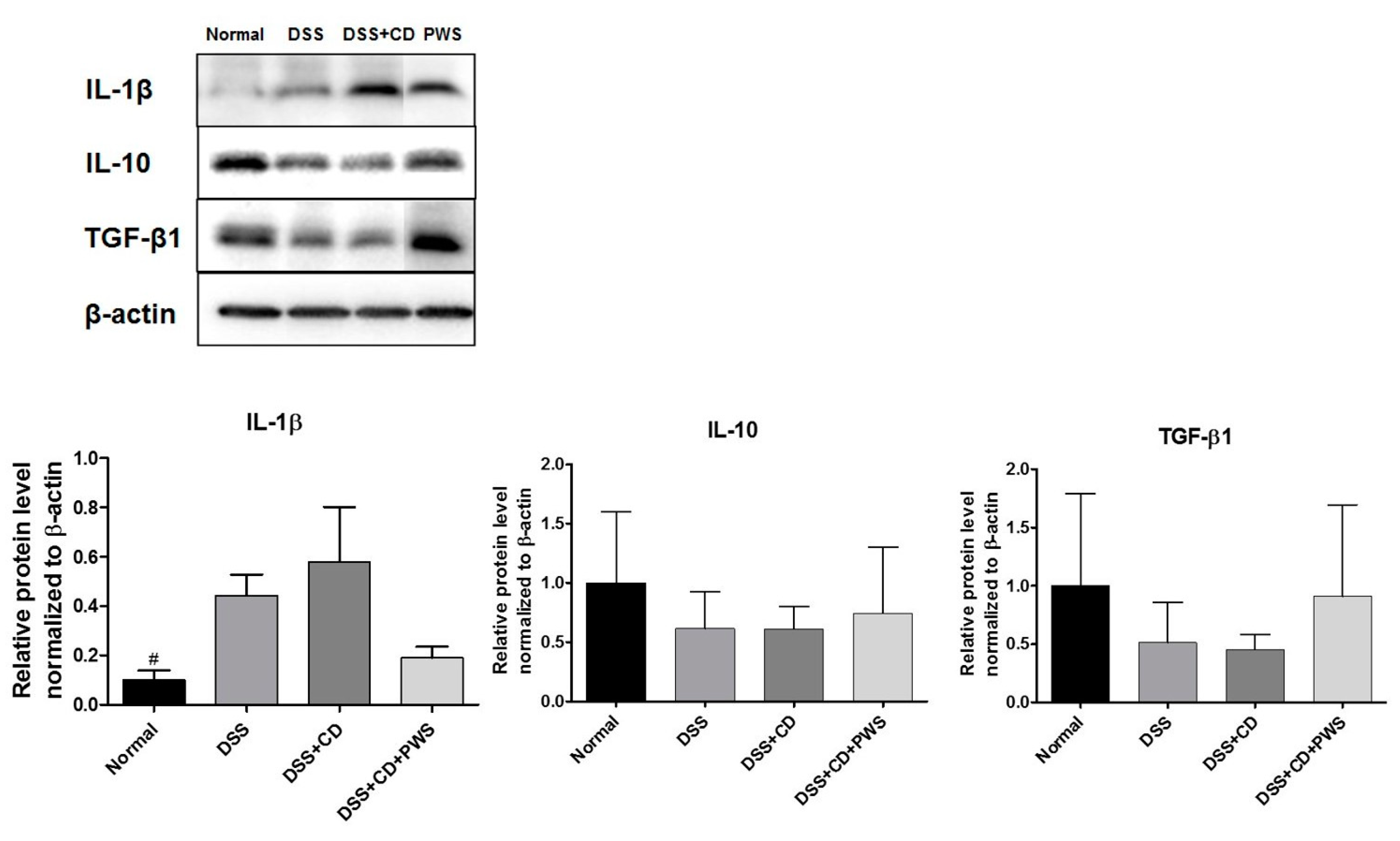
2.2. Effects of PWS on Colitis-Induced Inflammatory Insult
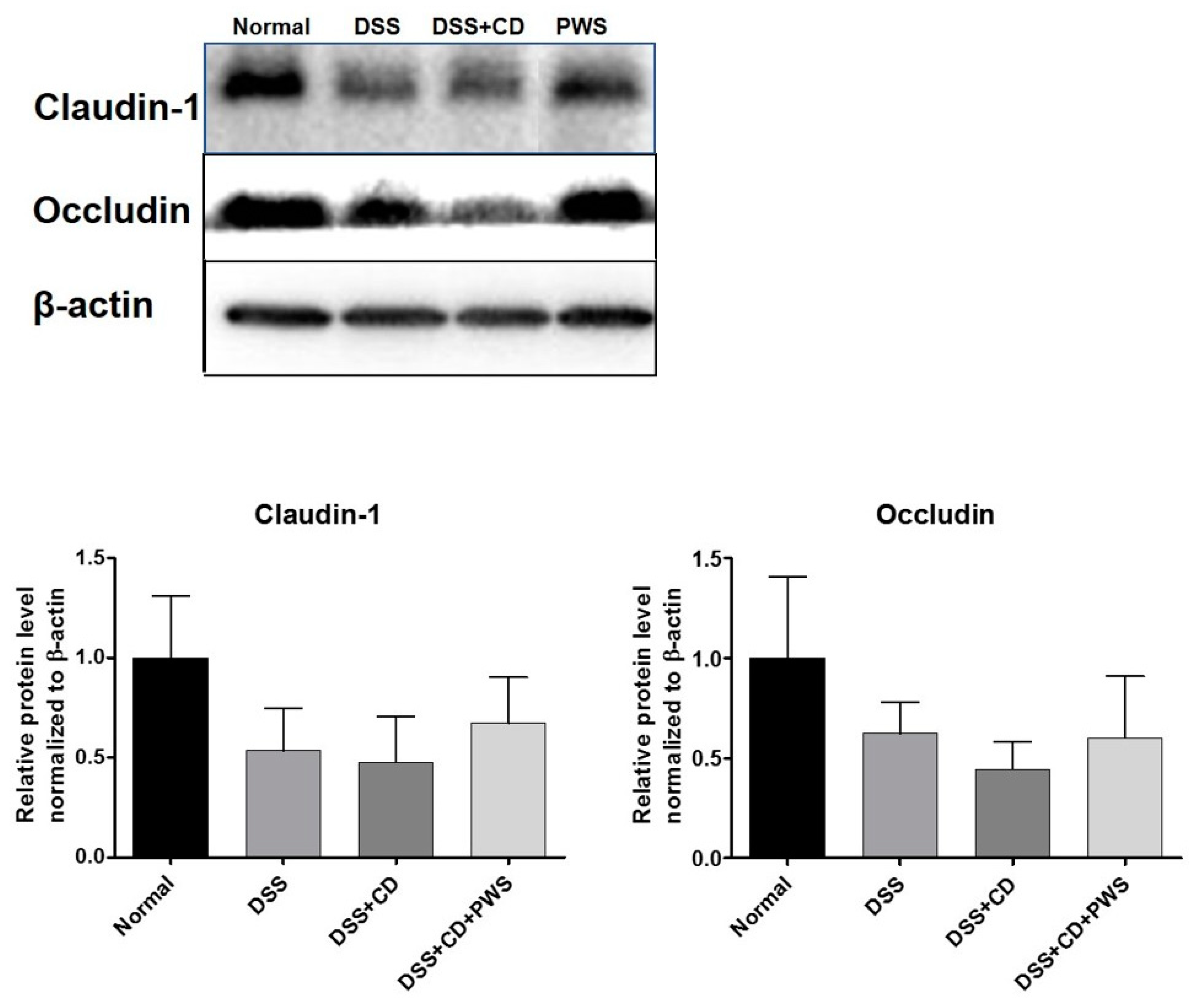
2.3. PWS Protects Intestinal Tight Junction Proteins from Colitis-Induced Damage
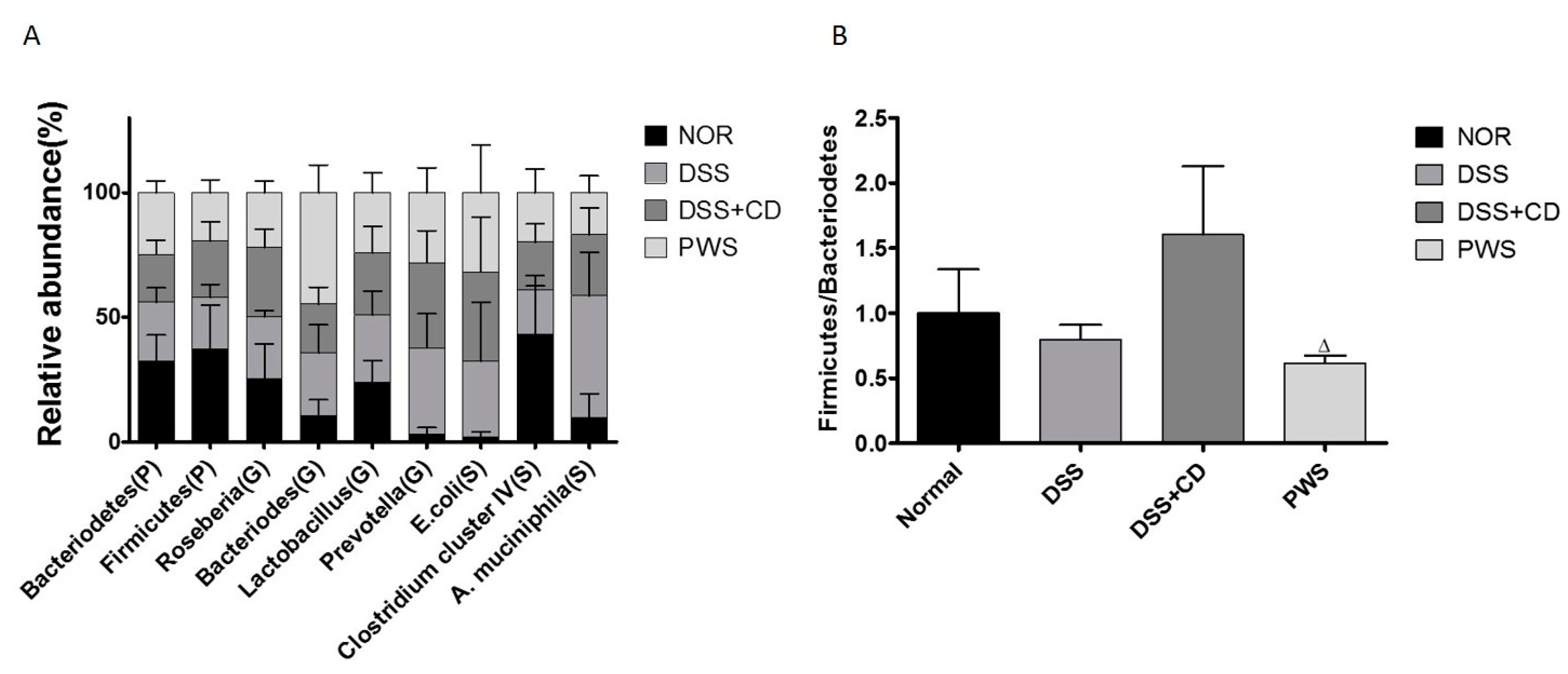
2.4. PWS Treatment Restores the Distribution of Gut Microbial Communities in Colitis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of C. difficile
4.3. Preparation of PWS
4.4. Mice Model of Colitis and Treatment Regimen
4.5. Tissue Sampling, Histology, and Measurement of Disease Activity
4.6. Extraction of Bacterial DNA from Feces
4.7. Purification of DNA by Lithium Chloride
4.8. 16S rRNA-Based Terminal Fragment Length Polymorphism (T-RFLP) Analysis of Gut Microbial Community
4.9. Analysis of Inflammatory and Anti-Inflammatory Markers and Tight Junction Proteins by Western Blotting
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Podolsky, D.K. Inflammatory Bowel Disease. N. Engl. J. Med. 1991, 325, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, C.G.; Loftus, E.V., Jr.; Sandborn, W.J.; Tremaine, W.J.; Harmsen, S.W.; Zinsmeister, A.R.; Melton, J.L., III. Update on the incidence and prevalence of Crohn’s disease and ulcerative colitis in Olmsted County, Minnesota, 1940–2000. Inflamm. Bowel Dis. 2006, 13, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Gassler, N.; Rohr, C.; Schneider, A.; Kartenbeck, J.; Bach, A.; Obermüller, N.; Otto, H.F.; Autschbach, F. Inflammatory bowel disease is associated with changes of enterocytic junctions. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G216–G228. [Google Scholar] [CrossRef]
- Poritz, L.S.; Garver, K.I.; Tilberg, A.F.; Koltun, W.A. Tumor necrosis factor alpha disrupts tight junction assembly. J. Surg. Res. 2004, 116, 14–18. [Google Scholar] [CrossRef]
- McGuckin, M.A.; Eri, R.; Simms, L.A.; Florin, T.H.; Radford-Smith, G. Intestinal barrier dysfunction in inflammatory bowel diseases. Inflamm. Bowel Dis. 2009, 15, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Mielke, L.; Preaudet, A.; Belz, G.; Putoczki, T. Confocal laser endomicroscopy to monitor the colonic mucosa of mice. J. Immunol. Methods 2015, 421, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.; Wang, A.; Shang, H.; Chen, L.; Bao, C.; Wu, L.; Wu, H.; Shi, Y. Herb-partitioned moxibustion upregulated the expression of colonic epithelial tight junction-related proteins in Crohn’s disease model rats. Chin. Med. 2016, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Cummins, P.M. Occludin: One protein, many forms. Mol. Cell. Biol. 2012, 32, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Oshitani, N.; Watanabe, K.; Nakamura, S.; Fujiwara, Y.; Higuchi, K.; Arakawa, T. Dislocation of tight junction proteins without F-actin disruption in inactive Crohn’s disease. Int. J. Mol. Med. 2005, 15, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Walsh, S.V.; Chen, J.; Parkos, C.A.; Nusrat, A. Neutrophil Transmigration in Inflammatory Bowel Disease Is Associated with Differential Expression of Epithelial Intercellular Junction Proteins. Am. J. Pathol. 2001, 159, 2001–2009. [Google Scholar] [CrossRef] [Green Version]
