Translational Control of Canonical and Non-Canonical Translation Initiation Factors at the Sea Urchin Egg to Embryo Transition



Abstract
:1. Introduction
2. Results
2.1. mRNA Coding Translation Initiation Factors Involved in mRNA Recognition Are Present as Maternal mRNAs in Sea Urchin Eggs
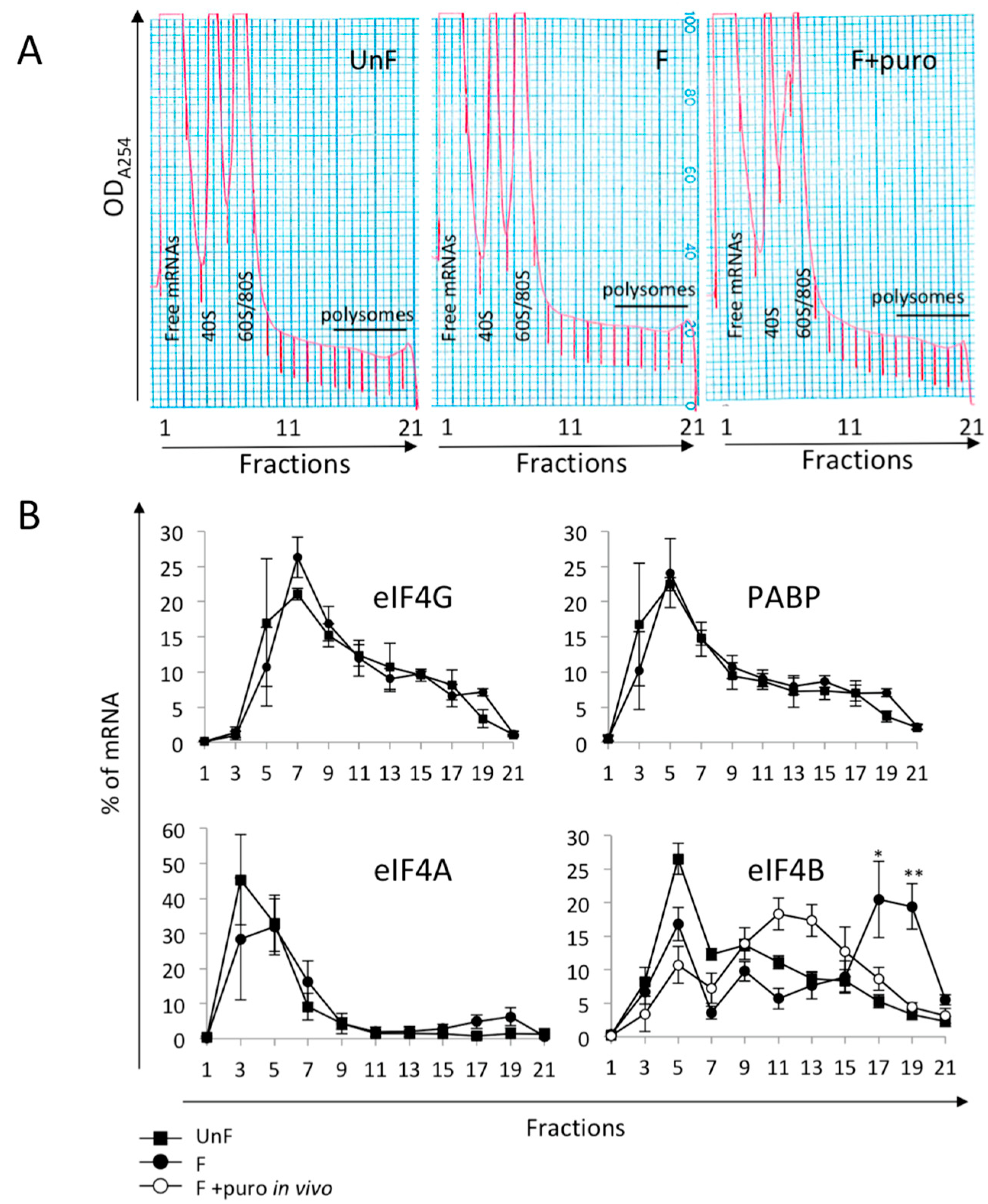
2.2. Initiation Factors eIF4A, eIF4G, or PABP Are not Recruited into Polysomes after Fertilization
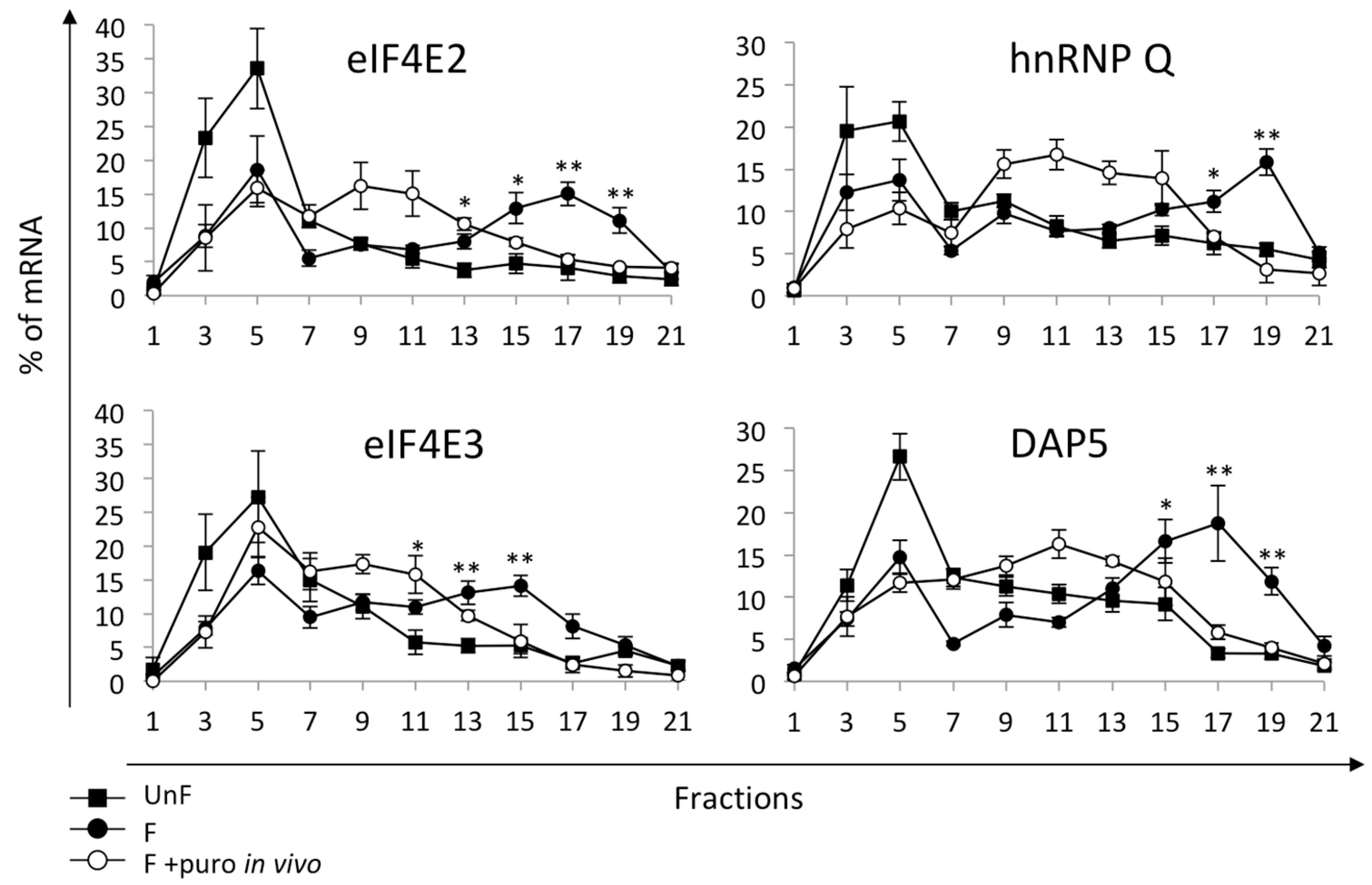
2.3. Non-Canonical Initiation Complex mRNAs Are Present in Unfertilized Eggs and Translated at Fertilization
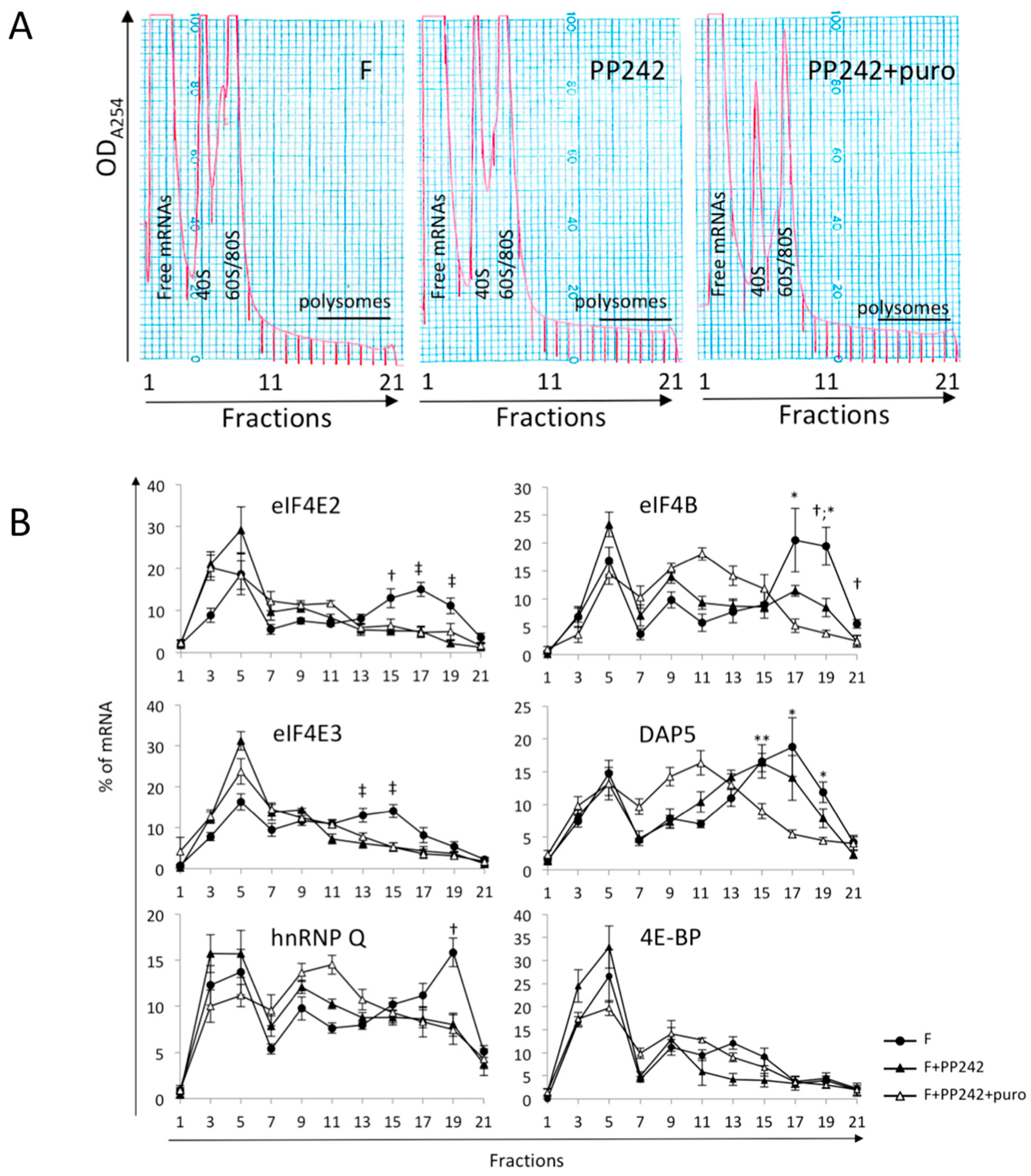
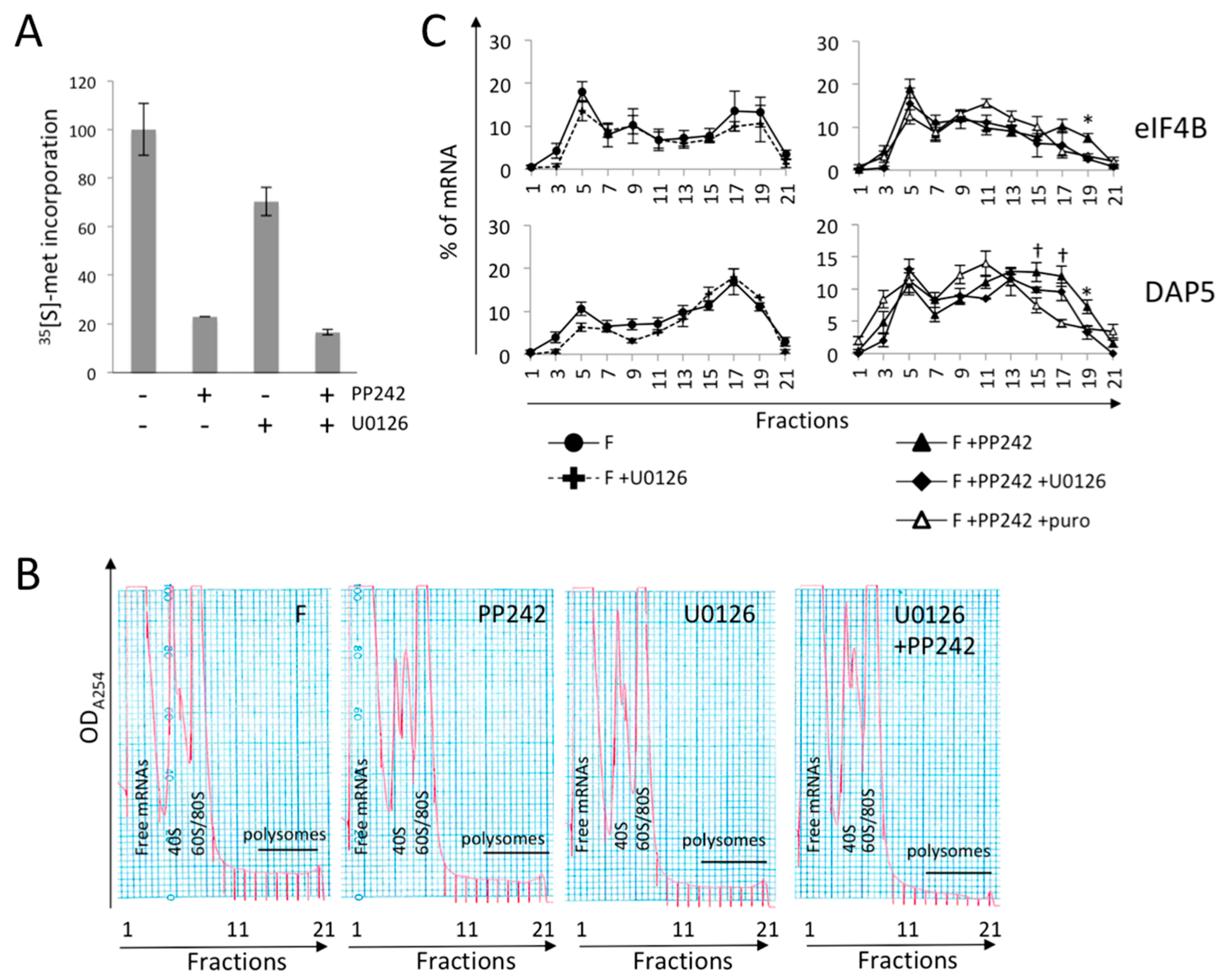
2.4. mTOR Pathway Differentially Impacts the Polysomal Recruitment of Initiation Factor mRNAs
3. Discussion
4. Materials and Methods
4.1. Handling and Treatment of Eggs and Embryos
4.2. Polysome Gradients and RT-PCR Analysis
4.3. In Vivo Protein Synthesis Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sonenberg, N.; Hinnebusch, A.G. New modes of translational control in development, behavior, and disease. Mol. Cell 2007, 28, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, R.J.; Hellen, C.U.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell. Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrick, W.C. eIF4F: A retrospective. J. Biol. Chem. 2015, 290, 24091–24099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozen, F.; Edery, I.; Meerovitch, K.; Dever, T.E.; Merrick, W.C.; Sonenberg, N. Bidirectional RNA helicase activity of eucaryotic translation initiation factors 4A and 4F. Mol. Cell. Biol. 1990, 10, 1134–1144. [Google Scholar] [CrossRef]
- Kahvejian, A.; Roy, G.; Sonenberg, N. The mRNA closed-loop model: The function of PABP and PABP-interacting proteins in mRNA translation. Cold Spring Harb. Symp. Quant. Biol. 2001, 66, 293–300. [Google Scholar] [CrossRef]
- Mader, S.; Lee, H.; Pause, A.; Sonenberg, N. The translation initiation factor eIF-4E binds to a common motif shared by the translation factor eIF-4 gamma and the translational repressors 4E-binding proteins. Mol. Cell. Biol. 1995, 15, 4990–4997. [Google Scholar] [CrossRef]
- Bah, A.; Vernon, R.M.; Siddiqui, Z.; Krzeminski, M.; Muhandiram, R.; Zhao, C.; Sonenberg, N.; Kay, L.E.; Forman-Kay, J.D. Folding of an intrinsically disordered protein by phosphorylation as a regulatory switch. Nature 2015, 519, 106–109. [Google Scholar] [CrossRef]
