Ferulic Acid Improves Depressive-Like Behavior in Prenatally-Stressed Offspring Rats via Anti-Inflammatory Activity and HPA Axis

Abstract
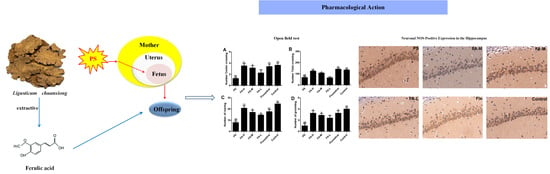
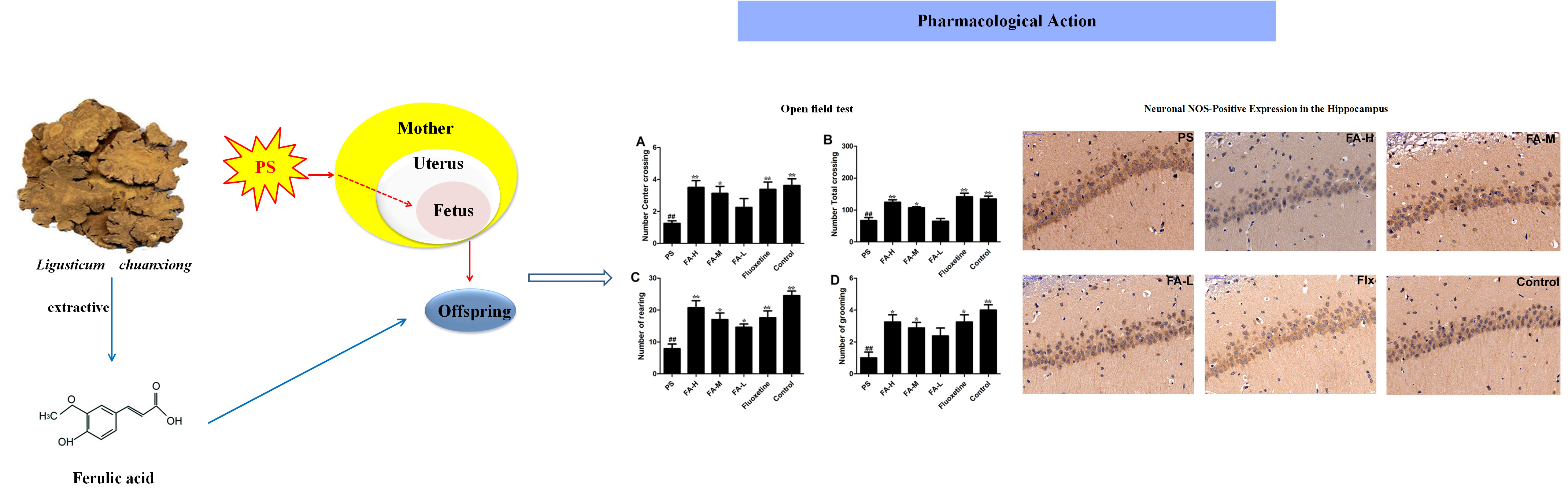
:
1. Introduction
2. Results
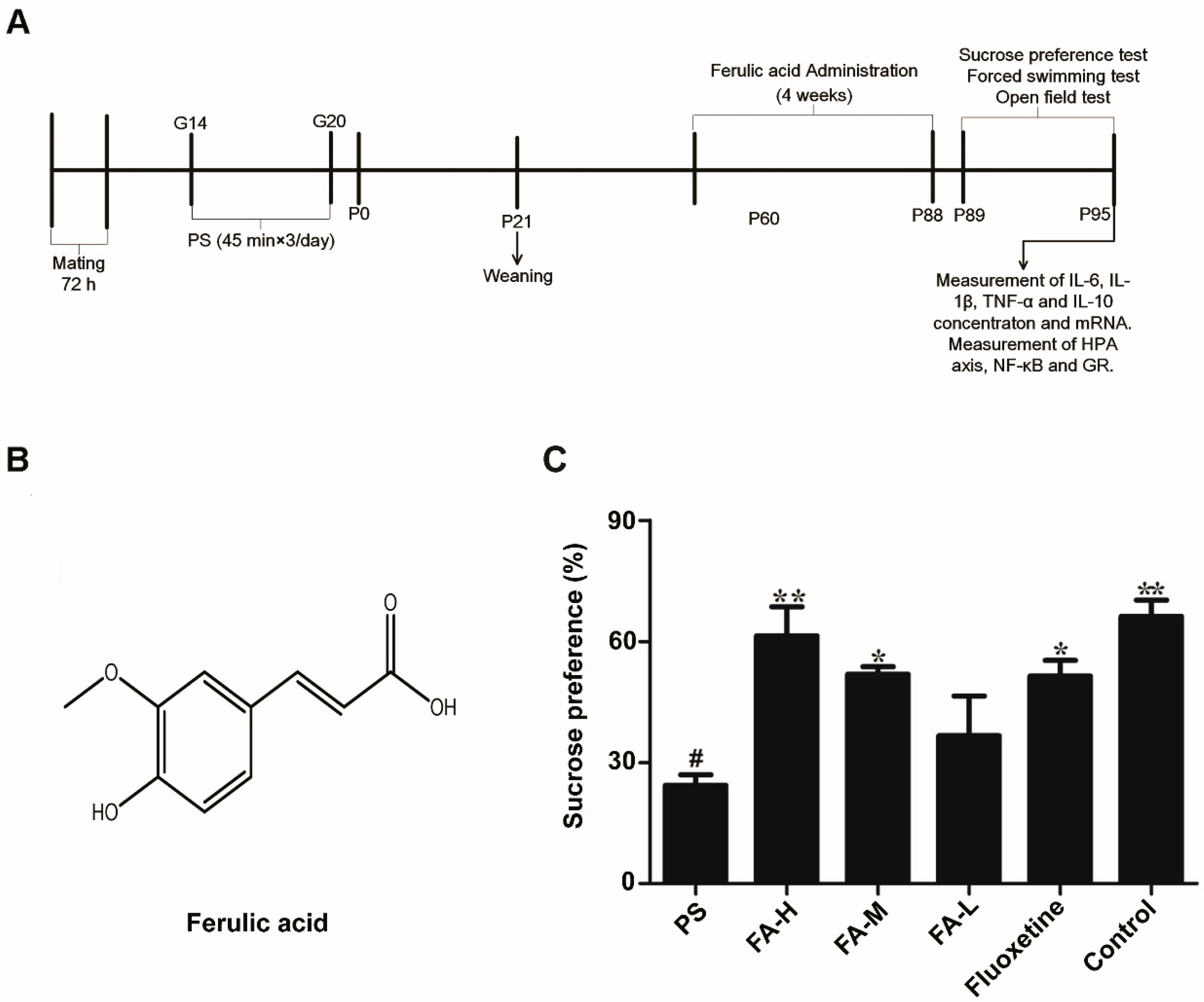
2.1. Effects of FA on Sucrose Preference Test
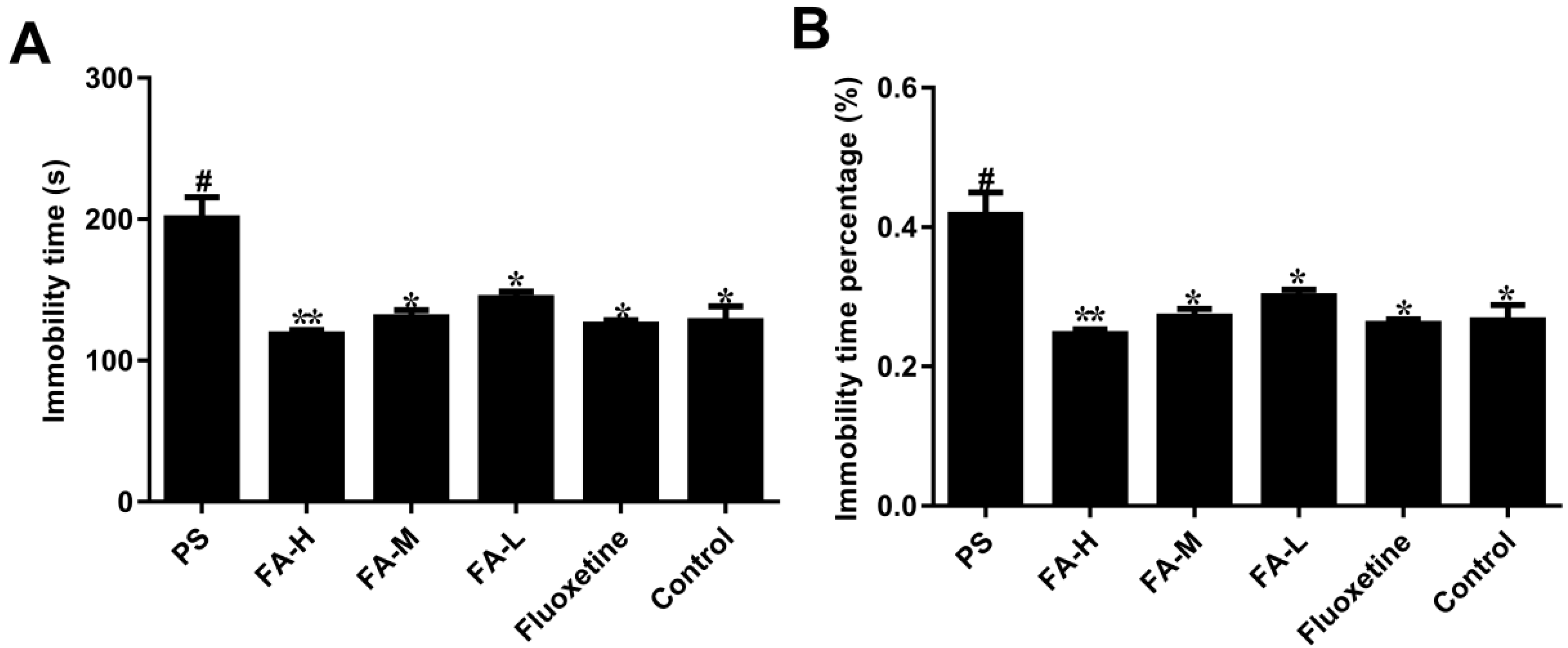
2.2. Effects of FA on Immobility in the Forced Swim Test
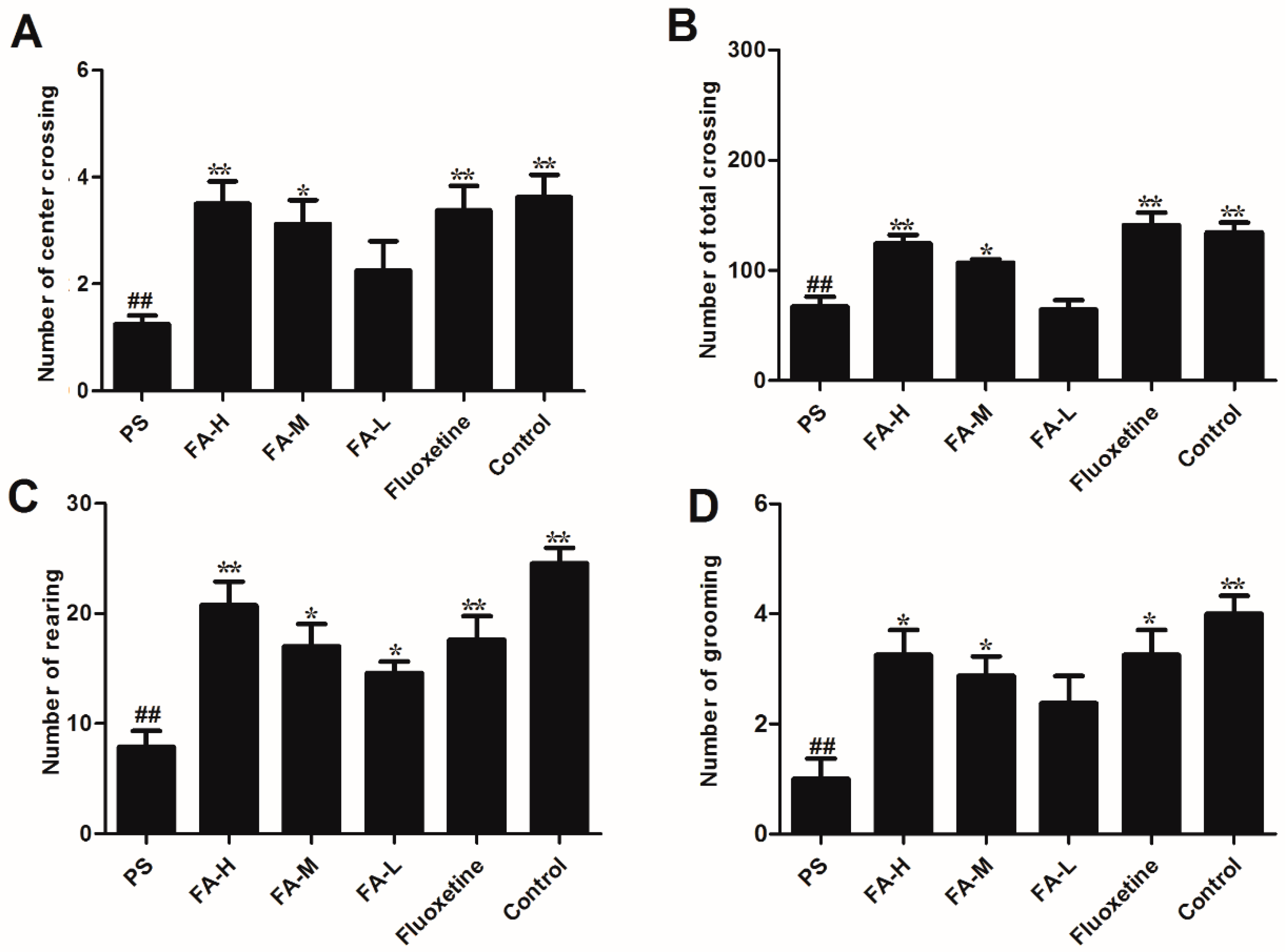
2.3. Effects of FA on Open Field Test
2.4. Effects of FA on ACTH and Corticosterone Levels
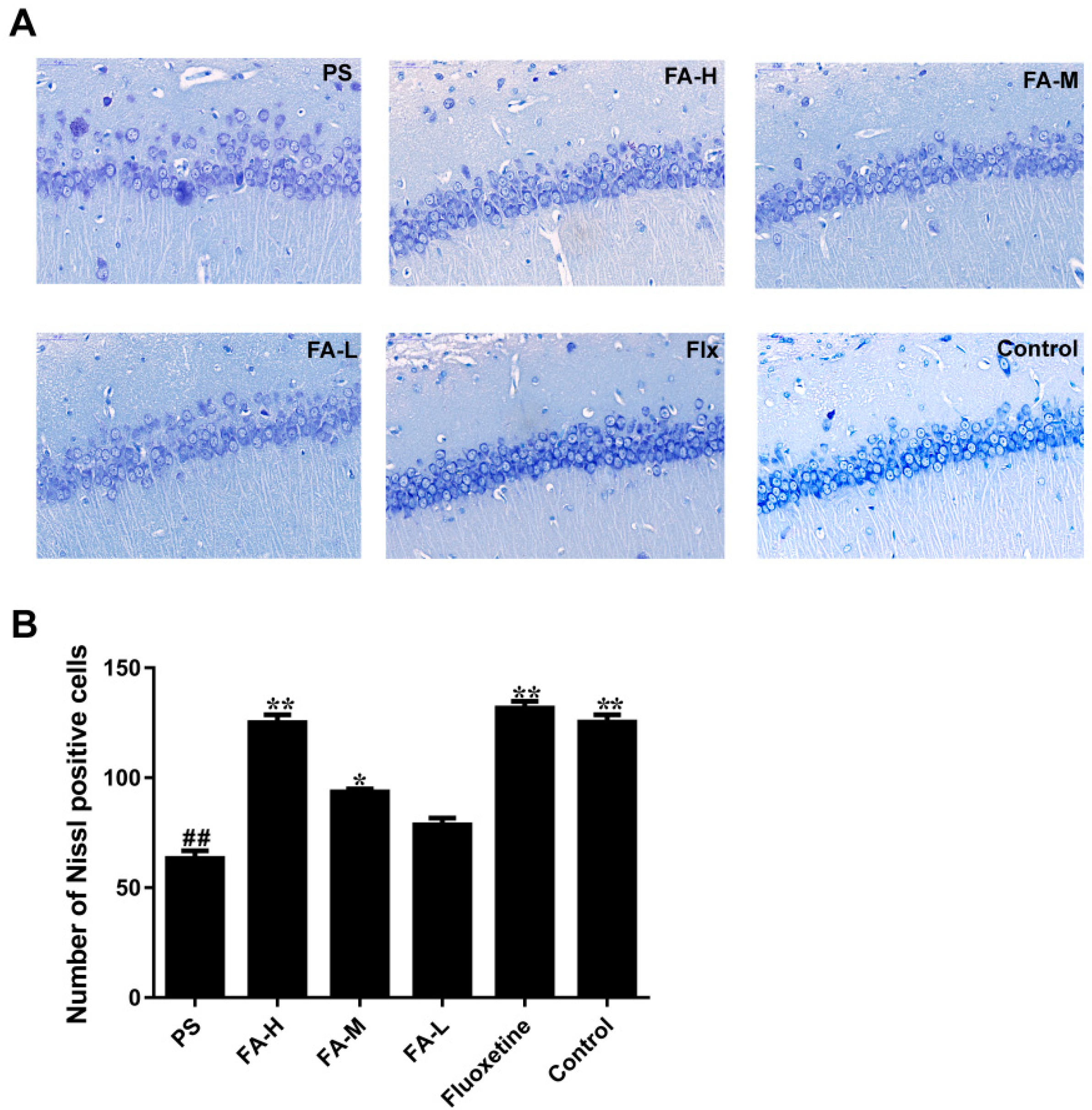
2.5. Effects of FA on Nissl Staining in the Hippocampus of Offspring
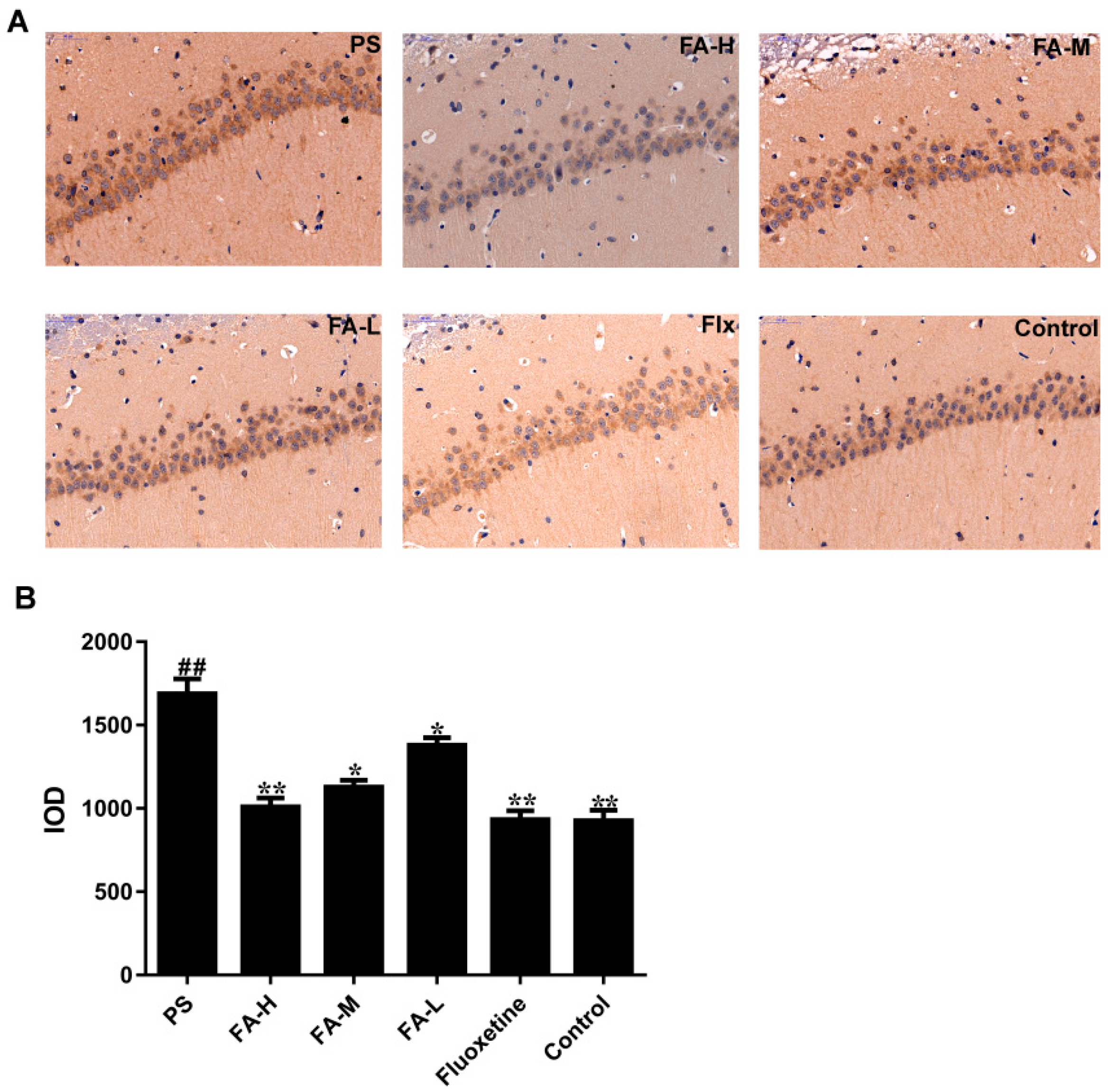
2.6. Effects of FA on Neuronal NOS-Positive Expression in the Hippocampus of Offspring
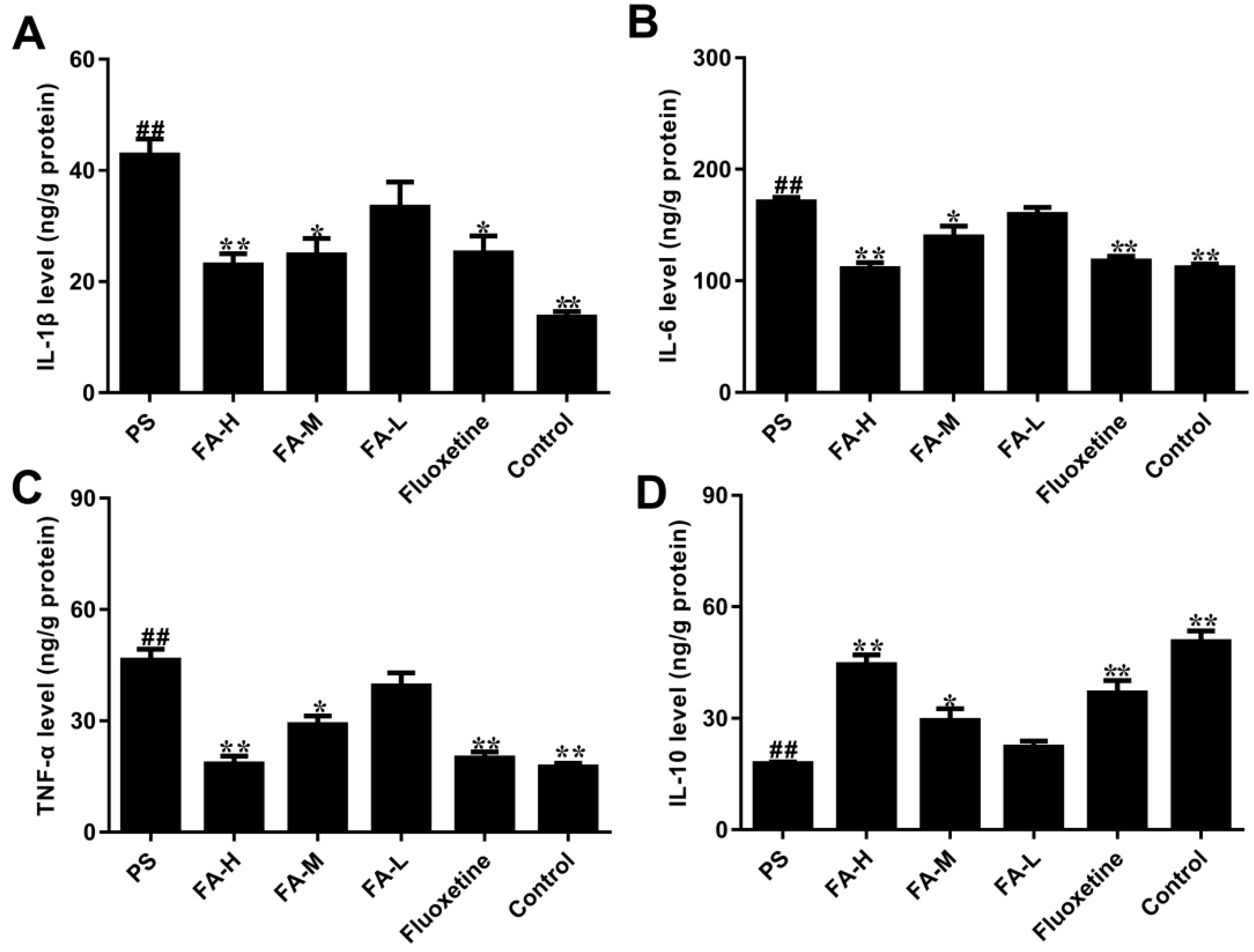
2.7. Effects of FA on Cytokine Production
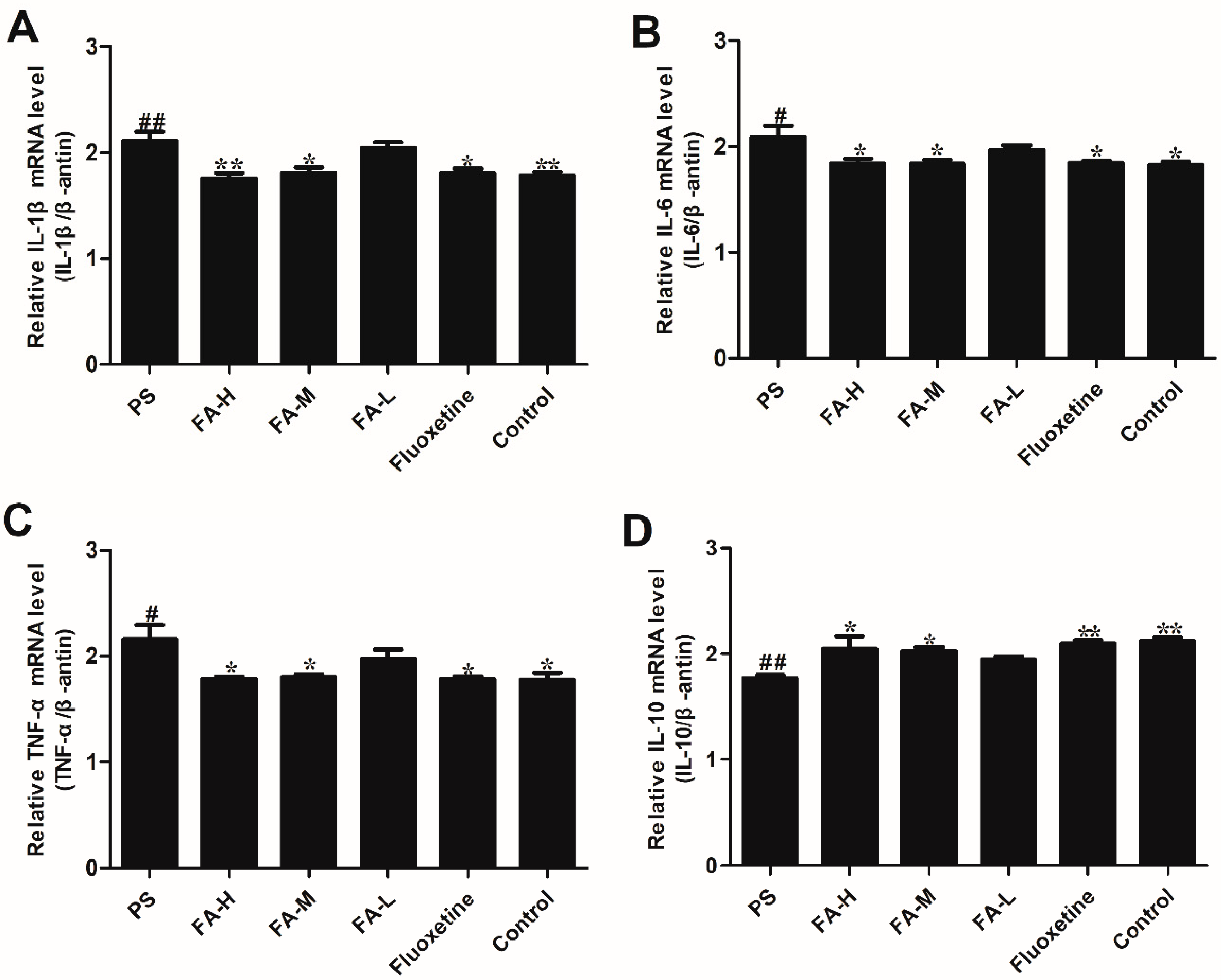
2.8. Effects of FA on Cytokine mRNA Expression
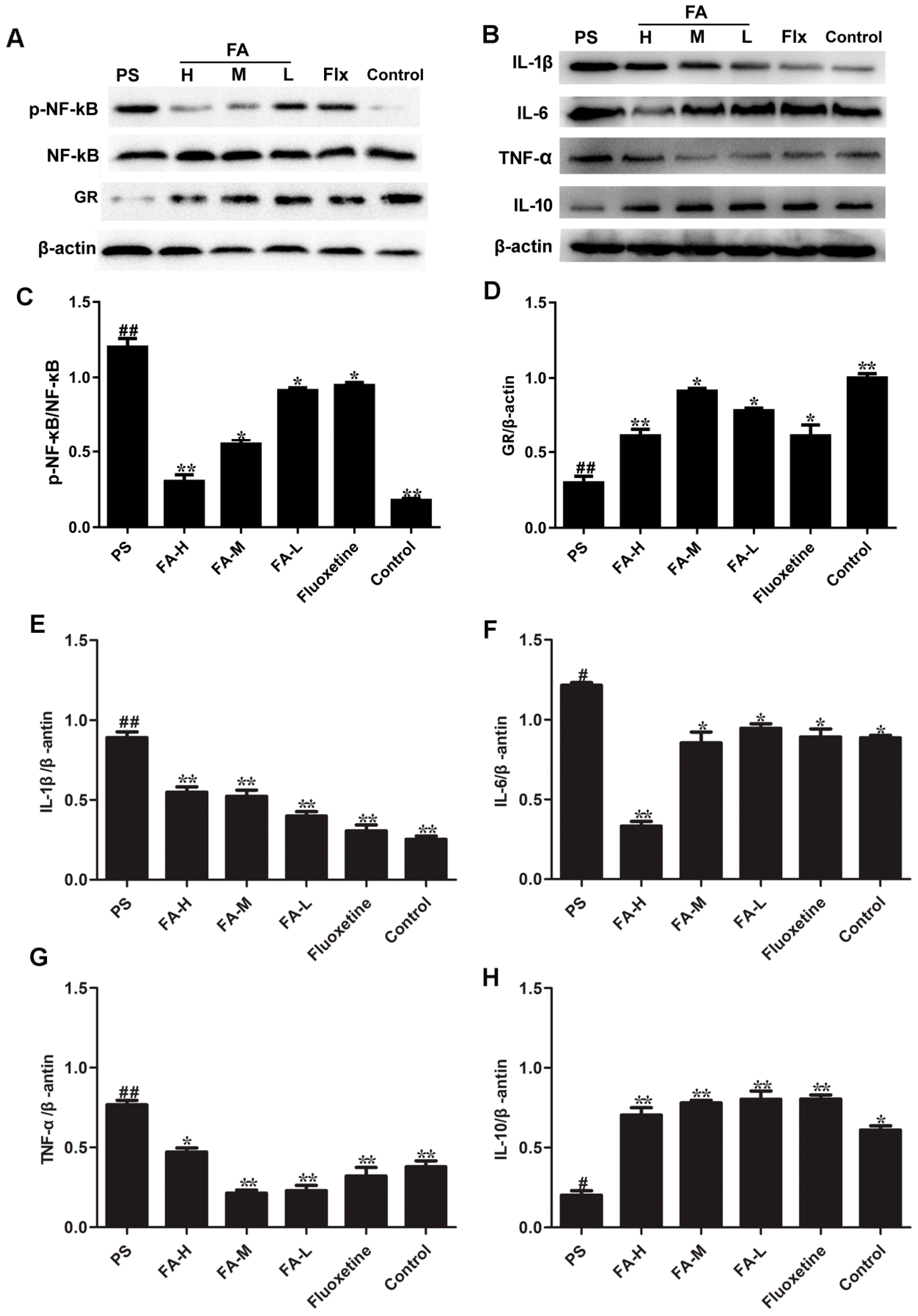
2.9. Effects of FA on NF-κB Activation and GR Expression
3. Discussion
4. Materials and Methods
4.1. Animals and Procedures
4.2. Administration of FA in Rats
4.3. Sucrose Preference Test
4.4. Forced Swimming Test (FST)
4.5. Open Field Test (OFT)
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. RNA Isolation and Real-time PCR
4.8. Nissl Staining
4.9. Immunohistochemical Assay for nNOS