- McAlindon, M.; Hawkey, C.; Mahida, Y. Expression of interleukin 1β and interleukin 1β converting enzyme by intestinal macrophages in health and inflammatory bowel disease. Gut 1998, 42, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Coccia, M.; Harrison, O.J.; Schiering, C.; Asquith, M.J.; Becher, B.; Powrie, F.; Maloy, K.J. IL-1β mediates chronic intestinal inflammation by promoting the accumulation of IL-17A secreting innate lymphoid cells and CD4+ Th17 cells. J. Exp. Med. 2012, 209, 1595–1609. [Google Scholar] [CrossRef] [PubMed]
- Di Sabatino, A.; Pickard, K.M.; Rampton, D.; Kruidenier, L.; Rovedatti, L.; Leakey, N.A.; Corazza, G.R.; Monteleone, G.; MacDonald, T.T. Blockade of transforming growth factor β upregulates T-box transcription factor T-bet, and increases T helper cell type 1 cytokine and matrix metalloproteinase-3 production in the human gut mucosa. Gut 2008, 57, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Jarry, A.; Bossard, C.; Bou-Hanna, C.; Masson, D.; Espaze, E.; Denis, M.G.; Laboisse, C.L. Mucosal IL-10 and TGF-β play crucial roles in preventing LPS-driven, IFN-γ–mediated epithelial damage in human colon explants. J. Clin. Investig. 2008, 118, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Ng, S.C. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef]
- Reiff, C.; Kelly, D. Inflammatory bowel disease, gut bacteria and probiotic therapy. Int. J. Med. Microbiol. 2010, 300, 25–33. [Google Scholar] [CrossRef]
- Nitzan, O.; Elias, M.; Chazan, B.; Raz, R.; Saliba, W. Clostridium difficile and inflammatory bowel disease: Role in pathogenesis and implications in treatment. World J. Gastroenterol. 2013, 19, 7577. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, G.; Yang, Q.; Ye, J.; Cai, X.; Tsering, P.; Cheng, X.; Hu, C.; Zhang, S.; Cao, P. Gut microbiota drives the attenuation of dextran sulphate sodium-induced colitis by Huangqin decoction. Oncotarget 2017, 8, 48863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashtoub, S.; Hoang, B.V.; Vu, M.; Lymn, K.A.; Feinle-Bisset, C.; Howarth, G.S. Clinical and structural effects of traditional Chinese medicine and the herbal preparation, Iberogast, in a rat model of ulcerative colitis. J. Evid. Based Complement. Altern. Med. 2014, 19, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.S.; Lee, J.A.; Jung, D.; Lee, H.Y.; Lee, J.K.; Ha, H.; Lee, M.Y.; Shin, H.K. Simultaneous determination of liquiritin, hesperidin, and glycyrrhizin by HPLC-photodiode array detection and the anti-inflammatory effect of Pyungwi-san. Arch. Pharm. Res. 2011, 34, 203–210. [Google Scholar] [CrossRef]