- Mamane, Y.; Petroulakis, E.; Martineau, Y.; Sato, T.A.; Larsson, O.; Rajasekhar, V.K.; Sonenberg, N. Epigenetic Activation of a Subset of mRNAs by eIF4E Explains Its Effects on Cell Proliferation. PLoS ONE 2007, 2, e242. [Google Scholar] [CrossRef]
- Modelska, A.; Turro, E.; Russell, R.; Beaton, J.; Sbarrato, T.; Spriggs, K.; Miller, J.; Gräf, S.; Provenzano, E.; Blows, F.; et al. The malignant phenotype in breast cancer is driven by eIF4A1-mediated changes in the translational landscape. Cell Death Dis. 2015, 6, e1603. [Google Scholar] [CrossRef]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Topisirovic, I. Signaling pathways involved in the regulation of mRNA translation. Mol. Cell. Biol. 2018, 38, MCB.00070-18. [Google Scholar] [CrossRef]
- Carracedo, A.; Ma, L.; Teruya-Feldstein, J.; Rojo, F.; Salmena, L.; Alimonti, A.; Egia, A.; Sasaki, A.T.; Thomas, G.; Kozma, S.C.; et al. Inhibition of mTORC1 leads to MAPK pathway activation through a PI3K-dependent feedback loop in human cancer. J. Clin. Investig. 2008, 118, 3065–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.; Cameron, A.; Jagus, R. Characterization of mammalian eIF4E-family members. Eur. J. Biochem. 2004, 271, 2189–2203. [Google Scholar] [CrossRef] [Green Version]
- Joshi, B.; Lee, K.; Maeder, D.L.; Jagus, R. Phylogenetic analysis of eIF4E-family members. BMC Evol. Biol. 2005, 5, 48. [Google Scholar] [CrossRef]
- Morales, J.; Mulner-Lorillon, O.; Cosson, B.; Morin, E.; Bellé, R.; Bradham, C.A.; Beane, W.S.; Cormier, P. Translational control genes in the sea urchin genome. Dev. Biol. 2006, 300, 293–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, M.; Ler, L.W.; Fabian, M.R.; Siddiqui, N.; Mullin, M.; Henderson, V.C.; Alain, T.; Fonseca, B.D.; Karashchuk, G.; Bennett, C.F.; et al. A novel 4EHP-GIGYF2 translational repressor complex is essential for mammalian development. Mol. Cell. Biol. 2012, 32, 3585–3593. [Google Scholar] [CrossRef] [PubMed]
