Integrated Multi-Assay Culture Model for Stem Cell Chondrogenic Differentiation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
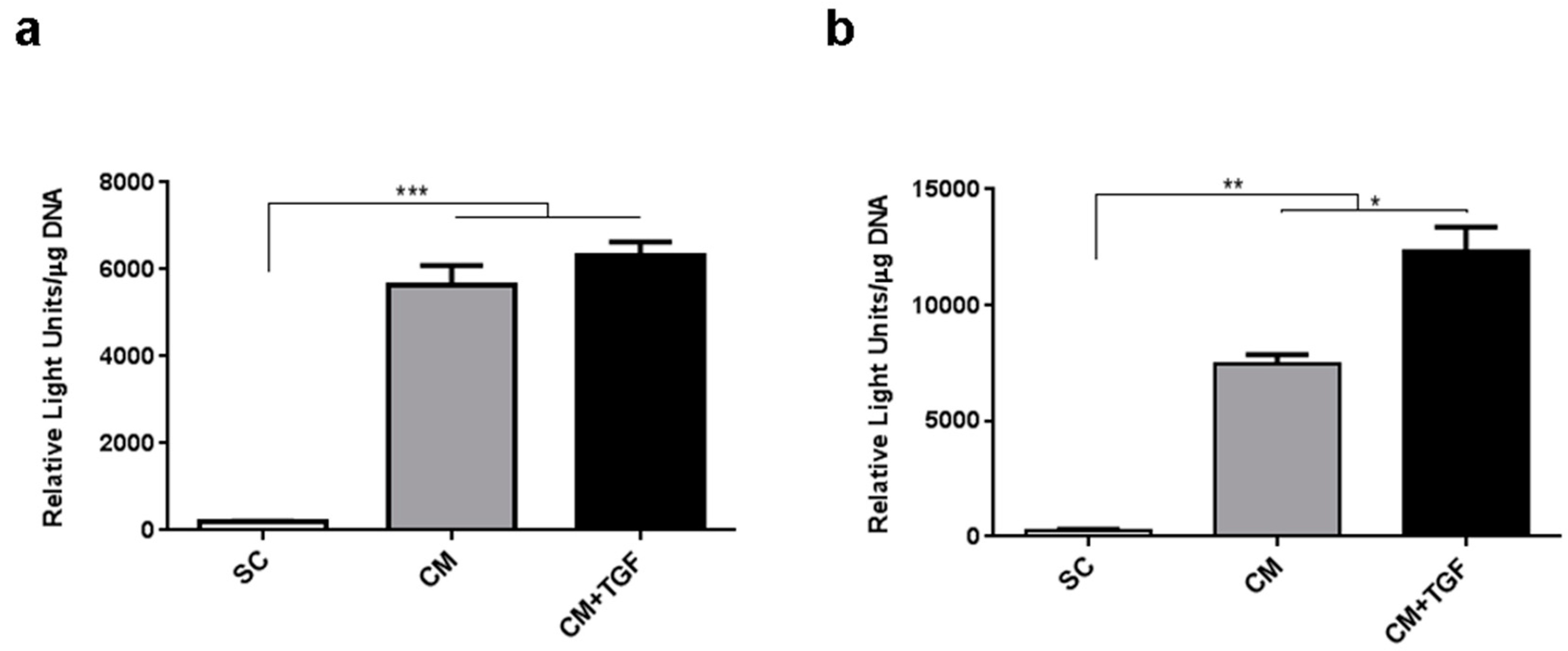
2.1. Optimising In Vitro Conditions for MSC Chondrogenesis
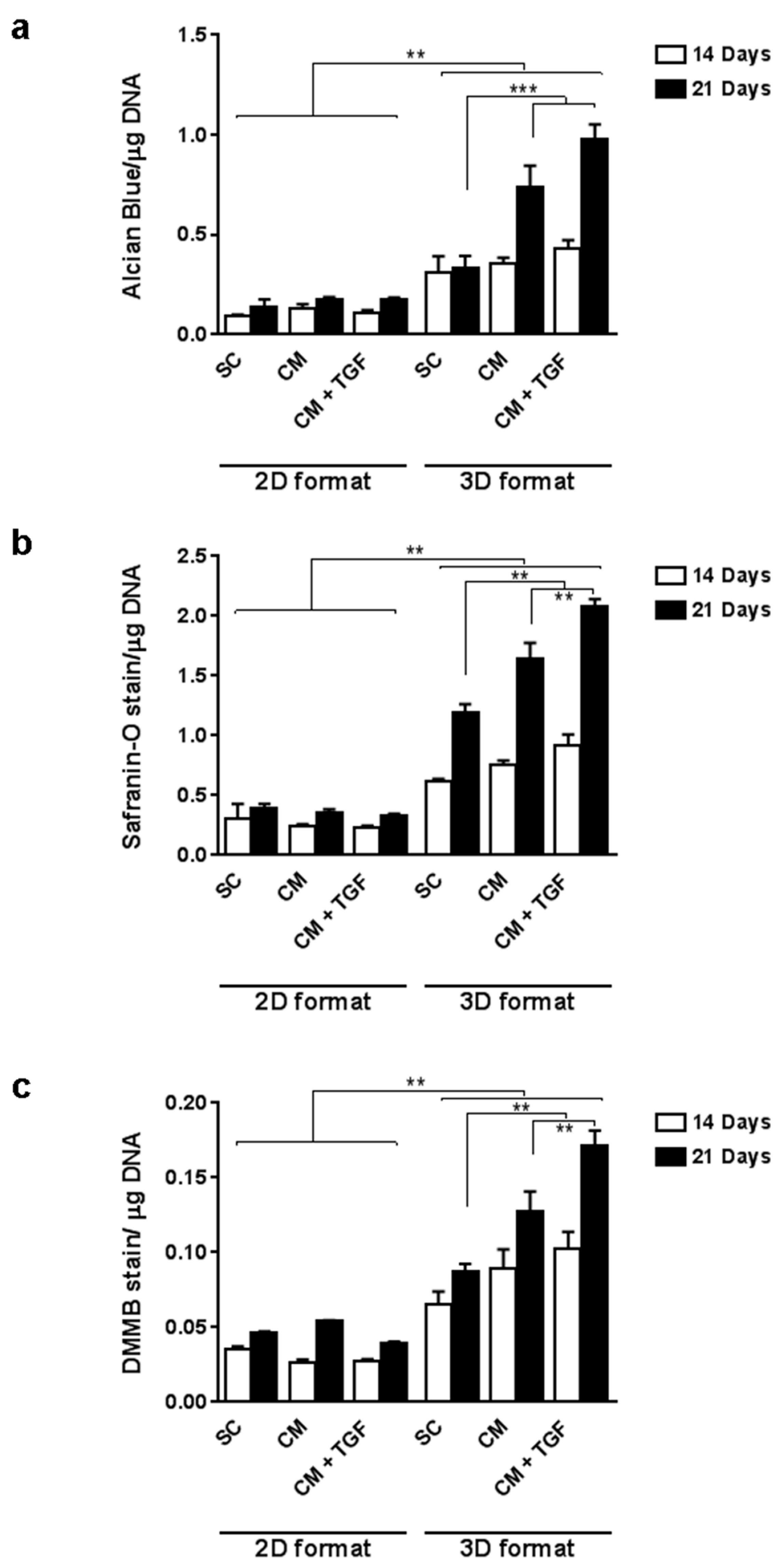