- Xu, J.Q.; Formulas, W.M. Teaching Reference Series for Higher Educational Institutes in Traditional Chinese Medicine; People’s Health Press: Beijing, China, 1995. [Google Scholar]
- Scheid, V.; Bensky, D.; Ellis, A.; Barolet, R. Chinese Herbal Medicine: Formulas & Strategies; Eastland Press: Vista, CA, USA, 2009. [Google Scholar]
- Tierra, M. Planetary Herbology; Lotus Press: Twin Lakes, WI, USA, 2018. [Google Scholar]
- Keys, J.D. Chinese Herbs, Their Botany, Chemistry, and Pharmacodynamics; Tuttle Company, Inc.: Rutland, VT, USA; Tokyo, Japan, 1976. [Google Scholar]
- Fu, W. Traditional Chinese Medicine and Pharmacology; Foreign Languages Press: Beijing, China, 1985. [Google Scholar]
- Wu, Y.; Fischer, W. Practical Therapeutics of Traditional Chinese Medicine; Paradigm Publications: Tokyo, Japan, 1997. [Google Scholar]
- Choi, I.-Y.; Kim, S.-J.; Jeong, H.-J.; Park, S.-H.; Song, Y.-S.; Lee, J.-H.; Kang, T.-H.; Park, J.-H.; Hwang, G.-S.; Lee, E.-J. Hesperidin inhibits expression of hypoxia inducible factor-1 alpha and inflammatory cytokine production from mast cells. Mol. Cell. Biochem. 2007, 305, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Schröfelbauer, B.; Raffetseder, J.; Hauner, M.; Wolkerstorfer, A.; Ernst, W.; Szolar, O.H. Glycyrrhizin, the main active compound in liquorice, attenuates pro-inflammatory responses by interfering with membrane-dependent receptor signalling. Biochem. J. 2009, 421, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-W.; Jin, Y.; Shin, J.-H.; Kim, I.-D.; Lee, H.-K.; Park, S.; Han, P.-L.; Lee, J.-K. Glycyrrhizic acid affords robust neuroprotection in the postischemic brain via anti-inflammatory effect by inhibiting HMGB1 phosphorylation and secretion. Neurobiol. Dis. 2012, 46, 147–156. [Google Scholar] [CrossRef] [PubMed]
- WANG, J.P.; HO, T.F.; CHANG, L.C.; CHEN, C.C. 857-860Anti-inflammatory Effect of Magnolol, Isolated from Magnolia officinalis, on A23187-induced Pleurisy in Mice. J. Pharm. Pharmacol. 1995, 47, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-P.; Hsu, M.-F.; Raung, S.-Z.; Chen, C.-C.; Kuo, J.-S.; Teng, C.-M. Anti-inflammatory and analgesic effects of magnolol. Naunyn Schmiedeberg Arch. Pharmacol. 1992, 346, 707–712. [Google Scholar] [CrossRef]
- Tsuneki, H.; Ma, E.-L.; Kobayashi, S.; Sekizaki, N.; Maekawa, K.; Sasaoka, T.; Wang, M.-W.; Kimura, I. Antiangiogenic activity of β-eudesmol in vitro and in vivo. Eur. J. Pharmacol. 2005, 512, 105–115. [Google Scholar] [CrossRef]