4.10. Western Blot Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Rice, F.I.; Thapar, J.A. The impact of gestational stress and prenatal growth on emotional problems in offspring: A review. Acta Psychiatr. Scand. 2007, 115, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, P.J.; Ferreira, H.H.A.; Antunes, E.; Teixeira, N.A. Chronic mild prenatal stress exacerbates the allergen-induced airway inflammation in rats. Mediat. Inflamm. 1999, 8, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, Y.; Hu, J.; Cheng, W.; Jiang, H.; Zhang, X.; Li, M.; Ren, J.; Li, X. Prenatal chronic mild stress induces depression-like behavior and sex-specific changes in regional glutamate receptor expression patterns in adult rats. Neuroscience 2015, 301, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, M. The long-term behavioural consequences of prenatal stress. Neurosci. Biobehav. Rev. 2008, 32, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Mailliot, V.; Wolowczuk, I.; Mairesse, J.; Viltart, O.; Delacre, M.; Khalife, J.; Chartier Harlin, M.C.; Maccari, S. Prenatal stress has pro-inflammatory consequences on the immune system in adult rats. Psychoneuroendocrino 2007, 32, 114–124. [Google Scholar]
- Wu, Y. Impact of prenatal stress and adulthood stress on immune system a review. Biomed. Res. 2012, 23, 315–320. [Google Scholar]
