Exogenous Application of Phytohormones Promotes Growth and Regulates Expression of Wood Formation-Related Genes in Populus simonii × P. nigra

Abstract
:1. Introduction
2. Results
2.1. Exogenous Phytohormones Promote Plant Growth and Cellulose Synthesis
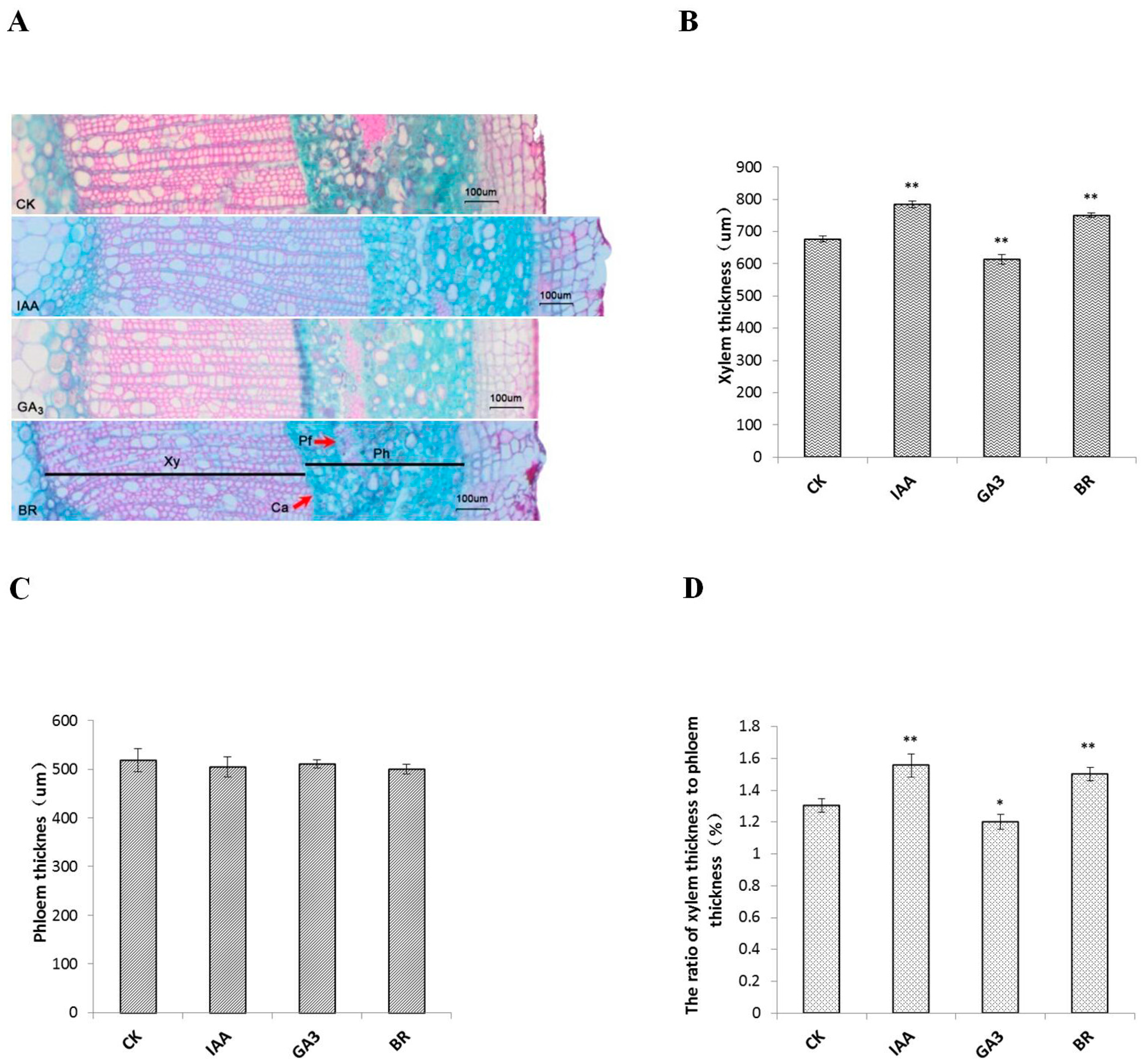
2.2. Phytohormones Promote Xylem Differentiation in Populus
2.3. Expression Profiles of Genes in Response to IAA, GA3, and BR Treatments
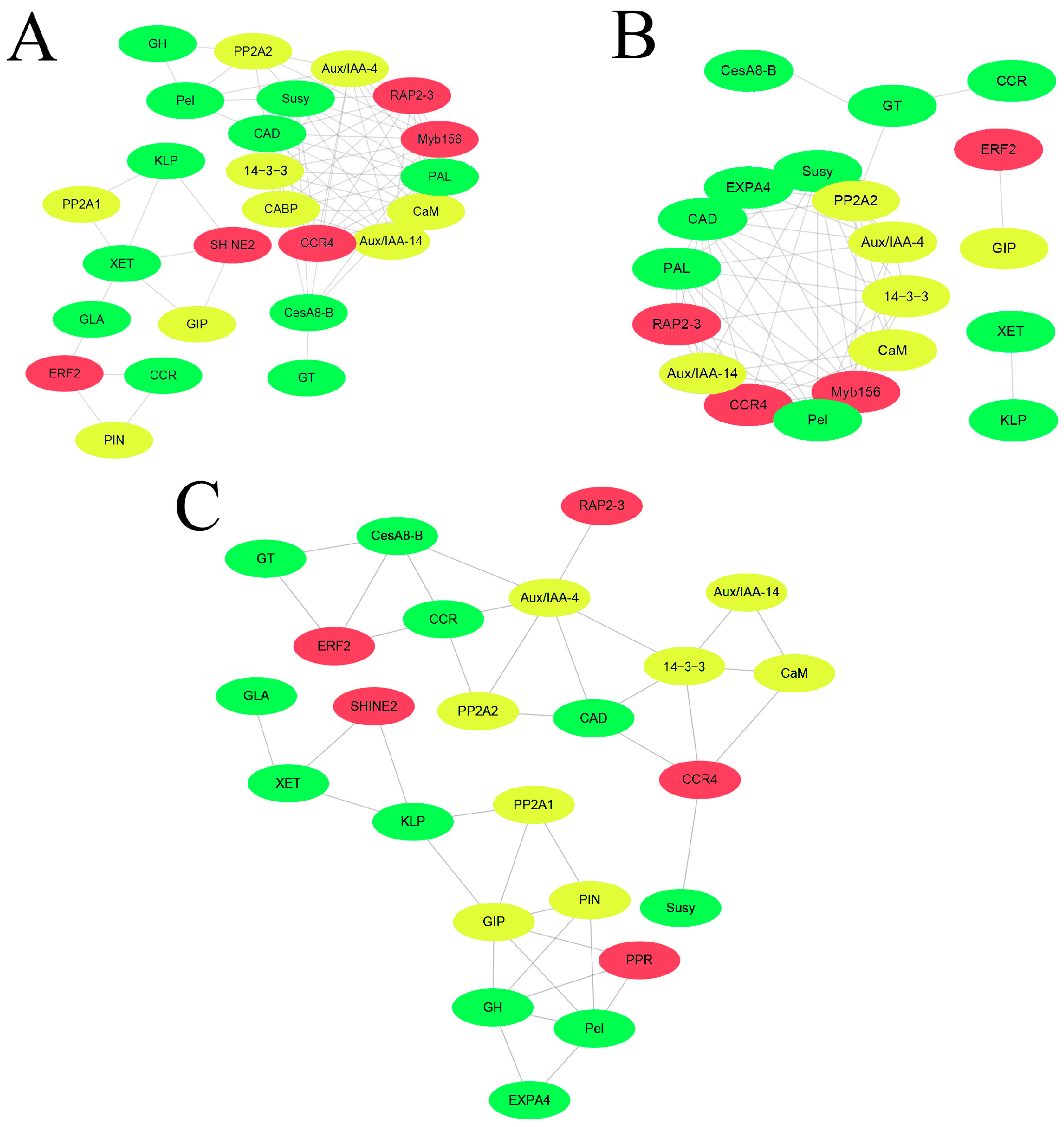
2.4. Correlation Network Analysis of Genes in Response to IAA, GA3, and BR Treatments
3. Discussion
4. Materials and Methods
4.1. Plant. Materials and Treatments
4.2. Anatomical Observations
4.3. RNA Extraction and Reverse Transcription (RT)
4.4. Quantitative Real-Time PCR
4.5. Correlation Network Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| BR | Brassinosteroid |
| CTAB | Cetyl trimethylammonium bromide |
| eBL | Epibrassinolide |
| GA | Gibberellin |
| GA3 | Gibberellic acid |
| IAA | Indole-3-acetic acid |
| Myb | Myeloblastosis |
| NAC | NAM–ATAF1,2–CUC2 |
| qPCR | quantitative real-time PCR |
| Susy | Sucrose synthase |
References
- Chen, J.; Chen, B.; Zhang, D. Transcript profiling of Populus tomentosa genes in normal, tension, and opposite wood by RNA-seq. BMC Genom. 2015, 16, 1641–1679. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, K.M.; Kauppinen, L.; Helariutta, Y. A weed for wood? Arabidopsis as a genetic model for xylem development. Plant Physiol. 2004, 135, 6536–6559. [Google Scholar] [CrossRef] [PubMed]
- Milhinhos, A.; Miguel, C.M. Hormone interactions in xylem development: A matter of signals. Plant Cell Rep. 2013, 32, 8678–8683. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, Y.; Liu, H.; Hu, P.; Jia, Y.; Zhang, C.; Wang, Y.; Gu, S.; Yang, C.; Wang, C. Exogenous GA3 Application Enhances Xylem Development and Induces the Expression of Secondary Wall Biosynthesis Related Genes in Betula platyphylla. Int. J. Mol. Sci. 2015, 16, 22960–22975. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, M.E.; Israelsson, M.; Olsson, O.; Moritz, T. Increased gibberellin biosynthesis in transgenic trees promotes growth, biomassproduction and xylem fiber length. Nat. Biotechnol. 2000, 18, 7847–7888. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, C.; Gómez Ros, L.V.; Pedreño, M.A.; Ros Barceló, A. Nitric oxide production by the differentiating xylem of Zinnia elegans. New Phytol. 2005, 165, 1211–1230. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Ye, Z.H. Secondary cell walls: Biosynthesis, patterned deposition and transcriptional regulation. Plant Cell Physiol. 2015, 56, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; McCarthy, R.L.; Lee, C.; Ye, Z.H. Dissection of the Transcriptional Program Regulating Secondary Wall Biosynthesis during Wood Formation in Poplar. Plant Physiol. 