Aspalathin-Enriched Green Rooibos Extract Reduces Hepatic Insulin Resistance by Modulating PI3K/AKT and AMPK Pathways

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
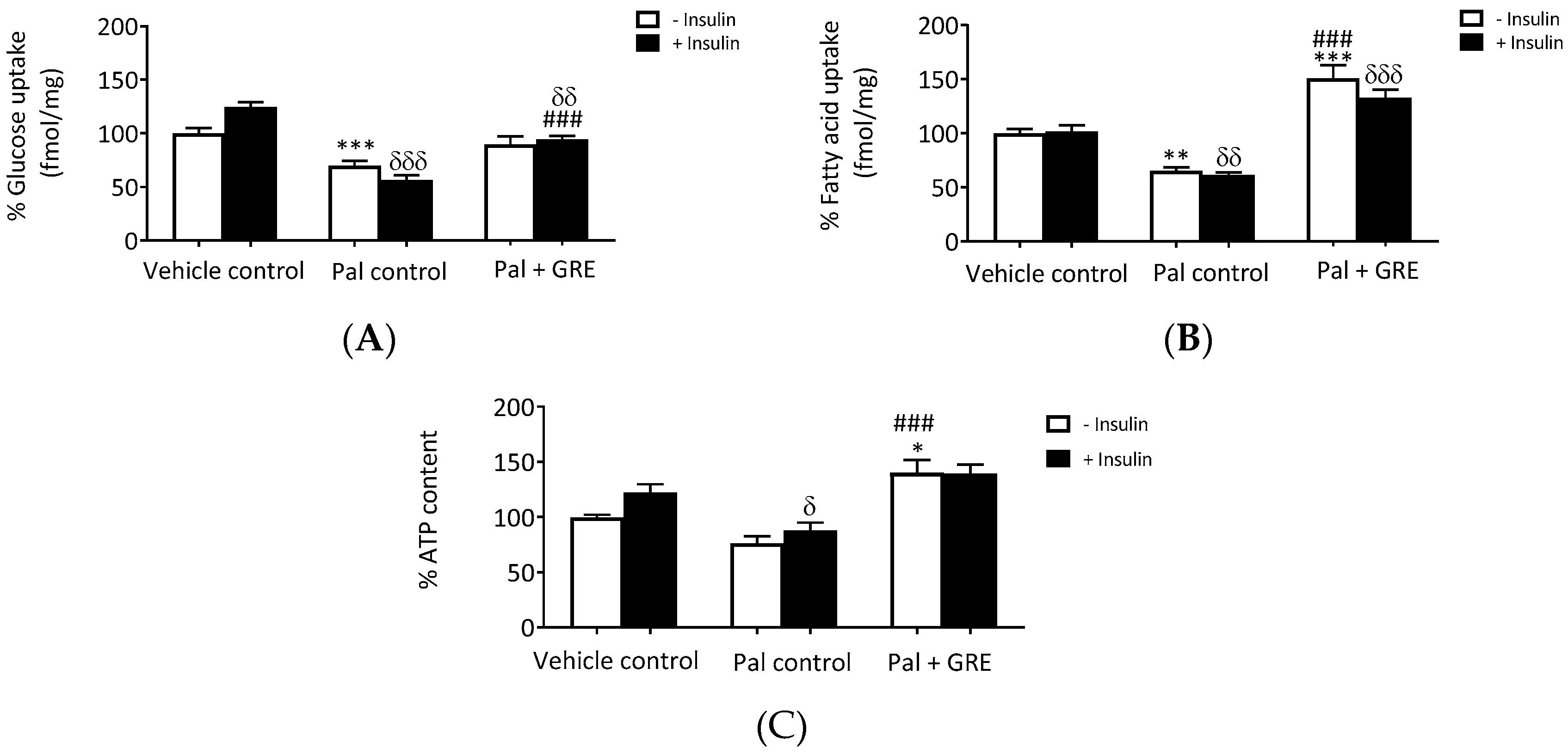
2.1. GRE Treatment Attenuated Palmitate-Induced Insulin Resistance in C3A Liver Cells
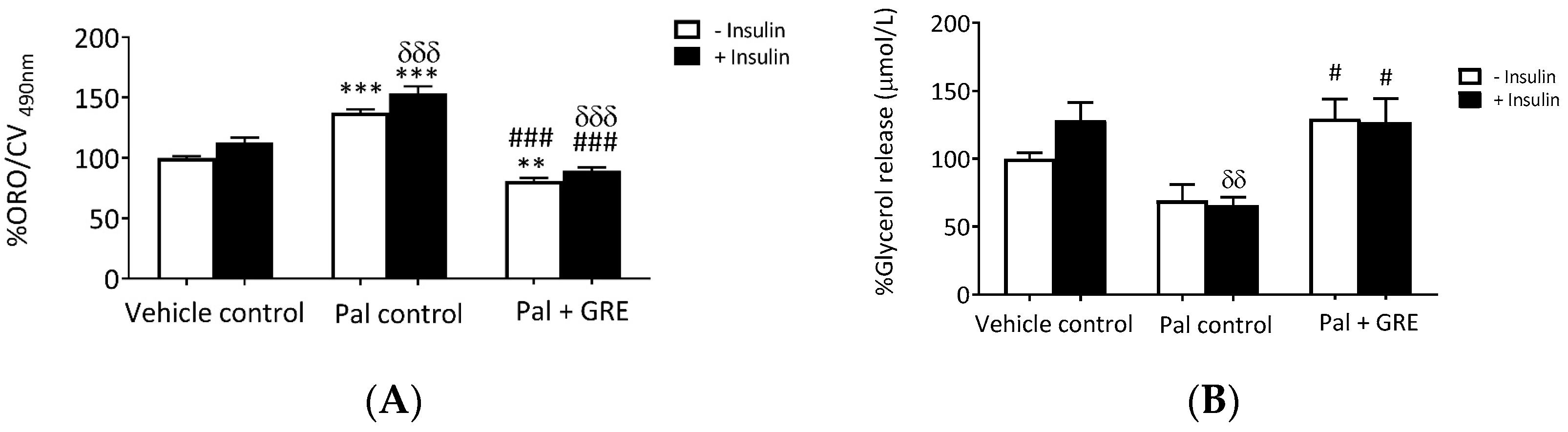
2.2. GRE Reduced Lipid Accumulation and Increased Lipolysis
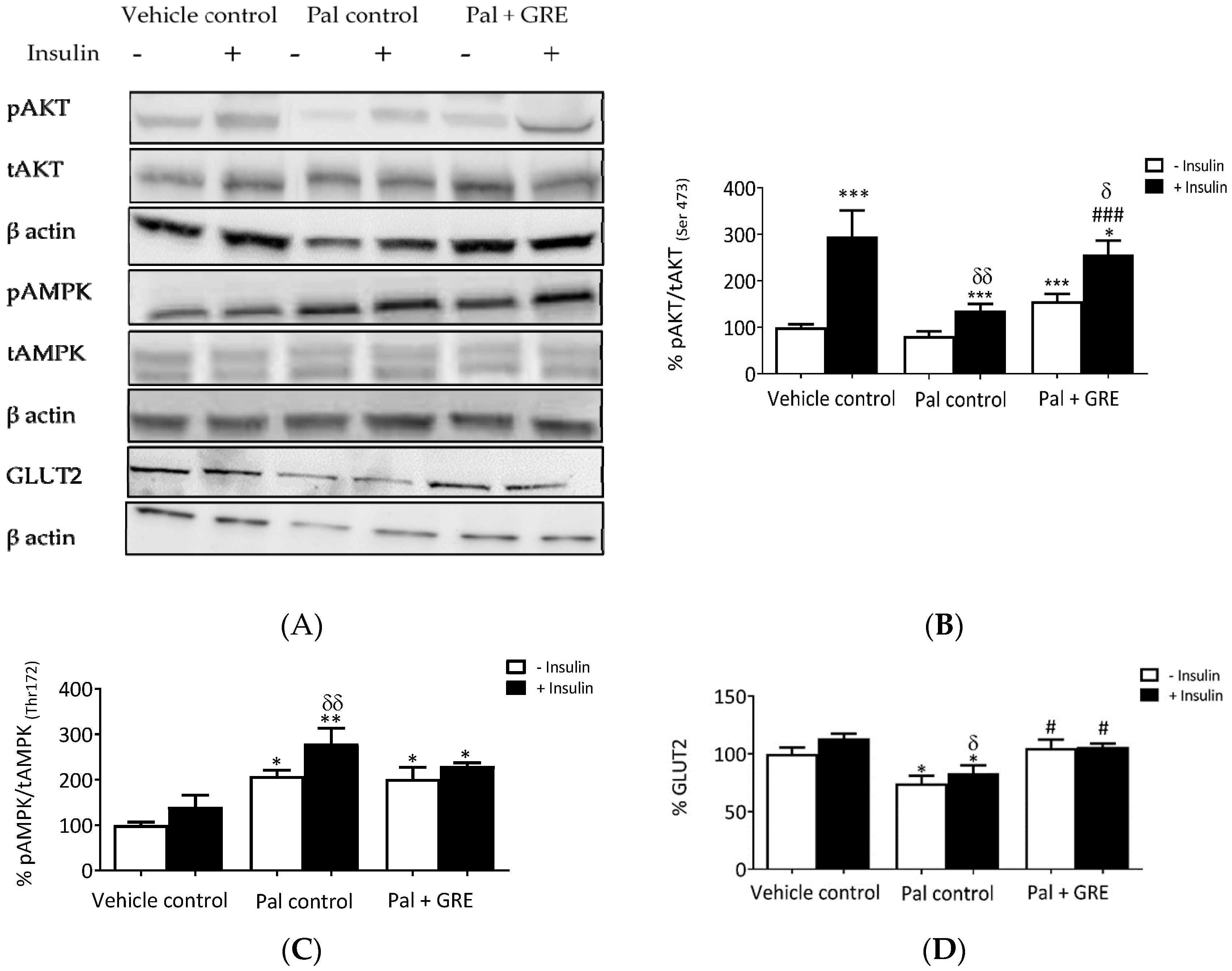
2.3. GRE Prevented Palmitate-Induced Insulin Resistance through Activation of AKT and AMPK Pathway In Vitro
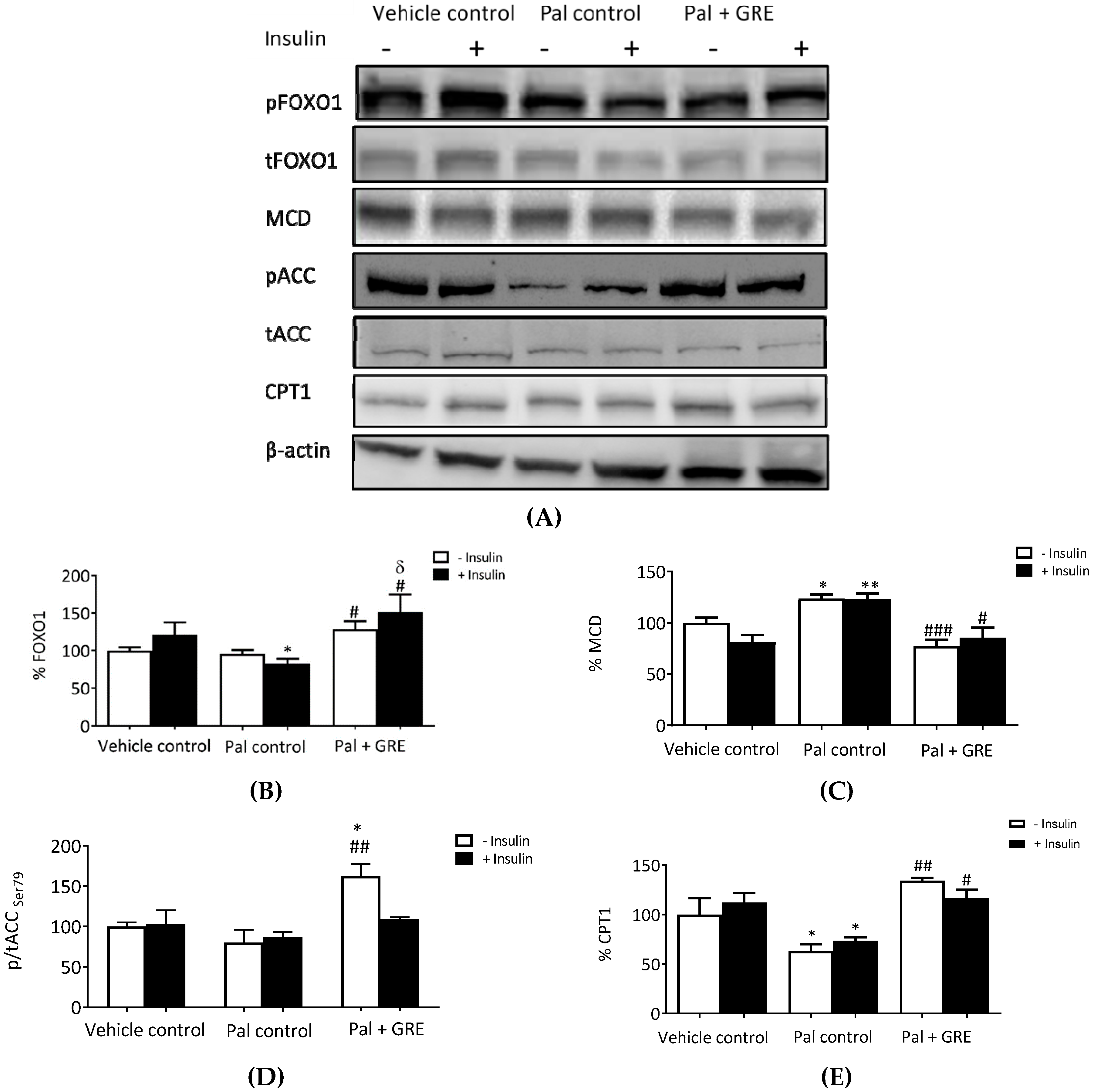
2.4. GRE Stimulated Fatty Acid Oxidation-Associated Gene Expression
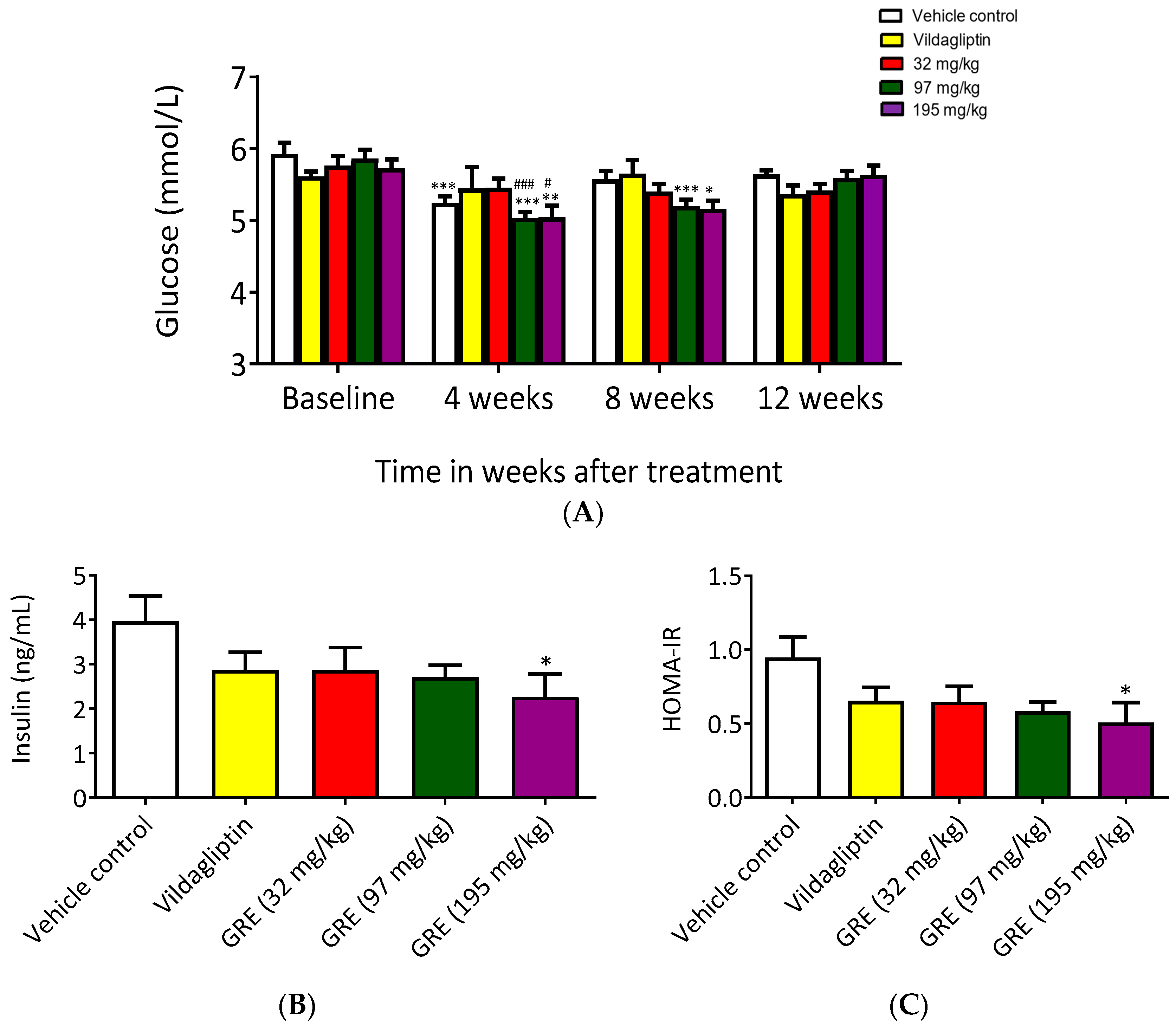
2.5. GRE Partially Enhanced Metabolic Profile in OBIR Rat Model
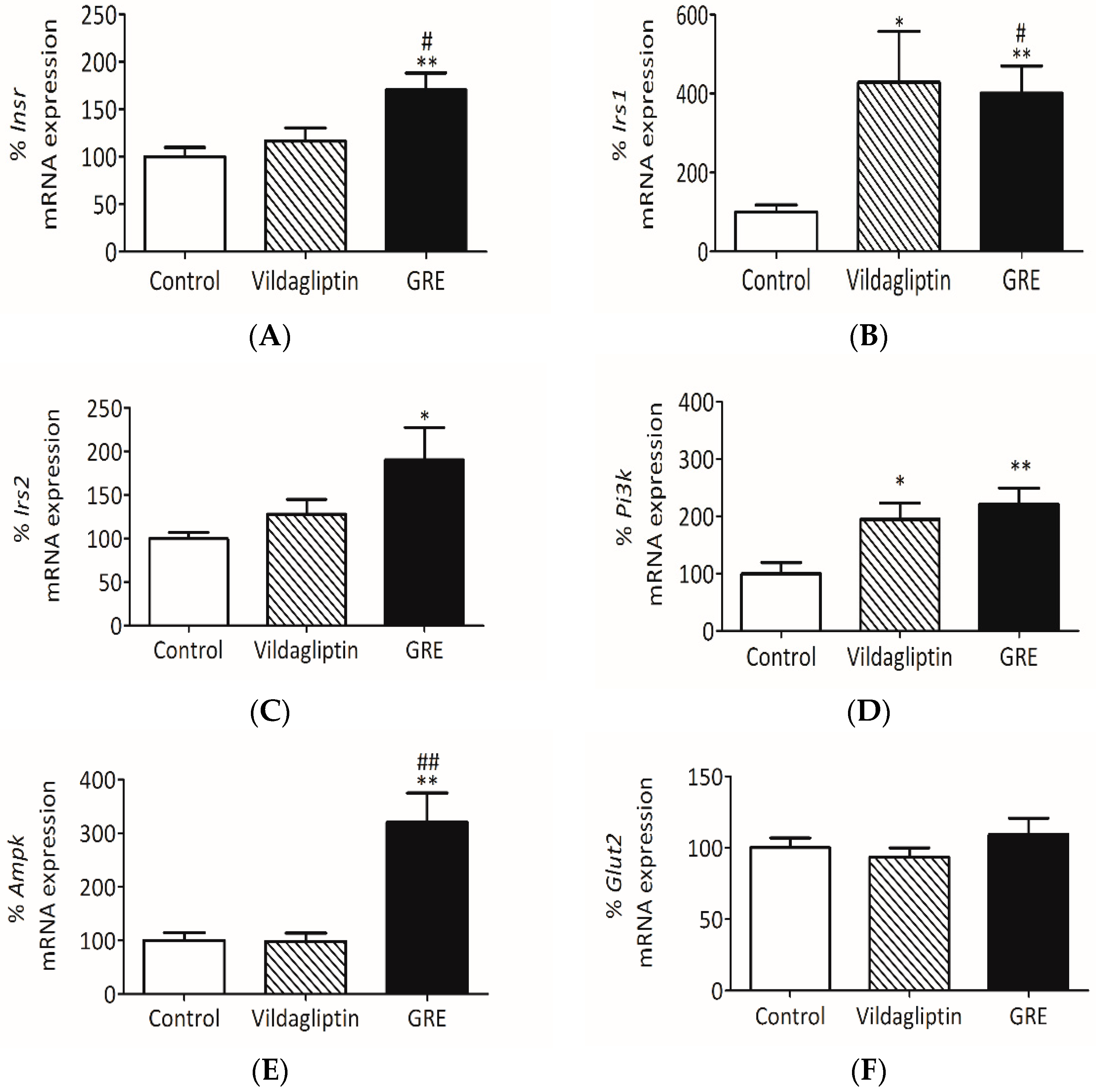
2.6. GRE Improved Insulin Signaling In Vivo
3. Discussion
4. Materials and Methods
4.1. Reagents and Kits
4.2. Antibodies and Gene Expression Primers
4.3. In Vitro Experiments: Cell Culture, Induction of Insulin Resistance, and Treatment with GRE
4.4. Measurement of 2-Deoxy-[3H]-d-Glucose and 14C Palmitate Uptake and ATP Contents
4.5. Oil Red O Staining
4.6. Free Glycerol Determination as a Measure of Lypolysis
4.7. Western Blot Analysis
4.8. In Vivo Experiments: Animal Housing and Ethical Clearance
4.9. Establishment of an OBIR Rat Model and Administration of GRE
4.10. Parameters Measured in OBIR Rats
4.11. RNA Extraction and Quantitative Real-Time PCR
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| AMP | Adenosine monophosphate |
| AMPK | AMP-activated protein kinase |
| ATP | Adenosine triphosphate |
| CPT1 | Carnitine palmitoyl transferase 1 |
| DMEM | Dulbecco Modified Eagle’s Medium |
| DMSO | Dimethyl sulfoxide |
| ECRA | Ethics Committee for Research on Animals |
| EMEM | Eagle’s Minimum Essential Medium |
| FOXO1 | Forkhead box protein O1 |
| GLUT | Glucose transporter |
| GRE | Green rooibos extract |
| GSK3 | Glycogen synthase kinase 3 |
