GmZPR3d Interacts with GmHD-ZIP III Proteins and Regulates Soybean Root and Nodule Vascular Development

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Identification of Soybean HD-ZIP III and ZPR Genes in Soybean
2.2. GmZPR3d Displayed Strong Interaction Capacity with GmHD-ZIP III Proteins
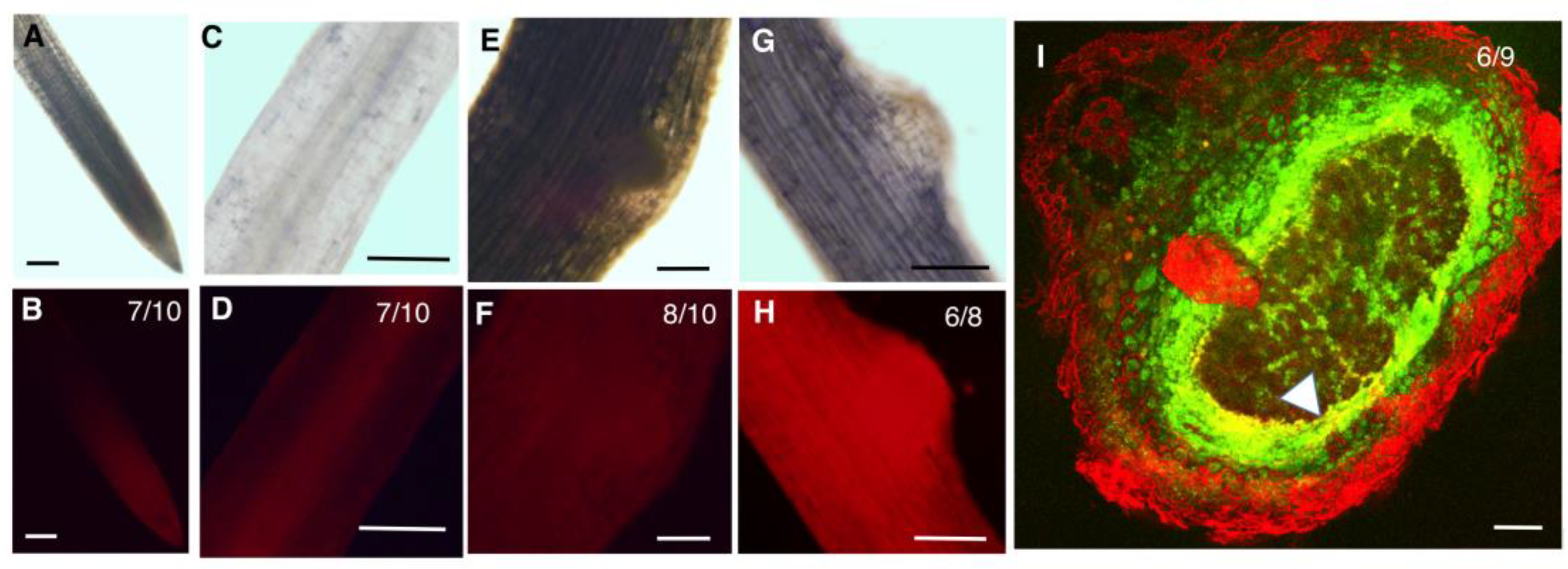
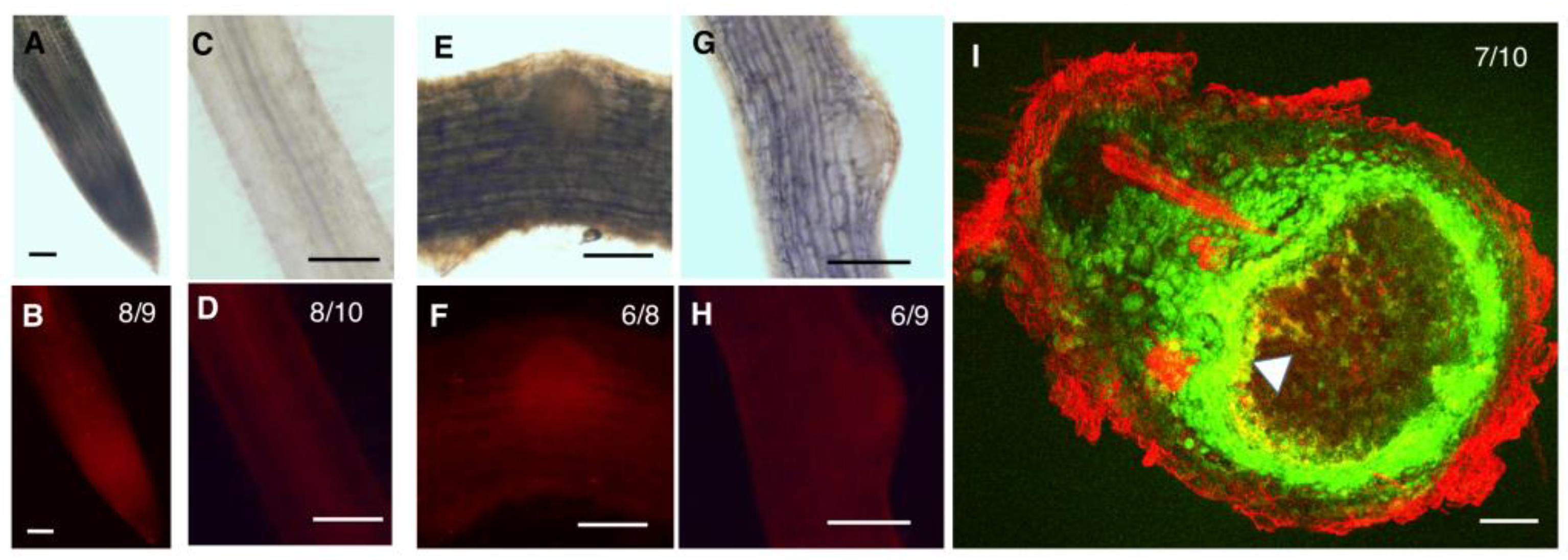
2.3. GmZPR3d and GmHD-ZIP III-1 and -2 Display Overlapping Expression Patterns in the Root and Nodule Vasculature
2.4. Over-Expression of GmZPR3d Alters Root Vascular Development
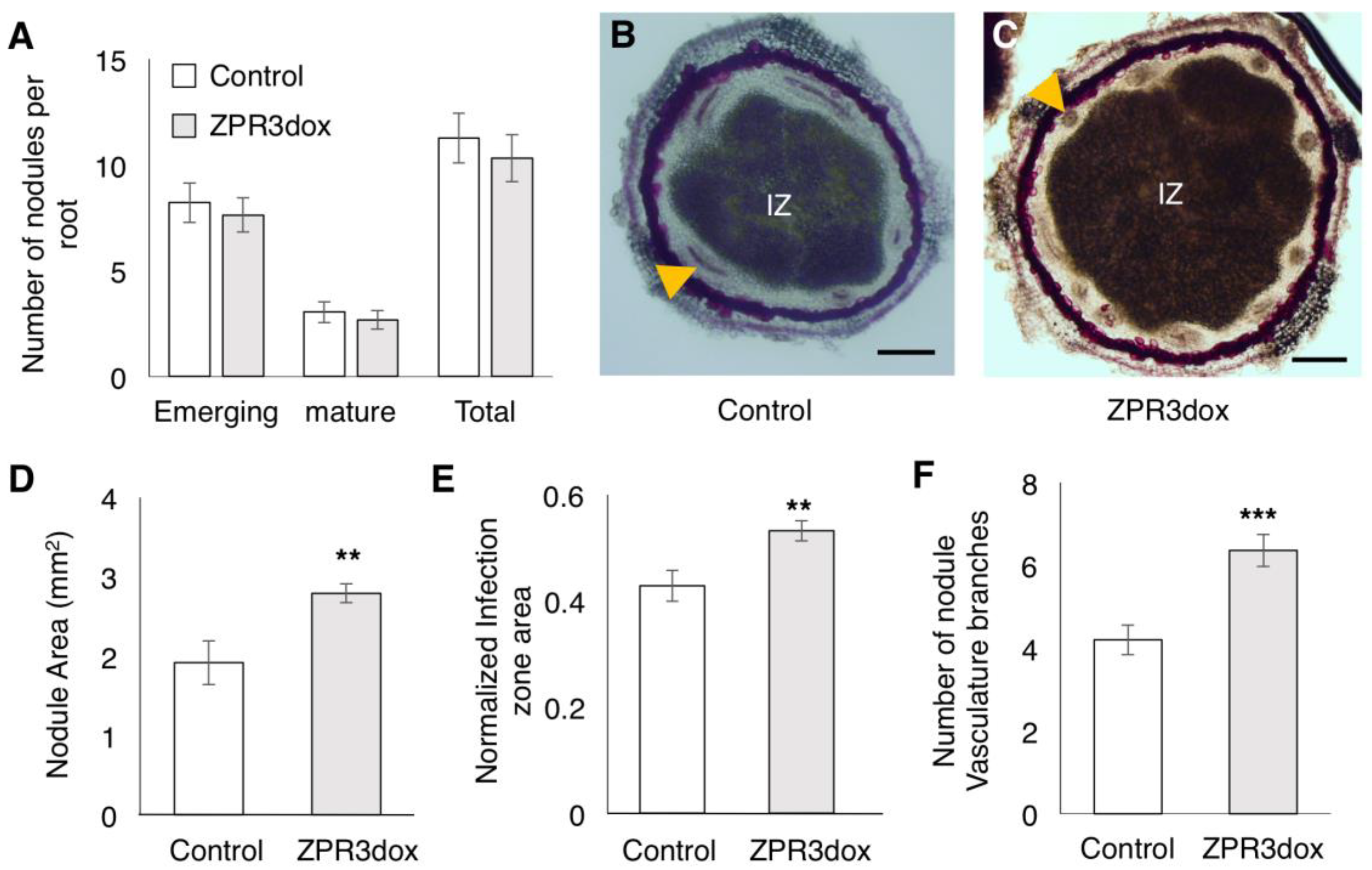
2.5. Over-Expression of GmZPR3d Alters Nodule Size and Vascular Bundle Branching
2.6. ZPR3d Interacts with A Number of Soybean Proteins
3. Discussion
4. Materials and Methods
4.1. HD-ZIP III and ZPR Phylogenetic Analysis
4.2. Cloning and Construction of DNA Vectors
4.3. One-on-One Yeast Two-Hybrid Assays
4.4. Plant Materials and Composite Plant Generation
4.5. Plant Transformation and Nodulation Assay
4.6. Staining, Microscopy and Image Analysis
4.7. Gene Expression Analysis by RT-qPCR
4.8. Genome-Wide Yeast Two-Hybrid Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ZPR | Small Leucine Zipper |
| HD-ZIP III | Class III Homeodomain Leucine Zipper |
| BLAST | Basic Local Alignment Search Tool |
| SD | Synthetic Dropout |
| Leu | Leucine |
| Trp | Tryptophan |
| QDO | Quadruple Drop Out |
| AbA | Aureobasidin |
| GFP | Green Fluorescent Protein |
References
- Hawker, N.P.; Bowman, J.L. Roles for Class III HD-ZIP and KANADI Genes in arabidopsis Root Development. Plant Physiol. 2004, 135, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Izhaki, A.; Bowman, J.L. KANADI and Class III HD-ZIP Gene Families Regulate Embryo Patterning and Modulate Auxin Flow during Embryogenesis in arabidopsis. Plant Cell 2007, 19, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial Patterning of Arabidopsis Shoots by Class III HD-ZIP and KANADI Genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-ZIP) proteins. Plant Signal. Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-ZIP family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Magnani, E.; Barton, M.K. A Per-ARNT-Sim-Like Sensor Domain Uniquely Regulates the Activity of the Homeodomain Leucine Zipper Transcription Factor REVOLUTA in arabidopsis. Plant Cell Online 2011, 23, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III Homeodomain-Leucine Zipper Gene Family Members Have Overlapping, Antagonistic, and Distinct Roles in arabidopsis Development. Plant Cell Online 2005, 17, 61–76. [Google Scholar] [CrossRef]
- Baima, S.; Possenti, M.; Matteucci, A.; Wisman, E.; Altamura, M.M.; Ruberti, I.; Morelli, G. The arabidopsis ATHB-8 HD-Zip Protein Acts as a Differentiation-Promoting Transcription Factor of the Vascular Meristems. Plant Physiol. 2001, 126, 643–655. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Indrasumunar, A.; Hayashi, S.; Lin, M.H.; Lin, Y.H.; Reid, D.E.; Gresshoff, P.M. Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant Biol. 2010, 52, 61–76. [Google Scholar] [CrossRef]
- Hirsch, A.M. Developmental biology of legume nodulation. New Phytol. 1992, 122, 211–237. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Walsh, K. Anatomy of the Legume Nodule Cortex With Respect to Nodule Permeability. Funct. Plant Biol. 1994, 21, 49–68. [Google Scholar] [CrossRef]
- Walsh, K.B.; Atkins, R.S.; Low, C.S. Vascular anatomy of fabaceous nodules of determinate growth. Plant Cell Environ. 1992, 15, 849–854. [Google Scholar] [CrossRef]
- Frugier, F.; Kosuta, S.; Murray, J.D.; Crespi, M.; Szczyglowski, K. Cytokinin: secret agent of symbiosis. Trends Plant Sci. 2008, 13, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Suzaki, T.; Ito, M.; Kawaguchi, M. Genetic basis of cytokinin and auxin functions during root nodule development. Front. Plant Sci. 2013, 4, 42. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, A.B.; Lombardo, F.; Miwa, H.; Perry, J.A.; Bunnewell, S.; Parniske, M.; Wang, T.L.; Downie, J.A. Lotus japonicus nodulation requires two GRAS domain regulators, one of which is functionally conserved in a non-legume. Plant Physiol. 2006, 142, 1739–1750. [Google Scholar] [CrossRef] [PubMed]