2011, 157, 1452–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björklund, S.; Antti, H.; Uddestrand, I.; Moritz, T.; Sundberg, B. Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 2007, 52, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, C.; Jin, X.; Xue, W.; Dubreuil, C.; Lezhneva, L.; Fischer, U. The plant hormone auxin directs timing of xylem development by inhibition of secondary cell wall deposition through repression of secondary wall NAC-domain transcription factors. Plant Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Little, C.H.A.; Pharis, R.P. Hormonal control of radial and longitudinal growth in the tree stem. In Plant Stems: Physiology and Functional Morphology; Gartner, B.L., Ed.; Academic Press: San Diego, CA, USA, 1995; pp. 281–319. [Google Scholar]
- Savidge, R.A. Xylogenesis, genetic and environmental regulation. IAWA J. 1996, 17, 269–310. [Google Scholar] [CrossRef]
- Little, C.H.A.; Savidge, R.A. The role of plant growth regulators in forest tree cambial growth. Plant Growth Regul. 1987, 6, 137–169. [Google Scholar] [CrossRef]
- Tuominen, H.; Puech, L.; Fink, S.; Sundberg, B. A radial concentration gradient of indole-3-acetic acid is Related to Secondary Xylem Development in Hybrid Aspen. Plant Physiol. 1997, 115, 5775–5785. [Google Scholar] [CrossRef]
- Uggla, C.; Mellerowicz, E.J.; Sundberg, B. Indole-3-acetic acid controls cambial growth in scots pine by positional signaling. Plant Physiol. 1998, 117, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Liu, L.H.; Zhao, N.; Zheng, S.X.; Xu, K.; Gu, S. Cooperation effects of GA3, IAA and uniconazole-P on tension wood formation and gravitropism in Fraxinus mandshurica Seedlings. J. Life Sci. 2009, 3, 1–7. [Google Scholar]
- Wareing, P.F. Interaction between indole-acetic acid and gibberellic acid in cambiaI activity. Nature 1958, 181, 17451–17746. [Google Scholar] [CrossRef]
- Funada, R.; Miura, T.; Shimizu, Y.; Kinase, T.; Nakaba, S.; Kubo, T.; Sano, Y. Gibberellin-induced formation of tension wood in angiosperm trees. Planta 2008, 227, 14091–14414. [Google Scholar] [CrossRef]
- Nugroho, W.D.; Yamagishi, Y.; Nakaba, S.; Fukuhara, S.; Begum, S.; Marsoem, S.N.; Ko, J.H.; Jin, H.O.; Funada, R. Gibberellin is required for the formation of tension wood and stem gravitropism in Acacia mangium seedlings. Ann. Bot. 2012, 110, 8878–8895. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Dilkes, B.P.; Gregory, B.D.; Ross, A.S.; Yuan, H.; Noguchi, T.; Fujioka, S.; Takatsuto, S.; Tanaka, A.; Yoshida, S.; et al. Arabidopsis dwarf1 is defective in the conversion of 24-methylenecholestrol to campesterol in brassinosteroid biosynthesis. Plant Physiol. 1999, 119, 8979–9007. [Google Scholar] [CrossRef]