| HOMA-IR | Homeostatic Model Assessment of Insulin Resistance |
| IRS | Insulin receptor substrate |
| LCFA | Long chain free fatty acids |
| MCD | Malonyl-CoA decarboxylase |
| OBIR | Obese insulin resistant |
| OGTTs | Oral glucose tolerance tests |
| PI3K | Phosphatidylinositol-4,5-bisphosphate 3-kinase |
| SAMRC | South African Medical Research Council |
References
- Sears, B.; Perry, M. The role of fatty acids in insulin resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.W.; Heden, T.D.; Matthew Morris, E.; Fritsche, K.L.; Vieira-Potter, V.J.; Thyfault, J.P. High-fat diet alters serum fatty acid profiles in obesity prone rats: Implications for in vitro studies. Lipids 2015, 50, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Walton, R.G.; Zhu, B.; Unal, R.; Spencer, M.; Sunkara, M.; Morris, A.J.; Charnigo, R.; Katz, W.S.; Daugherty, A.; Howatt, D.A.; et al. Increasing adipocyte lipoprotein lipase improves glucose metabolism in high fat diet-induced obesity. J. Biol. Chem. 2015, 290, 11547–11556. [Google Scholar] [CrossRef] [PubMed]
- Gruben, N.; Shiri-Sverdlov, R.; Koonen, D.P.Y.; Hofker, M.H. Nonalcoholic fatty liver disease: A main driver of insulin resistance or a dangerous liaison? Biochim. Biophys. Acta 2014, 1842, 2329–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jornayvaz, F.R.; Shulman, G.I. Diacylglycerol activation of protein kinase Cε and hepatic insulin resistance. Cell Metab. 2012, 15, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Birkenfeld, A.L.; Shulman, G.I. Nonalcoholic fatty liver disease, hepatic insulin resistance, and type 2 Diabetes. Hepatology 2014, 59, 713–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monsénégo, J.; Mansouri, A.; Akkaoui, M.; Lenoir, V.; Esnous, C.; Fauveau, V.; Tavernier, V.; Girard, J.; Prip-Buus, C. Enhancing liver mitochondrial fatty acid oxidation capacity in obese mice improves insulin sensitivity independently of hepatic steatosis. J. Hepatol. 2012, 56, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Han, S.; Kitamura, T.; Accili, D. Dual role of transcription factor FoxO1 in controlling hepatic insulin sensitivity and lipid metabolism. J. Clin. Investig. 2006, 116, 2464–2472. [Google Scholar] [CrossRef] [PubMed]