- Uniacke, J.; Holterman, C.E.; Lachance, G.; Franovic, A.; Jacob, M.D.; Fabian, M.R.; Payette, J.; Holcik, M.; Pause, A.; Lee, S. An oxygen-regulated switch in the protein synthesis machinery. Nature 2012, 486, 126–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landon, A.L.; Muniandy, P.A.; Shetty, A.C.; Lehrmann, E.; Volpon, L.; Houng, S.; Zhang, Y.; Dai, B.; Peroutka, R.; Mazan-Mamczarz, K.; et al. MNKs act as a regulatory switch for eIF4E1 and eIF4E3 driven mRNA translation in DLBCL. Nat. Commun. 2014, 5, 5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, M.J.; Volpon, L.; Kornblatt, J.A.; Culjkovic-Kraljacic, B.; Baguet, A.; Borden, K.L.B. eIF4E3 acts as a tumor suppressor by utilizing an atypical mode of methyl-7-guanosine cap recognition. Proc. Natl. Acad. Sci. USA 2013, 110, 3877–3882. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, F. Versatility of the translational machinery during stress: Changing partners to keep dancing. Cell Res. 2012, 22, 1634–1636. [Google Scholar] [CrossRef] [PubMed]
- Henis-Korenblit, S.; Strumpf, N.L.; Goldstaub, D.; Kimchi, A. A novel form of DAP5 protein accumulates in apoptotic cells as a result of caspase cleavage and internal ribosome entry site-mediated translation. Mol. Cell. Biol. 2000, 20, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Liberman, N.; Gandin, V.; Svitkin, Y.V.; David, M.; Virgili, G.; Jaramillo, M.; Holcik, M.; Nagar, B.; Kimchi, A.; Sonenberg, N. DAP5 associates with eIF2 and eIF4AI to promote Internal Ribosome Entry Site driven translation. Nucl. Acids Res. 2015, 43, 3764–3775. [Google Scholar] [CrossRef] [PubMed]
- Marash, L.; Liberman, N.; Henis-Korenblit, S.; Sivan, G.; Reem, E.; Elroy-Stein, O.; Kimchi, A. DAP5 promotes cap-independent translation of Bcl-2 and CDK1 to facilitate cell survival during mitosis. Mol. Cell 2008, 30, 447–459. [Google Scholar] [CrossRef] [PubMed]
- de la Parra, C.; Ernlund, A.; Alard, A.; Ruggles, K.; Ueberheide, B.; Schneider, R.J. A widespread alternate form of cap-dependent mRNA translation initiation. Nat. Commun. 2018, 9, 3068. [Google Scholar] [CrossRef] [PubMed]
- Wigington, C.P.; Williams, K.R.; Meers, M.P.; Bassell, G.J.; Corbett, A.H. Poly(A) RNA-binding proteins and polyadenosine RNA: New members and novel functions. Wiley Interdiscip. Rev. RNA 2014, 5, 601–622. [Google Scholar] [CrossRef]
- Svitkin, Y.V.; Yanagiya, A.; Karetnikov, A.E.; Alain, T.; Fabian, M.R.; Khoutorsky, A.; Perreault, S.; Topisirovic, I.; Sonenberg, N. Control of translation and miRNA-dependent repression by a novel poly(A) binding protein, hnRNP-Q. PLoS Biol. 2013, 11, e1001564. [Google Scholar] [CrossRef]