- Yokota, T. The structure, biosynthesis and function of brassinosteroids. Trends Plant Sci. 1997, 2, 137–143. [Google Scholar] [CrossRef]
- Best, N.B.; Hartwig, T.; Budka, J.; Fujioka, S.; Johal, G.; Schulz, B.; Dilkes, B.P. nana plant2 Encodes a Maize Ortholog of the Arabidopsis Brassinosteroid Biosynthesis Gene DWARF1, Identifying Developmental Interactions between Brassinosteroids and Gibberellins. Plant physiol. 2016, 171, 26332–26647. [Google Scholar]
- Miyashima, S.; Sebastian, J.; Lee, J.Y.; Helariutta, Y. Stem cell function during plant vascular development. EMBO J. 2013, 32, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, J.; Karlberg, A.; Antti, H.; Lopez-Vernaza, M.; Mellerowicz, E.; Perrot-Rechenmann, C.; Sandberg, G.; Bhalerao, R.P.G. Dissecting the molecular basis of the regulation of wood formation by auxin in hybrid aspen. Plant Cell 2008, 20, 8438–8455. [Google Scholar] [CrossRef] [PubMed]
- Uggla, C.; Moritz, T.; Sandberg, G.; Sundberg, B. Auxin as a positional signal in pattern formation in plants. Proc. Natl. Acad. Sci. USA 1996, 93, 9282–9286. [Google Scholar] [CrossRef] [PubMed]
- Uggla, C.; Magel, E.; Moritz, T.; Sundberg, B. Function and dynamics of auxin and carbohydrates During earlywood/latewood transition in scots pine. Plant Physiol. 2001, 125, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Ridoutt, B.G.; Pharis, R.P. Metabolism of deuterium- and tritium-labeled gibberellins in cambial region tissues of Eucalyptus globulus stems. Tree Physiol. 1998, 18, 659–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israelsson, M.; Sundberg, B.; Moritz, T. Tissue-specific localization of gibberellins and expression of gibberellin-biosynthetic and signaling genes in wood-forming tissues in aspen. Plant J. 2005, 44, 494–504. [Google Scholar] [CrossRef]
- Nieminen, K.; Blomster, T.; Helariutta, Y.; Mähönen, A.P. Vascular Cambium Development. Arabidopsis Book 2015, 13, e0177. [Google Scholar] [CrossRef]
- Hossain, Z.; McGarvey, B.; Amyot, L.; Gruber, M.; Jung, J.; Hannoufa, A. DIMINUTO 1 affects the lignin profile and secondary cell wall formation in Arabidopsis. Planta 2012, 235, 4854–4898. [Google Scholar] [CrossRef]
- Li, W.F.; Yang, W.H.; Zhang, S.G.; Han, S.Y.; Qi, L.W. Transcriptome analysis provides insights into wood formation during larch tree aging. Tree Genet. Genomes 2017, 13, 19. [Google Scholar]
- Que, F.; Khadr, A.; Wang, G.L.; Li, T.; Wang, Y.H.; Xu, Z.S.; Xiong, A.S. Exogenous brassinosteroids altered cell length, gibberellin content, and cellulose deposition in promoting carrot petiole elongation. Plant Sci. 2018, 277, 1101–1120. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Do, J.; Shin, S.J.; Choi, J.W.; Choi, Y.I.; Kim, W.; Kwon, M. Exogenously applied 24-epi Brassinolide reduces lignification and alters cell wall carbohydrate biosynthesis in the secondary xylem of Liriodendron tulipifera. Phytochemistry 2014, 101, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.Q.; Yang, C.J.; Wang, X.L. Brassinosteroids can regulate cellulose biosynthesis by controlling the expression of CESA genes in Arabidopsis. J. Exp. Bot. 2016, 62, 44954–45506. [Google Scholar] [CrossRef]