- Nontachaiyapoom, S.; Scott, P.T.; Men, A.E.; Kinkema, M.; Schenk, P.M.; Gresshoff, P.M. Promoters of orthologous Glycine max and Lotus japonicus nodulation autoregulation genes interchangeably drive phloem-specific expression in transgenic plants. Mol. Plant Microbe Interact. 2007, 20, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Turner, M.; Yu, O.; Subramanian, S. miR393 and miR164 influence indeterminate but not determinate nodule development. Plant Signal. Behav. 2013, 9, e26753. [Google Scholar] [CrossRef]
- Boualem, A.; Laporte, P.; Jovanovic, M.; Laffont, C.; Plet, J.; Combier, J.P.; Niebel, A.; Crespi, M.; Frugier, F. MicroRNA166 controls root and nodule development in Medicago truncatula. Plant J. 2008, 54, 876–887. [Google Scholar] [CrossRef] [Green Version]
- Wenkel, S.; Emery, J.; Hou, B.-H.; Evans, M.M.S.; Barton, M.K. A Feedback Regulatory Module Formed by LITTLE ZIPPER and HD-ZIPIII Genes. Plant Cell Online 2007, 19, 3379–3390. [Google Scholar] [CrossRef] [Green Version]
- Brandt, R.; Salla-Martret, M.; Bou-Torrent, J.; Musielak, T.; Stahl, M.; Lanz, C.; Ott, F.; Schmid, M.; Greb, T.; Schwarz, M.; et al. Genome-wide binding-site analysis of REVOLUTA reveals a link between leaf patterning and light-mediated growth responses. Plant J. 2012, 72, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Deppmann, C.D.; Acharya, A.; Rishi, V.; Wobbes, B.; Smeekens, S.; Taparowsky, E.J.; Vinson, C. Dimerization specificity of all 67 B-ZIP motifs in Arabidopsis thaliana: A comparison to Homo sapiens B-ZIP motifs. Nucleic Acids Res. 2004, 32, 3435–3445. [Google Scholar] [CrossRef] [PubMed]
- Landschulz, W.H.; Johnson, P.F.; McKnight, S.L. The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins. Science 1988, 240, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, S.; Subramanian, S. South Dakota State University, Brookings, SD. Unpublished observations from yeast-2-hybrid assays between GmZPR and GmHD-ZIP III proteins. 