- Kiso, Y.; Tohkin, M.; Hikino, H. Mechanism of Antihepatotoxic Activity of Atractylon, I: Effect on Free Radical Generation and Lipid Peroxidation1. Planta Med. 1985, 51, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Obukhova, A. Some pharmacological properties of the plant Atracylodes ovata Thunb. Farmakol. Toksikol. 1961, 24, 565–568. [Google Scholar] [PubMed]
- Matsuda, H.; Li, Y.; Taniguchi, K.; Yamahara, J.; Tamai, Y. Imaging analysis of antiulcer action and the active constituent of Atractylodis rhizoma. Yakugaku Zasshi J. Pharm. Soc. Jpn. 1991, 111, 36–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, S.; Kim, H. Evaluation of in vitro anti-inflammatory activities and protective effect of fermented preparations of Rhizoma Atractylodis Macrocephalae on intestinal barrier function against lipopolysaccharide insult. Evid. Based Complement. Altern. Med. 2013, 2013, 363076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-H.; Bose, S.; Kim, H.-G.; Han, K.-S.; Kim, H. Fermented RhizomaAtractylodis Macrocephalae alleviates high fat diet-induced obesity in association with regulation of intestinal permeability and microbiota in rats. Sci. Rep. 2015, 5, 8391. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.K.; Kim, M.; Oh, S.D.; Lee, S.-M.; Sun, B.; Choi, G.S.; Kim, S.-K.; Bae, H.; Kang, C.; Min, B.-I. Effects of Atractylodes macrocephala Koidzumi rhizome on 3T3-L1 adipogenesis and an animal model of obesity. J. Ethnopharmacol. 2011, 137, 396–402. [Google Scholar] [CrossRef]
- Riedlinger, J.E.; Tan, P.W.; Lu, W. Ping Wei San, a Chinese Medicine for Gastrointestinal Disorders. Ann. Pharmacother. 2001, 35, 228–235. [Google Scholar] [CrossRef]
- He, S.-T.; He, F.-Z.; Wu, C.-R.; Li, S.-X.; Liu, W.-X.; Yang, Y.-F.; Jiang, S.-S.; He, G. Treatment of rotaviral gastroenteritis with Qiwei Baizhu powder. World J. Gastroenterol. 2001, 7, 735. [Google Scholar] [CrossRef]
- Kaistha, A.; Levine, J. Inflammatory bowel disease: The classic gastrointestinal autoimmune disease. Curr. Probl. Pediatr. Adolesc. Health Care 2014, 44, 328–334. [Google Scholar] [CrossRef]
- Mylonaki, M.; Langmead, L.; Pantes, A.; Johnson, F.; Rampton, D.S. Enteric infection in relapse of inflammatory bowel disease. Eur. J. Gastroenterol. Hepatol. 2004, 16, 775–778. [Google Scholar] [CrossRef]
- Rodemann, J.F.; Dubberke, E.R.; Reske, K.A.; Seo, D.H.; Stone, C.D. Incidence of Clostridium difficile infection in inflammatory bowel disease. Clin. Gastroenterol. Hepatol. 2007, 5, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Viennois, E.; Chen, F.; Laroui, H.; Baker, M.T.; Merlin, D. Dextran sodium sulfate inhibits the activities of both polymerase and reverse transcriptase: Lithium chloride purification, a rapid and efficient technique to purify RNA. BMC Res. Notes 2013, 6, 360. [Google Scholar] [CrossRef] [PubMed]