- Lee, K.-H.; Woo, K.-C.; Kim, D.-Y.; Kim, T.-D.; Shin, J.; Park, S.M.; Jang, S.K.; Kim, K.-T. Rhythmic interaction between Period1 mRNA and hnRNP Q leads to circadian time-dependent translation. Mol. Cell. Biol. 2012, 32, 717–728. [Google Scholar] [CrossRef]
- Kim, D.-Y.; Kim, W.; Lee, K.-H.; Kim, S.-H.; Lee, H.-R.; Kim, H.-J.; Jung, Y.; Choi, J.-H.; Kim, K.-T. hnRNP Q regulates translation of p53 in normal and stress conditions. Cell Death Differ. 2012, 20, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Epel, D. Protein synthesis in sea urchin eggs: A “late” response to fertilization. Proc. Natl. Acad. Sci. USA 1967, 57, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Wagenaar, E.B. The timing of synthesis of proteins required for mitosis in the cell cycle of the sea urchin embryo. Exp. Cell Res. 1983, 144, 393–403. [Google Scholar] [CrossRef]
- Sodergren, E.; Weinstock, G.M.; Davidson, E.H.; Cameron, R.A.; Gibbs, R.A.; Angerer, R.C.; Angerer, L.M.; Arnone, M.I.; Burgess, D.R.; Burke, R.D.; et al. The genome of the sea urchin Strongylocentrotus purpuratus. Science 2006, 314, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Malkin, L.I.; Gross, P.R.; Romanoff, P. Polyribosomal protein synthesis in fertilized sea urchin eggs: The effect of Actinomycin treatment. Dev. Biol. 1964, 10, 378–394. [Google Scholar] [CrossRef]
- Davidson, E.H. The nature and function of maternal transcripts. In Gene Activity in Early Development; Academic Press: Cambridge, MA, USA, 1986; pp. 46–125. [Google Scholar]
- Winkler, M.M.; Nelson, E.M.; Lashbrook, C.; Hershey, J.W. Multiple levels of regulation of protein synthesis at fertilization in sea urchin eggs. Dev. Biol. 1985, 107, 290–300. [Google Scholar] [CrossRef]
- Jagus, R.; Huang, W.I.; Hansen, L.J.; Wilson, M.A. Changes in rates of protein synthesis and eukaryotic initiation factor-4 inhibitory activity in cell-free translation systems of sea urchin eggs and early cleavage stage embryos. J. Biol. Chem. 1992, 267, 15530–15536. [Google Scholar] [PubMed]