2016. [Google Scholar]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Turner, M.; Nizampatnam, N.R.; Baron, M.; Coppin, S.; Damodaran, S.; Adhikari, S.; Arunachalam, S.P.; Yu, O.; Subramanian, S. Ectopic expression of miR160 results in auxin hypersensitivity, cytokinin hyposensitivity, and inhibition of symbiotic nodule development in soybean. Plant Physiol. 2013, 162, 2042–2055. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Carlsbecker, A.; Etchells, J.P. Class III HD-ZIPs govern vascular cell fate: an HD view on patterning and differentiation. J. Exp. Bot. 2017, 68, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Avci, U.; Li, S.; Gallego-Giraldo, L.; Pattathil, S.; Qi, L.; Hahn, M.G.; Wang, H. Activation of miR165b represses AtHB15 expression and induces pith secondary wall development in Arabidopsis. Plant J. 2015, 83, 388–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Jung, J.-H.; Reyes, J.L.; Kim, Y.-S.; Kim, S.-Y.; Chung, K.-S.; Kim, J.A.; Lee, M.; Lee, Y.; Kim, V.N.; et al. microRNA-directed cleavage of ATHB15 mRNA regulates vascular development in Arabidopsis inflorescence stems. Plant J. 2005, 42, 84–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, M.; Udagawa, M.; Nishikubo, N.; Horiguchi, G.; Yamaguchi, M.; Ito, J.; Mimura, T.; Fukuda, H.; Demura, T. Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 2005, 19, 1855–1860. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Goue, N.; Igarashi, H.; Ohtani, M.; Nakano, Y.; Mortimer, J.C.; Nishikubo, N.; Kubo, M.; Katayama, Y.; Kakegawa, K.; et al. VASCULAR-RELATED NAC-DOMAIN6 and VASCULAR-RELATED NAC-DOMAIN7 effectively induce transdifferentiation into xylem vessel elements under control of an induction system. Plant Physiol. 2010, 153, 906–914. [Google Scholar] [CrossRef]
- Du, Q.; Wang, H. The role of HD-ZIP III transcription factors and miR165/166 in vascular development and secondary cell wall formation. Plant Signal. Behav. 2015, 10, e1078955. [Google Scholar] [CrossRef] [Green Version]
- Staudt, A.-C.; Wenkel, S. Regulation of protein function by “microProteins”. EMBO Rep. 2011, 12, 35–42. [Google Scholar] [CrossRef]