- Babu, P.V.; Liu, D.; Gilbert, E.R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 2013, 24, 1777–1789. [Google Scholar] [CrossRef] [Green Version]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Role of red grape polyphenols as antidiabetic agents. Integr. Med. Res. 2014, 3, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and glycemic control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Joubert, E.; de Beer, D. Rooibos (Aspalathus linearis) beyond the farm gate: From herbal tea to potential phytopharmaceutical. S. Afr. J. Bot. 2011, 77, 869–886. [Google Scholar] [CrossRef]
- Muller, C.J.F.; Joubert, E.; de Beer, D.; Sanderson, M.; Malherbe, C.J.; Fey, S.J.; Louw, J. Acute assessment of an aspalathin-enriched green rooibos (Aspalathus linearis) extract with hypoglycemic potential. Phytomedicine 2012, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Mazibuko, S.E.; Muller, C.J.; Joubert, E.; de Beer, D.; Johnson, R.; Opoku, A.R.; Louw, J. Amelioration of palmitate-induced insulin resistance in C2C12 muscle cells by rooibos (Aspalathus linearis). Phytomedicine 2013, 20, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, M.; Mazibuko, S.E.; Joubert, E.; de Beer, D.; Johnson, R.; Pheiffer, C.; Louw, J.; Muller, C.J. Effects of fermented rooibos (Aspalathus linearis) on adipocyte differentiation. Phytomedicine 2014, 21, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Muller, C.J.; Louw, J.; Joubert, E.; Salie, R.; Opoku, A.R.; Johnson, R. The cardioprotective effect of an aqueous extract of fermented rooibos (Aspalathus linearis) on cultured cardiomyocytes derived from diabetic rats. Phytomedicine 2014, 21, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Kamakura, R.; Son, M.J.; de Beer, D.; Joubert, E.; Miura, Y.; Yagasaki, K. Antidiabetic