- Reis, D.; Vian, B. Helicoidal pattern in secondary cell walls and possible role of xylans in their construction. C. R. Biol. 2004, 327, 7857–7890. [Google Scholar] [CrossRef]
- Mortimer, J.C.; Miles, G.P.; Brown, D.M.; Zhang, Z.; Segura, M.P.; Weimar, T.; Yu, X.; Seffen, K.A.; Stephens, E.; Turner, S.R.; et al. Absence of branches from xylan in Arabidopsis gux mutants reveals potential for simplification of lignocellulosic biomass. Proc. Natl. Acad. Sci. USA 2010, 107, 17409–17414. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.H.; Sun, J.; Oh, D.H.; Zielinski, R.E.; Clouse, S.D.; Huber, S.C. Enhancing Arabidopsis leaf growth by engineering the brassinosteroid insensitive receptor kinase. Plant Physiol. 2011, 157, 1201–1231. [Google Scholar] [CrossRef]
- Coleman, H.D.; Yan, J.; Mansfield, S.D. Sucrose synthase affects carbon partitioning to incresae cellulose production and altered cell wall ultrastructure. Proc. Natl. Acad. Sci. USA 2009, 106, 131181–133123. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zhou, R.; Louie, G.V.; Mühlemann, J.K.; Bomati, E.K.; Bowman, M.E.; Dudareva, N.; Dixon, R.A.; Noel, J.P.; Wang, X. Structural studies of cinnamoyl-CoA reductase and cinnamyl-alcohol dehydrogenase, key enzymes of monolignol biosynthesis. Plant Cell 2014, 26, 37093–37727. [Google Scholar] [CrossRef] [PubMed]
- Opassiri, R.; Pomthong, B.; Onkoksoong, T.; Akiyama, T.; Esen, A.; Ketudat Cairns, J.R. Analysis of rice glycosyl hydrolase family 1 and expression of Os4bglu12 β-glucosidase. BMC Plant Biol. 2006, 6, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Burk, D.H.; Morrison, W.H.; Ye, Z.H. A kinesin-like protein is essential for oriented deposition of cellulose microfibrils and cell wall strength. Plant Cell 2002, 14, 31013–31117. [Google Scholar] [CrossRef]
- Biswal, A.K.; Soeno, K.; Gandla, M.L.; Immerzeel, P.; Pattathil, S.; Lucenius, J.; Serimaa, R.; Hahn, M.G.; Moritz, T.; Jönsson, L.J.; Israelsson-Nordström, M.; Mellerowicz, E.J. Aspen pectate lyase PtxtPL1-27 mobilizes matrix polysaccharides from woody tissues and improves saccharification yield. Biotechnol. Biofuels 2014, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikubo, N.; Takahashi, J.; Roos, A.A.; Derba-Maceluch, M.; Piens, K.; Brumer, H.; Teeri, T.T.; Stålbrand, H.; Mellerowicz, E.J. Xyloglucan endo-transglycosylase-mediated xyloglucan rearrangements in developingwood of hybrid aspen. Plant Physiol. 