- Salaga, M.; Zatorski, H.; Sobczak, M.; Chen, C.; Fichna, J. Chinese herbal medicines in the treatment of IBD and colorectal cancer: A review. Curr. Treat. Options Oncol. 2014, 15, 405–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, I.S.; Seo, C.S.; Ha, H.K.; Lee, M.Y.; Huang, D.S.; Huh, J.I.; Shin, H.K. Genotoxicity assessment of Pyungwi-san (PWS), a traditional herbal prescription. J. Ethnopharmacol. 2011, 133, 696–703. [Google Scholar] [CrossRef]
- Yan, Y. Clinical Observation on 43 Cases of Ulcerative Colitis Treated by Jiawei Pingwei Powder. Guid. J. Tradit. Chin. Med. Pharm. 2009. [Google Scholar] [CrossRef]
- Sasaki, S.; Ishida, Y.; Nishio, N.; Ito, S.; Isobe, K. Thymic involution correlates with severe ulcerative colitis induced by oral administration of dextran sulphate sodium in C57BL/6 mice but not in BALB/c mice. Inflammation 2008, 31, 319–328. [Google Scholar] [CrossRef]
- Morteau, O.; Morham, S.G.; Sellon, R.; Dieleman, L.A.; Langenbach, R.; Smithies, O.; Sartor, R.B. Impaired mucosal defense to acute colonic injury in mice lacking cyclooxygenase-1 or cyclooxygenase-2. J. Clin. Investig. 2000, 105, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Peyrin-Biroulet, L.; Eisenhut, M.; Shin, J.I. IBD immunopathogenesis: A comprehensive review of inflammatory molecules. Autoimmun. Rev. 2017, 16, 416–426. [Google Scholar] [CrossRef]
- Zhao, H.W.; Yue, Y.H.; Han, H.; Chen, X.L.; Lu, Y.G.; Zheng, J.M.; Hou, H.T.; Lang, X.M.; He, L.L.; Hu, Q.L.; et al. Effect of toll-like receptor 3 agonist poly I:C on intestinal mucosa and epithelial barrier function in mouse models of acute colitis. World J. Gastroenterol. 2017, 23, 999–1009. [Google Scholar] [CrossRef]
- Hahm, K.; Im, Y.; Parks, T.; Park, S.; Markowitz, S.; Jung, H.; Green, J.; Kim, S. Loss of transforming growth factor β signalling in the intestine contributes to tissue injury in inflammatory bowel disease. Gut 2001, 49, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.C.; Benjamin, J.L.; McCarthy, N.E.; Hedin, C.R.; Koutsoumpas, A.; Plamondon, S.; Price, C.L.; Hart, A.L.; Kamm, M.A.; Forbes, A.; et al. Relationship between human intestinal dendritic cells, gut microbiota, and disease activity in Crohn’s disease. Inflamm. Bowel Dis. 2011, 17, 2027–2037. [Google Scholar] [CrossRef] [PubMed]
- Casini-Raggi, V.; Kam, L.; Chong, Y.; Fiocchi, C.; Pizarro, T.T.; Cominelli, F. Mucosal imbalance of IL-1 and IL-1 receptor antagonist in inflammatory bowel disease. A novel mechanism of chronic intestinal inflammation. J. Immunol. 1995, 154, 2434–2440. [Google Scholar] [PubMed]