effect of green rooibos (Aspalathus linearis) extract in cultured cells and type 2 diabetic model KK-A(y) mice. Cytotechnology 2015, 67, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Orlando, P.; Chellan, N.; Muller, C.J.F.; Louw, J.; Chapman, C.C.; Joubert, E.; Tiano, L. Green rooibos extract improves plasma lipid profile and oxidative status in diabetic non-human primates. Free Radic. Biol. Med. 2017, 108, S97. [Google Scholar] [CrossRef]
- Son, M.J.; Minakawa, M.; Miura, Y.; Yagasaki, K. Aspalathin improves hyperglycemia and glucose intolerance in obese diabetic ob/ob mice. Eur. J. Nutr. 2013, 52, 1607–1619. [Google Scholar] [CrossRef]
- Mazibuko, S.E.; Joubert, E.; Johnson, R.; Louw, J.; Opoku, A.R.; Muller, C.J. Aspalathin improves glucose and lipid metabolism in 3T3-L1 adipocytes exposed to palmitate. Mol. Nutr. Food Res. 2015, 59, 2199–2208. [Google Scholar] [CrossRef]
- Najafian, M.; Najafian, B.; Najafian, Z. The Effect of aspalathin on levels of sugar and lipids in streptozotocin-induced diabetic and normal rats. Zahedan J. Res. Med. Sci. 2016, 18, e4963. [Google Scholar] [CrossRef]
- Johnson, R.; Dludla, P.; Joubert, E.; February, F.; Mazibuko, S.; Ghoor, S.; Muller, C.; Louw, J. Aspalathin, a dihydrochalcone C-glucoside, protects H9c2 cardiomyocytes against high glucose induced shifts in substrate preference and apoptosis. Mol. Nutr. Food Res. 2016, 60, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Joubert, E.; Muller, C.J.F.; Louw, J.; Johnson, R. Hyperglycemia-induced oxidative stress and heart disease-cardioprotective effects of rooibos flavonoids and phenylpyruvic acid-2-O-β-d-glucoside. Nutr. Metab. 2017, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.M.; Min, K.H.; Lee, W. Induction of miR-96 by dietary saturated fatty acids exacerbates hepatic insulin resistance through the suppression of INSR and IRS-1. PLoS ONE 2016, 11, e0169039. [Google Scholar] [CrossRef] [PubMed]