2011, 155, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, E.; Porth, I.; Chen, J.G.; Mansfield, S.D.; Douglas, C.J. Regulation of secondary cell wall biosynthesis by poplar R2R3 MYB transcription factor PtrMYB152 in Arabidopsis. Sci. Rep. 2014, 4, 5054–5060. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Ye, Z.H. Evolutionary conservation of the transcriptional network regulating secondary cell wall biosynthesis. Trends Plant Sci. 2010, 15, 625–632. [Google Scholar] [CrossRef]
- McCarthy, R.L.; Zhong, R.; Fowler, S.; Lyskowski, D.; Piyasena, H.; Carleton, K.; Spicer, C.; Ye, Z.H. The poplar MYB transcription factors, PtrMYB3 and PtrMYB20, are involved in the regulation of secondary wall biosynthesis. Plant Cell Physiol. 2010, 51, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, M.; Hou, C.; Lu, T.; Liu, L.; Wei, H.; Cheng, Y.; Wei, Z. PsnSHN2 in Coordinated Regulation of Secondary Wall Components in Tobacco. Sci. Rep. 2017, 7, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Delannoy, E.; Stanley, W.A.; Bond, C.S.; Small, I.D. Pentatricopeptide repeat (PPR) proteins as sequence-specificity factors in post-transcriptional processes in organelles. Biochem. Soc. Trans. 2007, 35, 1643–1647. [Google Scholar] [CrossRef]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant. Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Thyssen, G.N.; Fang, D.D.; Zeng, L.; Song, X.; Delhom, C.D.; Condon, T.L.; Li, P.; Kim, H.J. The Immature Fiber Mutant Phenotype of Cotton (Gossypium hirsutum) Is Linked to a 22-bp Frame-Shift Deletion in a Mitochondria Targeted Pentatricopeptide Repeat Gene. G3 (Bethesda) 2016, 6, 16271–16633. [Google Scholar] [CrossRef]
- Nishimura, C.; Ohashi, Y.; Sato, S.; Kato, T.; Tabata, S.; Ueguchi, C. Histidine kinase homologs that act as cytokinin receptors possess overlapping functions in the regulation of shoot and root growth in Arabidopsis. Plant Cell 2004, 16, 1365–1377. [Google Scholar] [CrossRef]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Gene Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin synthesized by the YUCCA flavin monooxygenases is essential for embryogenesis and leaf formation in Arabidopsis. Plant Cell 2007, 19, 24302–24439. [Google Scholar] [CrossRef] [PubMed]
- Guilfoyle, T.; Hagen, G.; Ulmasov, T.; Murfett, J. How does auxin turn on genes? Plant Physiol. 1998, 118, 3413–3448. [Google Scholar] [CrossRef]
- Leyser, O. Molecular genetics of auxin signaling. Annu. Rev. Plant Biol. 2002, 53, 3773–3798. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Eckart, P.; Alsamadisi, N.; Noble, H.; Martin, C.; Spicer, R. Polar auxin transport is implicated in vessel differentiation and spatial patterning during secondary growth in Populus. Am. J. Bot. 2018, 105, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, H. Genome-wide identification and evolution of the PIN-FORMED (PIN) gene family in Glycine max. Genome 2017, 60, 5645–5671. [Google Scholar] [CrossRef]