- Zhou, L.; Liu, D.; Xie, Y.; Yao, X.; Li, Y. Bifidobacterium infantis Induces Protective Colonic PD-L1 and Foxp3 Regulatory T Cells in an Acute Murine Experimental Model of Inflammatory Bowel Disease. Gut Liver 2019, 13, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Feng, B.-S.; Yang, S.-B.; Chen, X.; Su, J.; Yang, P.-C. Interleukin (IL)-23 suppresses IL-10 in inflammatory bowel disease. J. Biol. Chem. 2012, 287, 3591–3597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, J.Y.; Jung, J.Y.; Jung, J.Y.; Lee, J.R.; Cho, I.J.; Ku, S.K.; Byun, S.H.; Ahn, Y.-T.; Lee, C.W.; Kim, S.C. Inhibitory effects of traditional herbal formula pyungwi-san on inflammatory response in vitro and in vivo. Evid. Based Complement. Altern. Med. 2013, 2013, 630198. [Google Scholar] [CrossRef] [PubMed]
- Ham, S.-H.; Jeong, S.-E.; Kim, J.-H.; Choi, I.-Y.; Jeong, D.-E.; Lee, G.-Y.; Cho, J.-H. Effects of Pyungwi-san on the production of inflammatory cytokines in LPS-treated rats. Nat. Altern. Med. 2016, 5, 64. [Google Scholar] [CrossRef]
- Schulzke, J.D.; Bojarski, C.; Zeissig, S.; Heller, F.; Gitter, A.H.; Fromm, M. Disrupted barrier function through epithelial cell apoptosis. Ann. N. Y. Acad. Sci. 2006, 1072, 288–299. [Google Scholar] [CrossRef]
- Cammarota, G.; Ianiro, G.; Cianci, R.; Bibbo, S.; Gasbarrini, A.; Curro, D. The involvement of gut microbiota in inflammatory bowel disease pathogenesis: Potential for therapy. Pharmacol. Ther. 2015, 149, 191–212. [Google Scholar] [CrossRef]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef]
- Nell, S.; Suerbaum, S.; Josenhans, C. The impact of the microbiota on the pathogenesis of IBD: Lessons from mouse infection models. Nat. Rev. Microbiol. 2010, 8, 564–577. [Google Scholar] [CrossRef]
- Berry, D.; Schwab, C.; Milinovich, G.; Reichert, J.; Ben Mahfoudh, K.; Decker, T.; Engel, M.; Hai, B.; Hainzl, E.; Heider, S.; et al. Phylotype-level 16S rRNA analysis reveals new bacterial indicators of health state in acute murine colitis. ISME J. 2012, 6, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.; Berry, D.; Rauch, I.; Rennisch, I.; Ramesmayer, J.; Hainzl, E.; Heider, S.; Decker, T.; Kenner, L.; Muller, M.; et al. Longitudinal study of murine microbiota activity and interactions with the host during acute inflammation and recovery. ISME J. 2014, 8, 1101–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musa, S.; Thomson, S.; Cowan, M.; Rahman, T. Clostridium difficile infection and inflammatory bowel disease. Scand. J. Gastroenterol. 2010, 45, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, A.; Zorzi, F.; Del Chierico, F.; Altomare, A.; Cocca, S.; Avola, A.; De Biasio, F.; Russo, A.; Cella, E.; Reddel, S. Fecal and Mucosal microbiota profiling in Irritable bowel syndrome and Inflammatory Bowel Disease. Front. Microbiol. 2019, 10, 1655. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, E.; Amir, I.; Zafran, M.; Gophna, U.; Samra, Z.; Pitlik, S.; Bishara, J. The correlation between Clostridium-difficile infection and human gut concentrations of Bacteroidetes phylum and clostridial species. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 377–383. [Google Scholar] [CrossRef]