- Kanuri, B.N.; Kanshana, J.S.; Rebello, S.C.; Pathak, P.; Gupta, A.P.; Gayen, J.R.; Jagavelu, K.; Dikshit, M. Altered glucose and lipid homeostasis in liver and adipose tissue pre-dispose inducible NOS knockout mice to insulin resistance. Sci. Rep. 2017, 7, 41009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.T.; Chang, T.W.; Lee, M.S.; Lin, J.K. Suppression of free fatty acid-induced insulin resistance by phytopolyphenols in C2C12 mouse skeletal muscle cells. J. Agric. Food Chem. 2012, 60, 1059–1066. [Google Scholar] [CrossRef]
- Park, J.-Y.; Kim, Y.; Im, J.A.; Lee, H. Oligonol suppresses lipid accumulation and improves insulin resistance in a palmitate-induced in HepG2 hepatocytes as a cellular steatosis model. BMC Complement. Altern. Med. 2015, 15, 185. [Google Scholar] [CrossRef]
- Watt, M.J.; Steinberg, G.R.; Chen, Z.P.; Kemp, B.E.; Febbraio, M.A. Fatty acids stimulate AMP-activated protein kinase and enhance fatty acid oxidation in L6 myotubes. J. Physiol. 2006, 574, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.-Q.; Li, X.-Y.; Wang, L.; Feng, Z.-L.; Li, X.-F.; Wen, Y.-F.; Han, J.-X. Palmitate induces fat accumulation by activating C/EBPβ-mediated G0S2 expression in HepG2 cells. World J. Gastroenterol. 2017, 23, 7705–7715. [Google Scholar] [CrossRef]
- Li, Y.-C.; Qiao, J.-Y.; Wang, B.-Y.; Bai, M.; Shen, J.-D.; Cheng, Y.-X. Paeoniflorin Ameliorates Fructose-Induced Insulin Resistance and Hepatic Steatosis by Activating LKB1/AMPK and AKT Pathways. Nutrients 2018, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, M.D.; Galic, S.; Marcinko, K.; Sikkema, S.; Pulinilkunnil, T.; Chen, Z.-P.; Steinberg, G.R. Single phosphorylation sites in Acc1 and Acc2 regulate lipid homeostasis and the insulin-sensitizing effects of metformin. Nat. Med. 2013, 19, 1649–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ussher, J.R.; Fillmore, N.; Keung, W.; Zhang, L.; Mori, J.; Sidhu, V.K.; Fukushima, A.; Gopal, K.; Lopaschuk, D.G.; Wagg, C.S.; et al. Genetic and pharmacological inhibition of malonyl CoA decarboxylase does not exacerbate age-related insulin resistance in mice. Diabetes 2016, 65, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Sparks, J.D.; Dong, H.H. FoxO1 and hepatic lipid metabolism. Curr. Opin. Lipidol. 