- Diz-Chaves, Y.; Astiz, M.; Bellini, MJ.; Garcia-Segura, L.M. Prenatal stress increases the expression of proinflammatory cytokines and exacerbates the inflammatory response to LPS in the hippocampal formation of adult male mice. Brain Behav. Immun. 2013, 28, 196–206. [Google Scholar] [PubMed] [Green Version]
- Veru, F.; Laplante, D.P.; Luheshi, G.; King, S. Prenatal maternal stress exposure and immune function in the offspring. Stress 2014, 17, 133–148. [Google Scholar] [CrossRef]
- Hankin, B.L. Adolescent depression: Description, causes, and interventions. Epilepsy Behav. 2006, 8, 102–114. [Google Scholar]
- Zhu, Z.L.; Li, X.; Chen, W.N.; Zhao, Y.; Li, H.; Cai, Q.; Jia, N.; Bai, Z.L.; Liu, J.K. Prenatal stress causes gender-dependent neuronal loss and oxidative in Rat Hippocampus. J. Neurosci. Res. 2004, 78, 837–844. [Google Scholar] [CrossRef]
- Glover, V.; O’Connor, T.G.; O’Donnell, K. Prenatal stress and the programming of the HPA axis. Neurosci. Biobehav. Rev. 2010, 35, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Hodes, G.E.; Kana, V.; Menard, C.; Merad, M.; Russo, S.J. Neuroimmune mechanisms of depression. Nat. Neurosci. 2015, 18, 1386–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Neuroscience 2008, 9, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, F.N.; Costa, A.P.; Ghisleni, G.; Diaz, A.P.; Rodrigues, A.L.S.; Peluffo, H.; Kaster, M.P. NLRP3 inflammasome-driven pathways in depression: Clinical and preclinical findings. Brain Behav. Immun. 