- Shirakawa, M.; Ueda, H.; Shimada, T.; Nishiyama, C.; Hara-Nishimura, I. Vacuolar SNAREs function in the formation of the leaf vascular network by regulating auxin distribution. Plant Cell Physiol. 2009, 50, 13191–13328. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef]
- Li, L.; Xu, J.; Xu, Z.H.; Xue, H.W. Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis. Plant Cell 2005, 17, 27382–27753. [Google Scholar] [CrossRef]
- Karampelias, M.; Neyt, P.; De Groeve, S.; Aesaert, S.; Coussens, G.; Rolčík, J.; Bruno, L.; De Winne, N.; Van Minnebruggen, A.; Van Montagu, M.; et al. ROTUNDA3 function in plant development by phosphatase 2A-mediated regulation of auxin transporter recycling. Proc. Natl. Acad. Sci. USA 2016, 113, 27682–27773. [Google Scholar] [CrossRef]
- Lottersberger, F.; Panza, A.; Lucchini, G.; Piatti, S.; Longhese, M.P. The Saccharomyces cerevisiae 14-3-3 Proteins Are Required for the G1/S Transition, Actin Cytoskeleton Organization and Cell Wall Integrity. Genetics 2006, 173, 6616–6675. [Google Scholar] [CrossRef] [PubMed]
- Jaspert, N.; Weckermann, K.; Möller, C.; Throm, C.; Kintzi, A.; Oecking, C. Arabidopsis 14-3-3 epsilon members contribute to polarity of PIN auxin carrier and auxin transport-related development. Elife 2017, 6, E24336–E24356. [Google Scholar]
- Chen, J.; Mao, L.; Mi, H.; Lu, W.; Ying, T.; Luo, Z. Involvement of three annexin genes in the ripening of strawberry fruit regulated by phytohormone and calcium signal transduction. Plant Cell Rep. 2016, 35, 7337–7343. [Google Scholar] [CrossRef] [PubMed]
- Blakeslee, J.J.; Peer, W.A.; Murphy, A.S. Auxin transport. Curr. Opin. Plant Biol. 2005, l8, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Ibañes, M.; Fàbregas, N.; Chory, J.; Caño-Delgado, A.I. Brassinosteroid signaling and auxin transport are required to establish the periodic pattern of Arabidopsis shoot vascular bundles. Proc. Natl. Acad. Sci. USA 2009, 106, 13630–13635l. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D.; Dai, X.H.; Blackwell, H.E.; Schreiber, S.L.; Chory, J. SIR1, an upstream component in auxin signaling identified by chemical genetics. Science 2003, 301, 11071–11110. [Google Scholar] [CrossRef]
- Kim, B.; Kwon, M.; Jeon, J.; Schulz, B.; Corvalán, C.; Jeong, Y.J.; Choe, S. The Arabidopsis gulliver2/phyB mutant exhibits reduced sensitivity to brassinazole. J. Plant Biol. 2014, 57, 202–207. [Google Scholar] [CrossRef]
- Zubo, Y.O.; Yamburenko, M.V.; Kusnetsov, V.V.; Börner, T. Methyl jasmonate, gibberellic acid, and auxin affect transcription and transcript accumulation of chloroplast genes in barley. J. Plant Physiol. 2011, 168, 13354–13413. [Google Scholar] [CrossRef]