2009, 20, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, E.L.; Banko, M.R.; Brunet, A. AMP-activated protein kinase and FoxO transcription factors in dietary restriction-induced longevity. Ann. N. Y. Acad. Sci. 2009, 1170, 688–692. [Google Scholar] [CrossRef]
- Guo, S. Insulin signaling, resistance, and the metabolic syndrome: Insights from mouse models into disease mechanisms. J. Endocrinol. 2014, 220, T1–T23. [Google Scholar] [CrossRef]
- Mackenzie, R.W.; Elliott, B.T. Akt/PKB activation and insulin signaling: A novel insulin signaling pathway in the treatment of type 2 diabetes. Diabetes Metab. Syndr. Obes. 2014, 7, 55–64. [Google Scholar] [CrossRef]
- Kwon, Y.; Song, P.; Yoon, J.H.; Ghim, J.; Kim, D.; Kang, B.; Lee, T.G.; Kim, J.A.; Choi, J.K.; Youn, I.K.; et al. Xanthene derivatives increase glucose utilization through activation of LKB1-dependent AMP-activated protein kinase. PLoS ONE 2014, 9, e108771. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, C.W.; Park, S.M.; Jegal, K.H.; Kim, J.K.; Park, C.A.; Cho, I.J.; Jung, D.H.; An, W.G.; Ku, S.K.; et al. Activation of AMPK by Buddleja officinalis Maxim. Flower extract contributes to protecting hepatocytes from oxidative stress. Evid. Based Complement. Altern. Med. 2017, 2017, 9253462. [Google Scholar] [CrossRef]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.B.; Yang, Y.R.; Mo, Z.J.; Ding, Y.; Jiang, W.J. Silibinin improves palmitate-induced insulin resistance in C2C12 myotubes by attenuating IRS-1/PI3K/Akt pathway inhibition. Braz. J. Med. Biol. Res. 2015, 48, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.L.; Yang, Y.R.; Mo, Z.J.; Ding, Y.; Jiang, W.J. Timosaponin B-II ameliorates palmitate-induced insulin resistance and inflammation via IRS-1/PI3K/Akt