2017, 64, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lin, S.; Qin, T.; Li, H.; Ma, Z.; Ma, S. Senegenin exerts anti-depression effect in mice induced by chronic un-predictable mild stress via inhibition of NF-kappaB regulating NLRP3 signal pathway. Int. Immunopharmacol. 2017, 53, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xu, C.; Wu, X.; Liu, F.; Du, Y.; Sun, J.; Tao, J.; Dong, J. Icariin exerts an antidepressant effect in an unpredictable chronic mild stress model of depression in rats and is associated with the regulation of hippocampal neuroinflammation. Neuroscience 2015, 294, 193–205. [Google Scholar] [CrossRef]
- Song, Y.; Sun, R.; Ji, Z.; Li, X.; Fu, Q.; Ma, S. Perilla aldehyde attenuates CUMS-induced depressive-like behaviors via regulating TXNIP/TRX/NLRP3 pathway in rats. Life Sci. 2018, 206, 117–124. [Google Scholar] [CrossRef]
- Cao, Y.J.; Zhang, Y.M.; Qi, J.P.; Liu, R.; Zhang, H.; He, L.C. Ferulic acid inhibits H2O2-induced oxidative stress and inflammation in rat vascular smooth muscle cells via inhibition of the NADPH oxidase and NF-kappaB pathway. Int. Immunopharmacol. 2015, 28, 1018–1025. [Google Scholar] [CrossRef]
- Hassanzadeh, P.; Arbabi, E.; Atyabi, F.; Dinarvand, R. Ferulic acid exhibits antiepileptogenic effect and prevents oxidative stress and cognitive impairment in the kindlingmodel of epilepsy. Life Sci. 2017, 179, 9–14. [Google Scholar] [CrossRef]