- Lopo, A.; Lashbrook, C.; Hershey, J. Characterization of translation systems in vitro from three developmental stages of Strongylocentrotus purpuratus. Biochem. J. 1989, 258, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.I.; Hansen, L.J.; Merrick, W.C.; Jagus, R. Inhibitor of eukaryotic initiation factor 4F activity in unfertilized sea urchin eggs. Proc. Natl. Acad. Sci. USA 1987, 84, 6359–6363. [Google Scholar] [CrossRef]
- Cormier, P.; Pyronnet, S.; Morales, J.; Mulner-Lorillon, O.; Sonenberg, N.; Belle, R. eIF4E association with 4E-BP decreases rapidly following fertilization in sea urchin. Dev. Biol. 2001, 232, 275–283. [Google Scholar] [CrossRef]
- Salaun, P.; Pyronnet, S.; Morales, J.; Mulner-Lorillon, O.; Bellé, R.; Sonenberg, N.; Cormier, P. eIF4E/4E-BP dissociation and 4E-BP degradation in the first mitotic division of the sea urchin embryo. Dev. Biol. 2003, 255, 428–439. [Google Scholar] [CrossRef]
- Oulhen, N.; Boulben, S.; Bidinosti, M.; Morales, J.; Cormier, P.; Cosson, B. A variant mimicking hyperphosphorylated 4E-BP inhibits protein synthesis in a sea urchin cell-free, cap-dependent translation system. PLoS ONE 2009, 4, e5070. [Google Scholar] [CrossRef]
- Oulhen, N.; Salaun, P.; Cosson, B.; Cormier, P.; Morales, J. After fertilization of sea urchin eggs, eIF4G is post-translationally modified and associated with the cap-binding protein eIF4E. J. Cell Sci. 2007, 120, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Cormier, P.; Chassé, H.; Cosson, B.; Mulner-Lorillon, O.; Morales, J. Translational control in echinoderms: The calm before the storm. In Evolution of the Protein Synthesis Machinery and Its Regulation; Hernández, G., Jagus, R., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 413–432. ISBN 978-3-319-39468-8. [Google Scholar]
- Chassé, H.; Aubert, J.; Boulben, S.; Le Corguillé, G.; Corre, E.; Cormier, P.; Morales, J. Translatome analysis at the egg-to-embryo transition in sea urchin. Nucl. Acids Res. 2018, 46, 4607–4621. [Google Scholar] [CrossRef] [PubMed]
- Hoang, B.; Benavides, A.; Shi, Y.; Yang, Y.; Frost, P.; Gera, J.; Lichtenstein, A. The PP242 mammalian target of rapamycin (mTOR) inhibitor activates extracellular signal-regulated kinase (ERK) in multiple myeloma cells via a target of rapamycin complex 1 (TORC1)/eukaryotic translation initiation factor 4E (eIF-4E)/RAF pathway. J. Biol. Chem. 2012, 287, 21796–21805. [Google Scholar] [CrossRef]