and IKK/NF-κB Pathways. Am. J. Chin. Med. 2016, 44, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Sadi, G.; Pektaş, M.B.; Koca, H.B.; Tosun, M.; Koca, T. Resveratrol improves hepatic insulin signaling and reduces the inflammatory response in streptozotocin-induced diabetes. Gene 2015, 570, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Herrera, I.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Cocoa flavonoids improve insulin signalling and modulate glucose production via AKT and AMPK in HepG2 cells. Mol. Nutr. Food Res. 2013, 57, 974–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Merwe, J.; de Beer, D.; Joubert, E.; Gelderblom, W. Short-term and sub-chronic dietary exposure to aspalathin-enriched green rooibos (Aspalathus linearis) extract affects rat liver function and antioxidant status. Molecules 2015, 20, 22674–22690. [Google Scholar] [CrossRef] [PubMed]
- 4Wang, C.; Jiang, J.D.; Wu, W.; Kong, W.J. The compound of mangiferin-berberine salt has potent activities in modulating lipid and glucose metabolisms in HepG2 Cells. Biomed. Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Y.; Qin, Y.; Liu, L.; Wan, J.; Zou, L.; Zhang, Q.; Zhu, J.; Mi, M. Ampelopsin improves insulin resistance by activating PPARγ and subsequently up-regulating FGF21-AMPK signaling pathway. PLoS ONE 2016, 11, e0159191. [Google Scholar] [CrossRef] [PubMed]
- Smit, S.; Johnson, R.; Van Vuuren, M.; Huisamen, B. Myocardial glucose clearance by aspalathin treatment in young, mature, and obese insulin-resistant rats. Planta Med. 2018, 84, 75–82. [Google Scholar] [CrossRef]
- Johnson, R.; Dludla, P.V.; Muller, C.J.; Huisamen, B.; Essop, M.F.; Louw, J. The transcription profile unveils the cardioprotective effect of aspalathin against lipid toxicity in an in vitro H9c2 model. Molecules 2017, 22, 219. [Google Scholar] [CrossRef]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Krygsman, A.; Roux, C.R.; Muller, C.; Louw, J. Development of glucose intolerance in Wistar rats