- Ogle, W.O.; Speisman, R.B.; Ormerod, B.K. Potential of treating age-related depression and cognitive decline with nutraceutical approaches: A mini-review. Gerontology 2013, 59, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Mamiya, T.; Kise, M.; Morikawa, K. Ferulic acid attenuated cognitive deficits and increase in carbonyl proteins induced by buthionine-sulfoximine in mice. Neurosci. Lett. 2008, 430, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Yabe, T.; Hirahara, H.; Harada, N.; Ito, N.; Ito, N.; Nagai, T.; Sanagi, T.; Yamada, H. Ferulic acid induces neural progenitor cell proliferation in vitro and in vivo. Neuroscience 2010, 165, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Pincus-Knackstedt, M.K.; Joachim, R.A.; Blois, S.M.; Douglas, A.J.; Orsal, A.S.; Klapp, B.F.; Wahn, U.; Hamelmann, E.; Arck, P.C. Prenatal Stress Enhances Susceptibility of Murine Adult Offspring toward Airway Inflammation. J. Immunol. 2006, 177, 8484–8492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plant, D.T.; Pawlby, S.; Sharp, D.; Zunszain, P.A.; Pariante, C.M. Prenatal maternal depression is associated with offspring inflammation at 25 years: A prospective longitudinal cohort study. Transl. Psychiatry 2016, 6, e936. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of Depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Duman, R.S. Stress, depression, and neuroplasticity: A convergence of mechanisms. Neuropsychopharmacol 2008, 33, 88–109. [Google Scholar] [CrossRef]