- Husbands, A.Y.; Aggarwal, V.; Ha, T.; Timmermans, M.C.P. In Planta Single-Molecule Pull-Down Reveals Tetrameric Stoichiometry of HD-ZIPIII:LITTLE ZIPPER Complexes. Plant Cell 2016, 28, 1783–1794. [Google Scholar] [CrossRef] [Green Version]
- Xulvi-Brunet, R.; Li, H. Co-expression networks: Graph properties and topological comparisons. Bioinformatics 2010, 26, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Farmer, A.; Joshi, T.; Takahashi, K.; Langley, R.J.; Franklin, L.D.; He, J.; Xu, D.; May, G.; Stacey, G. An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J. 2010, 63, 86–99. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Kim, S.-G.; Lee, M.; Lee, I.; Park, H.-Y.; Seo, P.J.; Jung, J.-H.; Kwon, E.-J.; Suh, S.W.; Paek, K.-H.; et al. HD-ZIP III Activity Is Modulated by Competitive Inhibitors via a Feedback Loop in Arabidopsis Shoot Apical Meristem Development. Plant Cell Online 2008, 20, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, S.; Westfall, C.; Kisely, B.; Jez, J.; Subramanian, S. Nodule-Enriched GRETCHEN HAGEN 3 Enzymes Have Distinct Substrate Specificities and Are Important for Proper Soybean Nodule Development. Int. J. Mol. Sci. 2017, 18, 2547. [Google Scholar] [CrossRef] [PubMed]
- Van Nocker, S.; Ludwig, P. The WD-repeat protein superfamily in Arabidopsis: conservation and divergence in structure and function. BMC Genomics 2003, 4, 50. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yin, S.; Liu, X.; Liu, B.; Yang, S.; Xue, S.; Cai, Y.; Black, K.; Liu, H.; Dong, M.; et al. The WD-Repeat Protein CsTTG1 Regulates Fruit Wart Formation through Interaction with the Homeodomain-Leucine Zipper I Protein Mict. Plant Physiol. 2016, 171, 1156–1168. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Nizampatnam, N.R.; Schreier, S.J.; Damodaran, S.; Adhikari, S.; Subramanian, S. microRNA160 dictates stage-specific auxin and cytokinin sensitivities and directs soybean nodule development. Plant J. 2015, 84, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Collier, R.; Fuchs, B.; Walter, N.; Kevin