- Van den Bossche, L.; Hindryckx, P.; Devisscher, L.; Devriese, S.; Van Welden, S.; Holvoet, T.; Vilchez-Vargas, R.; Vital, M.; Pieper, D.H.; Vanden Bussche, J.; et al. Ursodeoxycholic Acid and Its Taurine- or Glycine-Conjugated Species Reduce Colitogenic Dysbiosis and Equally Suppress Experimental Colitis in Mice. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Dicksved, J.; Schreiber, O.; Willing, B.; Petersson, J.; Rang, S.; Phillipson, M.; Holm, L.; Roos, S. Lactobacillus reuteri maintains a functional mucosal barrier during DSS treatment despite mucus layer dysfunction. PLoS ONE 2012, 7, e46399. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, M.; Dogan, B.; Rishniw, M.; Weitzman, G.; Bosworth, B.; Yantiss, R.; Orsi, R.H.; Wiedmann, M.; McDonough, P.; Kim, S.G. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Järnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology 2010, 139, 1844–1854. [Google Scholar] [CrossRef]
- Willing, B.; Halfvarson, J.; Dicksved, J.; Rosenquist, M.; Järnerot, G.; Engstrand, L.; Tysk, C.; Jansson, J.K. Twin studies reveal specific imbalances in the mucosaassociated microbiota of patients with ileal Crohn’s disease. Inflamm. Bowel Dis. 2008, 15, 653–660. [Google Scholar] [CrossRef]
- Hall, G.; Kurosawa, S.; Stearns-Kurosawa, D. Dextran sulfate sodium colitis facilitates colonization with Shiga toxin-producing Escherichia coli: A novel murine model for the study of Shiga toxicosis. Infect. Immun. 2018, 86, e00530-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakansson, A.; Tormo-Badia, N.; Baridi, A.; Xu, J.; Molin, G.; Hagslatt, M.L.; Karlsson, C.; Jeppsson, B.; Cilio, C.M.; Ahrne, S. Immunological alteration and changes of gut microbiota after dextran sulfate sodium (DSS) administration in mice. Clin. Exp. Med. 2015, 15, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Zhou, G.; Wang, M.; Peng, Y.; Li, X. The metabolism of polysaccharide from Atractylodes macrocephala Koidz and its effect on intestinal microflora. Evid. Based Complement. Altern. Med. 2014, 2014, 926381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, P.; Li, D.; Hu, X.; Chen, F. Beneficial effects of ginger on prevention of obesity through modulation of gut microbiota in mice. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed]