- Silberman, D.M.; Acosta, G.B.; Zorrilla Zubilete, M.A. Long-term effects of early life stress exposure: Role of epigenetic mechanisms. Pharmacol. Res. 2016, 109, 64–73. [Google Scholar] [CrossRef]
- InbredHarris, A.; Seckl, J. Glucocorticoids, prenatal stress and the programming of disease. Horm. Behav. 2011, 59, 279–289. [Google Scholar]
- Anderson, G. Linking the biological underpinnings of depression: Role of mitochondria interactions with melatonin, inflammation, sirtuins, tryptophan catabolites, DNA repair and oxidative and nitrosative stress, with consequences for classification and cognition. Prog. Neuro-Psychopharmacol. 2017, 8, 255–266. [Google Scholar] [CrossRef]
- Zeni, A.L.B.; Zomkowski, A.D.E.; Maraschin, M.; Rodrigues, A.L.S.; Tasca, C.I. Ferulic acid exerts antidepressant-like effect in the tail suspension test in mice: Evidence for the involvement of the serotonergic system. Eur. J. Pharmacol. 2012, 679, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeni, A.L.B.; Zomkowski, A.D.E.; Maraschin, M.; Rodrigues, A.L.S.; Tasca, C. Involvement of PKA, CaMKII, PKC, MAPK/ERK and PI3K in the acute antidepressant-like effect of ferulic acid in the tail suspension test. Pharmacol. Biochem. Behav. 2012, 103, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenk, G.L.; Gramling, K.M.; Wegrzyniak, B.H.; Ronchetti, D.; Maucci, R.S.; Rosi, L.G.; Ongini, E. Attenuation of chronic neuroinflammation by a nitric oxide-releasing derivative of the antioxidant ferulic acid. J. Neurochem. 