- Chassé, H.; Boulben, S.; Costache, V.; Cormier, P.; Morales, J. Analysis of translation using polysome profiling. Nucl. Acids Res. 2017, 45, e15. [Google Scholar] [CrossRef] [PubMed]
- Robalino, J.; Joshi, B.; Fahrenkrug, S.C.; Jagus, R. Two zebrafish eIF4E family members are differentially expressed and functionally divergent. J. Biol. Chem. 2004, 279, 10532–10541. [Google Scholar] [CrossRef] [PubMed]
- Chassé, H.; Mulner-Lorillon, O.; Boulben, S.; Glippa, V.; Morales, J.; Cormier, P. Cyclin B translation depends on mTOR activity after fertilization in sea urchin embryos. PLoS ONE 2016, 11, e0150318. [Google Scholar] [CrossRef]
- Laurent, S.; Richard, A.; Mulner-Lorillon, O.; Morales, J.; Flament, D.; Glippa, V.; Bourdon, J.; Gosselin, P.; Siegel, A.; Cormier, P.; et al. Modelization of the regulation of protein synthesis following fertilization in sea urchin shows requirement of two processes: A destabilization of eIF4E:4E-BP complex and a great stimulation of the 4E-BP-degradation mechanism, both rapamycin-sensitive. Front. Genet. 2014, 5. [Google Scholar] [CrossRef]
- Mulner-Lorillon, O.; Chassé, H.; Morales, J.; Bellé, R.; Cormier, P. MAPK/ERK activity is required for the successful progression of mitosis in sea urchin embryos. Dev. Biol. 2017, 421, 194–203. [Google Scholar] [CrossRef]
- Dinkova, T.D.; Keiper, B.D.; Korneeva, N.L.; Aamodt, E.J.; Rhoads, R.E. Translation of a Small Subset of Caenorhabditis elegans mRNAs Is Dependent on a Specific Eukaryotic Translation Initiation Factor 4E Isoform. Mol. Cell. Biol. 2005, 25, 100–113. [Google Scholar] [CrossRef]
- Cho, P.F.; Gamberi, C.; Cho-Park, Y.A.; Cho-Park, I.B.; Lasko, P.; Sonenberg, N. Cap-dependent translational inhibition establishes two opposing morphogen gradients in Drosophila embryos. Curr. Biol. 2006, 16, 2035–2041. [Google Scholar] [CrossRef] [PubMed]
- Coffman, J.A.; McCarthy, J.J.; Dickey-Sims, C.; Robertson, A.J. Oral-aboral axis specification in the sea urchin embryo: II. Mitochondrial distribution and redox state contribute to establishing polarity in Strongylocentrotus purpuratus. Dev. Biol. 2004, 273, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Coffman, J.A.; Denegre, J.M. Mitochondria, redox signaling and axis specification in metazoan embryos. Dev. Biol. 2007, 308, 