- Schrader, J.; Baba, K.; May, S.T.; Palme, K.; Bennett, M.; Bhalerao, R.P.; Sandberg, G. Polar auxin transport in the wood-forming tissues of hybrid aspen is under simultaneous control of developmental and environmental signals. Proc. Natl. Acad. Sci. USA 2003, 100, 10096–100101. [Google Scholar] [CrossRef]
- Shimada, Y.; Goda, H.; Nakamura, A.; Takatsuto, S.; Fujioka, S.; Yoshida, S. Organ-specific expression of brassinosteroid-biosynthetic genes and distribution of endogenous brassinosteroids in Arabidopsis. Plant Physiol. 2003, 131, 287–297. [Google Scholar] [CrossRef]
- Gregory, M.S.; James, R. Brassinosteroids Do Not Undergo Long-Distance Transport in Pea. Implications for the Regulation of Endogenous Brassinosteroid Levels. Plant Physiol. 2004, 135, 2196–2206. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Accession NO | Description | Primer sequences (forward) | Primer sequences (reverse) |
|---|---|---|---|---|
| Cell wall biosynthesis | ||||
| CAD | Potri.009G095800 | Cinnamyl-alcohol dehydrogenase | GCATCTCGCTCCTTACACCT | TCCCACCTCAACAACCTCAC |
| CCR | Potri.009G076300 | Cinnamoyl-CoA reductase | ATACCGTTCACGCCACCG | GAAAGACGCCAGCACAGC |
| CesA8-B | Potri.004G059600 | Cellulose synthase 8-B | GGCTTGCCATGAGTGTAA | ATCCTGAGAATCGTTGAG |
| EXPA4 | Potri.010G202500 | Alpha-expansin 4 precursor | AATCCTCCTCGTCCTCACTTC | CCTTCACGCTCACTTTCACAA |
| GH | Potri.008G120000 | Glycosyl hydrolases family 31 | TGGGAAGTGCCATACAATCT | TCACCAAATGACCCTGAACC |
| GLA | Potri.007G099800 | Beta-galactosidase | CTGCTGCCATTCATTATCCT | TGTTTCAATCACATCCACCC |
| GT | Potri.002G200200 | Glycosyl transferase family 8 | AAGCCATTCACTCCTCCAA | GAGAAGCCGCATTCATCAG |
| KLP | Potri.010G153000 | Kinesin-like protein | GCAGCACAATCAGAGCCTAAC | ATAACCTCCCAATGCACCACC |
| PAL | Potri.008G038200 | Phenylalanine ammonialyase | AACCCAACTATTCCAAAC | CTTCAAGCATTCCAGCAT |
| Pel | Potri.015G087800 | Pectate lyase | GGAGAAGAACCGTCAGAGGC | ACGGGATCATCATTACCAGAGT |
| Susy | Potri.006G136700 | Sucrose synthase | AACTTCGTGCTTGAATTGGACT | AACAACTTAGCTGAAAGGTGGC |
| XET | Potri.005G201200 | Xyloglucan Endotransglycosylase | GATTCTGGTTCTGGCTTCC | GTCAATCTCGTCGTGGGTC |
| Transcription factors | ||||
| CCR4 | Potri.002G182500 | Transcriptional effector CCR4 | CTGCTAACTGCTGTGCGTAA | TTGGCATAAGGTTGAGTTTGTT |
| ERF2 | Potri.001G154100 | Ethylene-responsive transcription factor 2 | CTTACGACCGTGCTGCCTAT | TGCCACCAACCTTCTTCCT |
| Myb156 | Potri.009G134000 | Myb-like DNA-binding domain | GAAGATTACCAGGGAGAACAGAT | CTGAAGTAGTAGTCGTGGTGAAAG |
| PPR | Potri.011G120900 | Pentatricopeptide Repeat Protein | GGGTTGAAGAGGGCAAGAAA | AATCAGATCCAGCCGCACAG |
| RAP2-3 | Potri.008G210900 | Ethylene-responsive transcription factor | CGGAGATTGACAGTTGAGGAT | ACTTGCTGGACTTGGATGGTG |