2004, 89, 484–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Jia, N.; Zhao, X.Y.; Tang, G.K.; Guan, L.X.; Wang, D.; Sun, H.L.; Li, H.; Zhu, Z.L. Involvement of pGluR1, EAAT2 and EAAT3 in offspring depression induced by prenatal stress. Neuroscience 2013, 250, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.L.; Guan, L.X.; Zhu, Z.L.; Li, H. Reduced levels of NR1 and NR2A with depression-like behavior in different brain regions in prenatally stressed juvenile offspring. PLoS ONE 2013, 8, e81775. [Google Scholar] [CrossRef] [PubMed]
- Shatillo, A.; Salo, R.A.; Giniatullin, R.; Gröhna, O.H. 2015. Involvement of NMDA receptor subtypes in cortical spreading depression in rats assessed by fMRI. Neuropharmacology 2013, 93, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.J.; He, X.; Wang, N.; He, L.C. Effects of imperatorin, the active component from Radix Angelicae (Baizhi), on the blood pressure and oxidative stress in 2K,1C hypertensive rats. Phytomedicine 2013, 20, 1048–1054. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward/ Reverse | Sequence |
|---|---|---|
| IL-1β | Forward | 5′-aatgcctcgtgctgctg-3′ |
| Reverse | 5′-tgtcgttgcttgtctctcc-3′ | |
| IL-6 | Forward | 5′-ccagagtcattcagagcaatac-3′ |
| Reverse | 5′-gatggtcttggtccttagcc-3′ | |
| IL-10 | Forward | 5′-tgccttcagtcaagtgaagact-3′ |
| Reverse | 5′-aaactcattcatgccttgta-3′ | |
| TNF-α | Forward | 5′-ccacgctcttctgtctactg-3′ |
| Reverse | 5′-ctacgggcttgtcactcg-3′ | |