fed low and moderate fat diets differing in fatty acid profile. Exp. Clin. Endocrinol. Diabetes 2010, 118, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Joubert, E.; Beelders, T.; de Beer, D.; Malherbe, C.J.; de Villiers, A.J.; Sigge, G.O. Variation in phenolic content and antioxidant activity of fermented rooibos herbal tea infusions: Role of production season and quality grade. J. Agric. Food Chem. 2012, 60, 9171–9179. [Google Scholar] [CrossRef] [PubMed]
- Mathews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 2012, 28, 412–419. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazibuko-Mbeje, S.E.; Dludla, P.V.; Roux, C.; Johnson, R.; Ghoor, S.; Joubert, E.; Louw, J.; Opoku, A.R.; Muller, C.J.F. Aspalathin-Enriched Green Rooibos Extract Reduces Hepatic Insulin Resistance by Modulating PI3K/AKT and AMPK Pathways. Int. J. Mol. Sci. 2019, 20, 633. https://doi.org/10.3390/ijms20030633
Mazibuko-Mbeje SE, Dludla PV, Roux C, Johnson R, Ghoor S, Joubert E, Louw J, Opoku AR, Muller CJF. Aspalathin-Enriched Green Rooibos Extract Reduces Hepatic Insulin Resistance by Modulating PI3K/AKT and AMPK Pathways. International Journal of Molecular Sciences. 2019; 20(3):633. https://doi.org/10.3390/ijms20030633
Chicago/Turabian StyleMazibuko-Mbeje, Sithandiwe E., Phiwayinkosi V. Dludla, Candice Roux, Rabia Johnson, Samira Ghoor, Elizabeth Joubert, Johan Louw, Andy R. Opoku, and Christo J. F. Muller. 2019. "Aspalathin-Enriched Green Rooibos Extract Reduces Hepatic Insulin Resistance by Modulating PI3K/AKT and AMPK Pathways" International Journal of Molecular Sciences 20, no. 3: 633. https://doi.org/10.3390/ijms20030633
APA StyleMazibuko-Mbeje, S. E., Dludla, P. V., Roux, C., Johnson, R., Ghoor, S., Joubert, E., Louw, J., Opoku, A. R., & Muller, C. J. F. (2019). Aspalathin-Enriched Green Rooibos Extract Reduces Hepatic Insulin Resistance by Modulating PI3K/AKT and AMPK Pathways. International Journal of Molecular Sciences, 20(3), 633. https://doi.org/10.3390/ijms20030633