266–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.J.D.; Wang, M.; Audas, T.E.; Kwon, D.; Carlsson, S.K.; Timpano, S.; Evagelou, S.L.; Brothers, S.; Gonzalgo, M.L.; Krieger, J.R.; et al. Systemic reprogramming of translation efficiencies on oxygen stimulus. Cell Rep. 2016, 14, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Kelly, N.J.; Varga, J.F.A.; Specker, E.J.; Romeo, C.M.; Coomber, B.L.; Uniacke, J. Hypoxia activates cadherin-22 synthesis via eIF4E2 to drive cancer cell migration, invasion and adhesion. Oncogene 2018, 37, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Le Bouffant, R.; Cormier, P.; Mulner-lorillon, O.; Belle, R. Hypoxia and DNA-damaging agent bleomycin both increase the cellular level of the protein 4E-BP. J. Cell. Biochem. 2006, 99, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Rom, E.; Kim, H.C.; Gingras, A.C.; Marcotrigiano, J.; Favre, D.; Olsen, H.; Burley, S.K.; Sonenberg, N. Cloning and characterization of 4EHP, a novel mammalian eIF4E-related cap-binding protein. J. Biol. Chem. 1998, 273, 13104–13109. [Google Scholar] [CrossRef]
- Pyronnet, S.; Imataka, H.; Gingras, A.C.; Fukunaga, R.; Hunter, T.; Sonenberg, N. Human eukaryotic translation initiation factor 4G (eIF4G) recruits MNK1 to phosphorylate eIF4E. EMBO J. 1999, 18, 270–279. [Google Scholar] [CrossRef]
- Lee, A.S.Y.; Kranzusch, P.J.; Doudna, J.A.; Cate, J.H.D. eIF3d is an mRNA cap-binding protein required for specialized translation initiation. Nature 2016, 536, 96–99. [Google Scholar] [CrossRef]
- Liberman, N.; Marash, L.; Kimchi, A. The translation initiation factor DAP5 is a regulator of cell survival during mitosis. Cell Cycle 2009, 8, 204–209. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Transcript # | FPKM * | Primer Sequence | Product Length (bp) | Efficiency (R2) |
|---|---|---|---|---|---|
| eIF4A | comp73316_c0_seq1 | 59.53 | TGGTCAAGAAGGAAGAAC CGTCTCATACAAGTCACA | 103 | 0.990 |
| eIF4B | comp78411_c2_seq1 | 94.72 | GGAGGAGCAAAGCCTGTAGA ACGCGTTCTGCTTTCTCTTC | 200 | 0.992 |
| eIF4E1 | comp52193_c0_seq1 | <1 | GGTGGAAGGTGGCTCATAGG TCTTTCCTCCAGTCCCCTGT | 191 | 0.996 |
| eIF4E2 | comp72071_c0_seq2 | 30 | TATGGTCGGAGAGGAGAT ATTATTATCGCTGGCTGTG | 128 | 0.989 |
| eIF4E3 | comp75131_c0_seq1 | 20.3 | GTAAAGCCCCTATGGGAAGA TTGGTGCCCCTAATGCTTAC | 185 | 0.996 |