Lutke, W.; Taylor, C.G. Ex vitro composite plants: an inexpensive, rapid method for root biology. Plant J. 2005, 43, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Bhuvaneswari, T.V.; Turgeon, B.G.; Bauer, W.D. Early Events in the Infection of Soybean (Glycine max L. Merr) by Rhizobium japonicum: I. LOCALIZATION OF INFECTIBLE ROOT CELLS. Plant Physiol. 1980, 66, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Thibivilliers, S.; Bilgin, D.D.; Radwan, O.; Benitez, M.; Clough, S.J.; Stacey, G. Identification of Four Soybean Reference Genes for Gene Expression Normalization. Plant Genome 2008, 1, 44–54. [Google Scholar] [CrossRef]
- Formstecher, E.; Aresta, S.; Collura, V.; Hamburger, A.; Meil, A.; Trehin, A.; Reverdy, C.; Betin, V.; Maire, S.; Brun, C.; et al. Protein interaction mapping: A Drosophila case study. Genome Res. 2005, 15, 376–384. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damodaran, S.; Dubois, A.; Xie, J.; Ma, Q.; Hindié, V.; Subramanian, S. GmZPR3d Interacts with GmHD-ZIP III Proteins and Regulates Soybean Root and Nodule Vascular Development. Int. J. Mol. Sci. 2019, 20, 827. https://doi.org/10.3390/ijms20040827
Damodaran S, Dubois A, Xie J, Ma Q, Hindié V, Subramanian S. GmZPR3d Interacts with GmHD-ZIP III Proteins and Regulates Soybean Root and Nodule Vascular Development. International Journal of Molecular Sciences. 2019; 20(4):827. https://doi.org/10.3390/ijms20040827
Chicago/Turabian StyleDamodaran, Suresh, Amélie Dubois, Juan Xie, Qin Ma, Valérie Hindié, and Senthil Subramanian. 2019. "GmZPR3d Interacts with GmHD-ZIP III Proteins and Regulates Soybean Root and Nodule Vascular Development" International Journal of Molecular Sciences 20, no. 4: 827. https://doi.org/10.3390/ijms20040827
APA StyleDamodaran, S., Dubois, A., Xie, J., Ma, Q., Hindié, V., & Subramanian, S. (2019). GmZPR3d Interacts with GmHD-ZIP III Proteins and Regulates Soybean Root and Nodule Vascular Development. International Journal of Molecular Sciences, 20(4), 827. https://doi.org/10.3390/ijms20040827