| β-actin | Forward | 5′-tacaaccttcttgcagctcctc-3 |
| Reverse | 5′-gccgtgttcaatggggtact-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Cheng, Y.; Chen, Y.; Yue, Y.; Li, Y.; Xia, S.; Li, Y.; Deng, H.; Zhang, J.; Cao, Y. Ferulic Acid Improves Depressive-Like Behavior in Prenatally-Stressed Offspring Rats via Anti-Inflammatory Activity and HPA Axis. Int. J. Mol. Sci. 2019, 20, 493. https://doi.org/10.3390/ijms20030493
Zheng X, Cheng Y, Chen Y, Yue Y, Li Y, Xia S, Li Y, Deng H, Zhang J, Cao Y. Ferulic Acid Improves Depressive-Like Behavior in Prenatally-Stressed Offspring Rats via Anti-Inflammatory Activity and HPA Axis. International Journal of Molecular Sciences. 2019; 20(3):493. https://doi.org/10.3390/ijms20030493
Chicago/Turabian StyleZheng, Xingxing, Ying Cheng, Yiwei Chen, Yisong Yue, Yingchun Li, Sizhe Xia, Yang Li, Huanhuan Deng, Junli Zhang, and Yanjun Cao. 2019. "Ferulic Acid Improves Depressive-Like Behavior in Prenatally-Stressed Offspring Rats via Anti-Inflammatory Activity and HPA Axis" International Journal of Molecular Sciences 20, no. 3: 493. https://doi.org/10.3390/ijms20030493
APA StyleZheng, X., Cheng, Y., Chen, Y., Yue, Y., Li, Y., Xia, S., Li, Y., Deng, H., Zhang, J., & Cao, Y. (2019). Ferulic Acid Improves Depressive-Like Behavior in Prenatally-Stressed Offspring Rats via Anti-Inflammatory Activity and HPA Axis. International Journal of Molecular Sciences, 20(3), 493. https://doi.org/10.3390/ijms20030493