| SHINE2 | Potri.018G028000 | Ethylene-responsive transcription factor | GACCCTTCTCCTTCACTCACT | CCTCTTCTTCCGTACCATTTT |
| Signal transduction | ||||
| Aux/IAA 4 | Potri.013G041300 | Auxin-responsive protein 4 | GCCTGACATGAATGAAGAGCC | CTTGATGGGTGGAGCAGTTTC |
| Aux/IAA 14 | Potri.008G161200 | Auxin-responsive protein 14 | ATGGAGCCCAGGGAATGATAG | AGGATGGCACATACTCGGAAC |
| CABP | Potri.016G024700 | Calcium binding protein | ATAAGGATGGCGATGGTTGC | AGCCTCAGTCGGGTTCTGTC |
| CaM | Potri.012G041000 | Calmodulin-like protein 6a | GAATGGCTTTATTTCTGCTGC | CATCCTCACAAACTCCTCGTA |
| GIP | Potri.017G083000 | Gibberellin regulated protein | GCTGTCTTCCTCTTGGCTCT | GGCTTATGGTACTGGGTCTTG |
| PIN | Potri.015G038700 | Auxin efflux carrier component 1 | ACCATTACATTGTTCTCGCTTTC | GAGATGAGCAGTTTAGCACCC |
| PP2A1 | Potri.010G054300 | Serine/threonine protein phosphatase 2A | CTGGTCTTGATTCTGCTCCTC | GCAATGCTTCATACGGTGG |
| PP2A2 | Potri.003G015400 | Serine/threonine protein phosphatase 2A | CTTCGCCATCCCAACATAAT | CTCGTCTTCGCTGAATCGTC |
| 14-3-3 | Potri.T147900 | 14-3-3 protein | GGCTCCTACCCACCCAATA | GCAAGACTGCAAGCACGAT |
| Reference genes | ||||
| TUA | Potri.003G220300 | Tubulin alpha-3/alpha-5 chain | AGGTTCTGGTTTGGGGTCTT | TTGTCCAAAAGCACAGCAAC |
| UBQ | Potri.001G418500 | Polyubiquitin | GTTGATTTTTGCTGGGAAGC | GATCTTGGCCTTCACGTTGT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, H.; Zhao, L.; Guo, W.; Yu, Y.; Tao, L.; Zhang, L.; Song, X.; Huang, W.; Cheng, L.; Chen, J.; et al. Exogenous Application of Phytohormones Promotes Growth and Regulates Expression of Wood Formation-Related Genes in Populus simonii × P. nigra. Int. J. Mol. Sci. 2019, 20, 792. https://doi.org/10.3390/ijms20030792
Yuan H, Zhao L, Guo W, Yu Y, Tao L, Zhang L, Song X, Huang W, Cheng L, Chen J, et al. Exogenous Application of Phytohormones Promotes Growth and Regulates Expression of Wood Formation-Related Genes in Populus simonii × P. nigra. International Journal of Molecular Sciences. 2019; 20(3):792. https://doi.org/10.3390/ijms20030792
Chicago/Turabian StyleYuan, Hongmei, Lijuan Zhao, Wendong Guo, Ying Yu, Lei Tao, Liguo Zhang, Xixia Song, Wengong Huang, Lili Cheng, Jing Chen, and et al. 2019. "Exogenous Application of Phytohormones Promotes Growth and Regulates Expression of Wood Formation-Related Genes in Populus simonii × P. nigra" International Journal of Molecular Sciences 20, no. 3: 792. https://doi.org/10.3390/ijms20030792
APA StyleYuan, H., Zhao, L., Guo, W., Yu, Y., Tao, L., Zhang, L., Song, X., Huang, W., Cheng, L., Chen, J., Guan, F., Wu, G., & Li, H. (2019). Exogenous Application of Phytohormones Promotes Growth and Regulates Expression of Wood Formation-Related Genes in Populus simonii × P. nigra. International Journal of Molecular Sciences, 20(3), 792. https://doi.org/10.3390/ijms20030792