| eIF4G | comp69782_c0_seq2 | 2.6 | CCATGTTGAGTGAGGATGCG ACCTTCTCCTGGGATCCTCT | 225 | 0.979 |
| DAP5 | comp79103_c1_seq1 | 87.1 | AGACGAGCAGGACCAGAGAG GTCGGCCTACAGTGGTGATT | 205 | 0.994 |
| PABP | comp73981_c0_seq1 | 6.5 | GCACCTCAAGTTCGAGTTGG TGGTCTGGAAGTTAGGCTGG | 201 | 0.992 |
| hnRNP Q | comp75304_c0_seq1 | 1439.6 | GAGGAGATGAACGGCAGAGA GTAGCCTCCAAAGTCCCTGT | 230 | 0.999 |
| 4E-BP | comp78493_c0_seq1 | 13.2 | CCCATGATTACAGCACTAC GGAAGTTACGGTCATAGATG | 83 | 0.996 |
| Function | Protein | Interactions | Role and References | |||||
|---|---|---|---|---|---|---|---|---|
| Cap | eIF4G | 4E-BP | ||||||
| Cap-binding proteins | eIF4E1 | +++ | +++ | +++ | Canonical eIF [3,15,48] | |||
| eIF4E2 | + | / | + | Selective translation [15,18,19,20,21,48] | ||||
| eIF4E3 | + | + | / | |||||
| Scaffolding protein | eIF4E1 | PABP | eIF3 | eIF4A | ||||
| eIF4G | +++ | +++ | + | + | Canonical eIF [3] | |||
| DAP5 | / | / | + | + | Selective translation [24,25,26] | |||
| Poly(A)-binding protein | poly(A) | eIF4G | ||||||
| PABP | +++ | +++ | Canonical eIF [3] | |||||
| hnRNP Q | + | +++ | Selective translation [28,29,30] | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chassé, H.; Boulben, S.; Cormier, P.; Morales, J. Translational Control of Canonical and Non-Canonical Translation Initiation Factors at the Sea Urchin Egg to Embryo Transition. Int. J. Mol. Sci. 2019, 20, 626. https://doi.org/10.3390/ijms20030626
Chassé H, Boulben S, Cormier P, Morales J. Translational Control of Canonical and Non-Canonical Translation Initiation Factors at the Sea Urchin Egg to Embryo Transition. International Journal of Molecular Sciences. 2019; 20(3):626. https://doi.org/10.3390/ijms20030626
Chicago/Turabian StyleChassé, Héloïse, Sandrine Boulben, Patrick Cormier, and Julia Morales. 2019. "Translational Control of Canonical and Non-Canonical Translation Initiation Factors at the Sea Urchin Egg to Embryo Transition" International Journal of Molecular Sciences 20, no. 3: 626. https://doi.org/10.3390/ijms20030626
APA StyleChassé, H., Boulben, S., Cormier, P., & Morales, J. (2019). Translational Control of Canonical and Non-Canonical Translation Initiation Factors at the Sea Urchin Egg to Embryo Transition. International Journal of Molecular Sciences, 20(3), 626. https://doi.org/10.3390/ijms20030626