- Fimlaid, K.A.; Jensen, O.; Donnelly, M.L.; Siegrist, M.S.; Shen, A. Regulation of Clostridium difficile spore formation by the SpoIIQ and SpoIIIA proteins. PLoS Genet. 2015, 11, e1005562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theriot, C.M.; Koenigsknecht, M.J.; Carlson, P.E., Jr.; Hatton, G.E.; Nelson, A.M.; Li, B.; Huffnagle, G.B.; Li, J.Z.; Young, V.B. Antibiotic-induced shifts in the mouse gut microbiome and metabolome increase susceptibility to Clostridium difficile infection. Nat. Commun. 2014, 5, 3114. [Google Scholar] [CrossRef]
- Yamabayashi, S. Periodic acid—Schiff—Alcian Blue: A method for the differential staining of glycoproteins. Histochem. J. 1987, 19, 565–571. [Google Scholar] [CrossRef]
- Kerr, T.; Ciorba, M.; Matsumoto, H.; Davis, V.; Luo, J.; Kennedy, S.; Xie, Y.; Shaker, A.; Dieckgraefe, B.; Davidson, N. Dextran sodium sulfate inhibition of real-time polymerase chain reaction amplification: A poly-A purification solution. Inflamm. Bowel Dis. 2011, 18, 344–348. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Botanical Name | Pharmaceutical Name | Approximate Amount (%) |
|---|---|---|
| Atyractylodes lancea | rhizoma atractylodis | 29 |
| Magnolia officinalis | cortex magnoliae officinalis | 14 |
| Citrus reticulata Blanco | Pericarpium citri reticulata | 20 |
| Glycyrrhiza uralensis | radix glycyrrhizae uralensis | 9 |
| Zingiber officinalis | rhizoma zingiberis officinalis recens | 14 |
| Ziziphus jujuba mill | fructus zizyphi jujubae | 14 |
| Score | Diarrheal Stool Score | Bloody Stool Score |
|---|---|---|
| 0 | Normal stool | Normal color |
| 1 | Mildly soft stool | Dark brown |
| 2 | very soft stool | Reddish |
| 3 | watery stool | Bloody |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Bose, S.; Lim, S.-K.; Kim, H. Preventive Effects of Pyungwi-san against Dextran Sulfate Sodium- and Clostridium difficile-Induced Inflammatory Bowel Disease in Mice. Int. J. Mol. Sci. 2019, 20, 6346. https://doi.org/10.3390/ijms20246346
Yang M, Bose S, Lim S-K, Kim H. Preventive Effects of Pyungwi-san against Dextran Sulfate Sodium- and Clostridium difficile-Induced Inflammatory Bowel Disease in Mice. International Journal of Molecular Sciences. 2019; 20(24):6346. https://doi.org/10.3390/ijms20246346
Chicago/Turabian StyleYang, Meng, Shambhunath Bose, Soo-Kyoung Lim, and Hojun Kim. 2019. "Preventive Effects of Pyungwi-san against Dextran Sulfate Sodium- and Clostridium difficile-Induced Inflammatory Bowel Disease in Mice" International Journal of Molecular Sciences 20, no. 24: 6346. https://doi.org/10.3390/ijms20246346
APA StyleYang, M., Bose, S., Lim, S. -K., & Kim, H. (2019). Preventive Effects of Pyungwi-san against Dextran Sulfate Sodium- and Clostridium difficile-Induced Inflammatory Bowel Disease in Mice. International Journal of Molecular Sciences, 20(24), 6346. https://doi.org/10.3390/ijms20246346