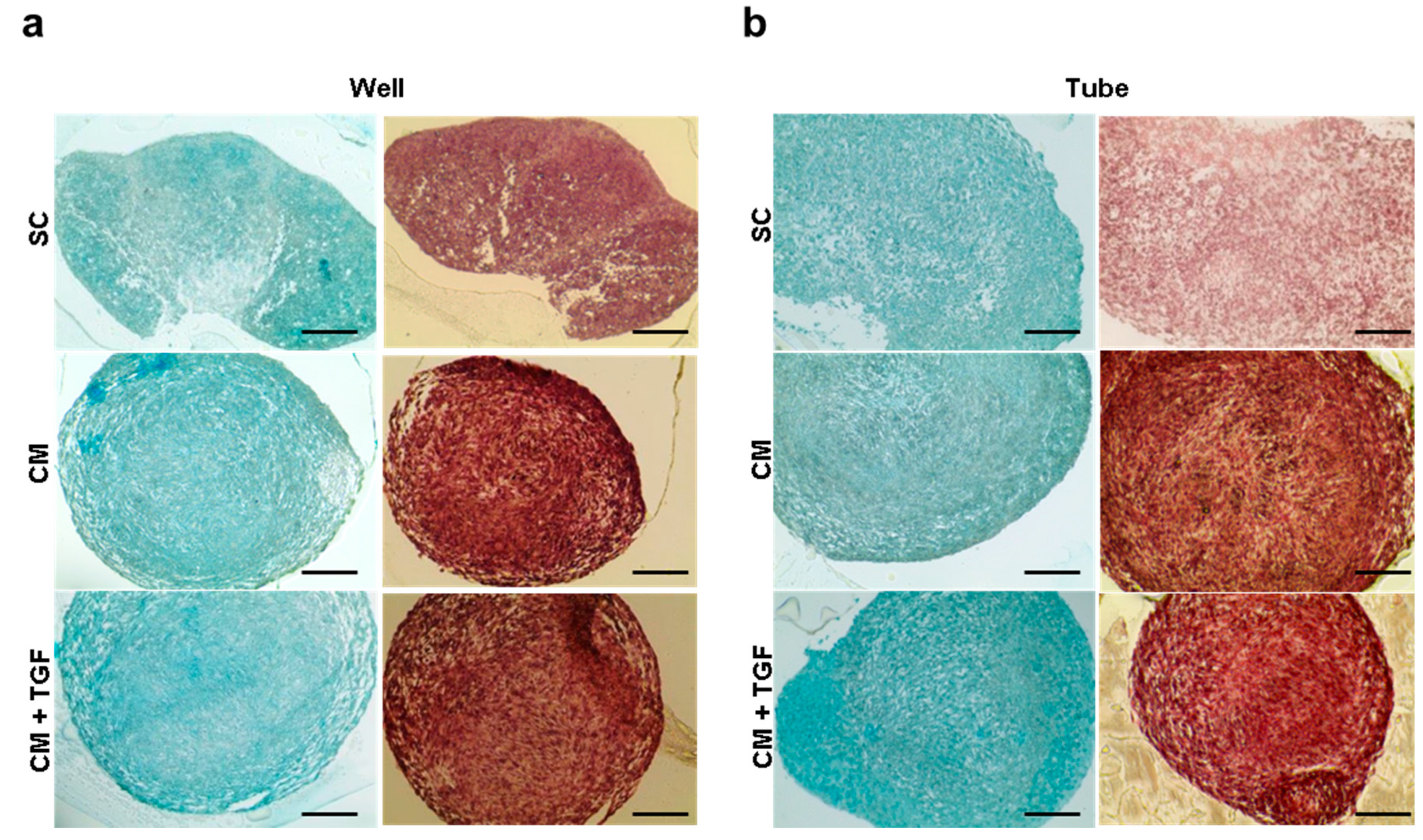
2.2. Comparing 2D and 3D Chondrogenic Differentiation Models
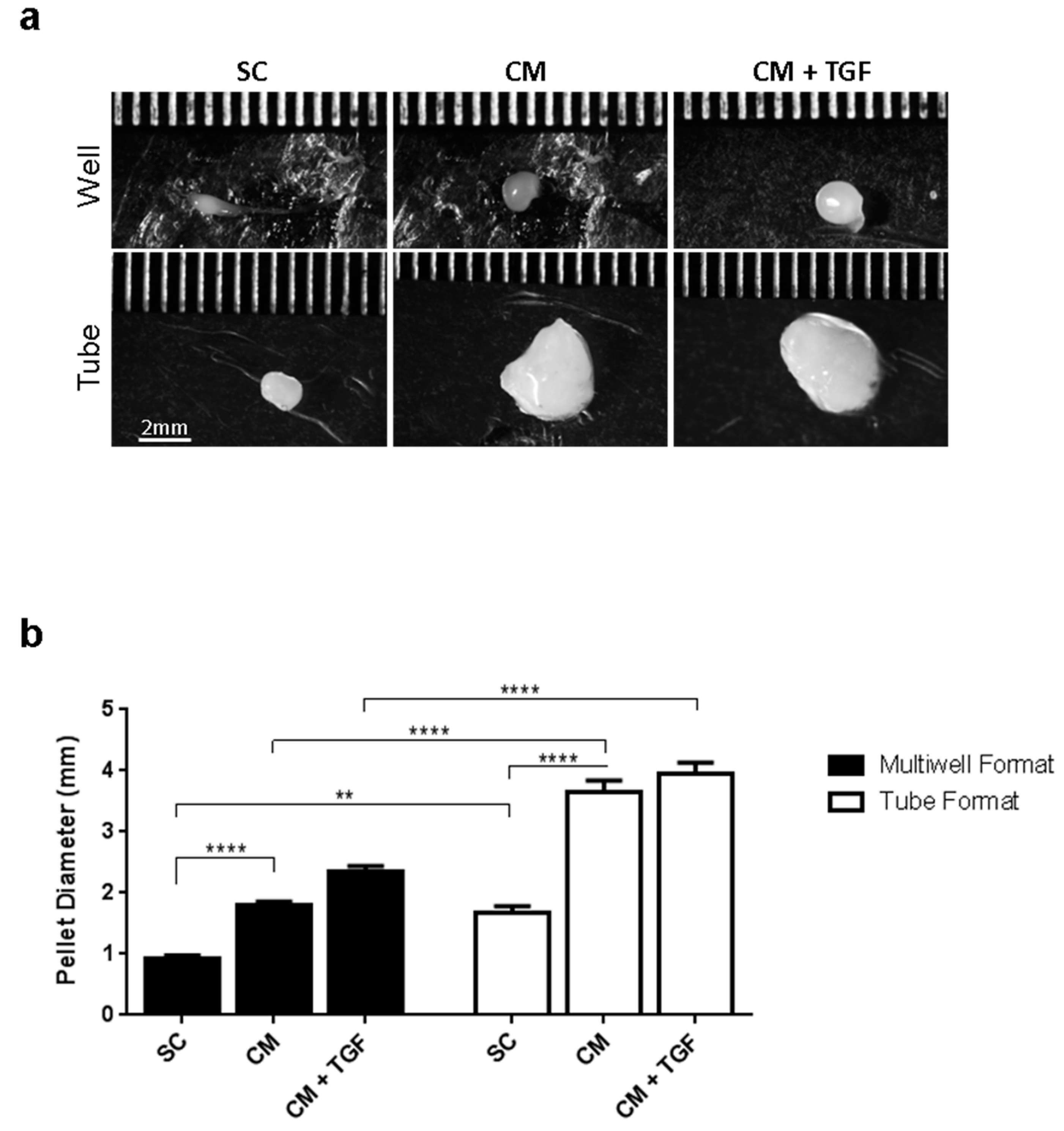
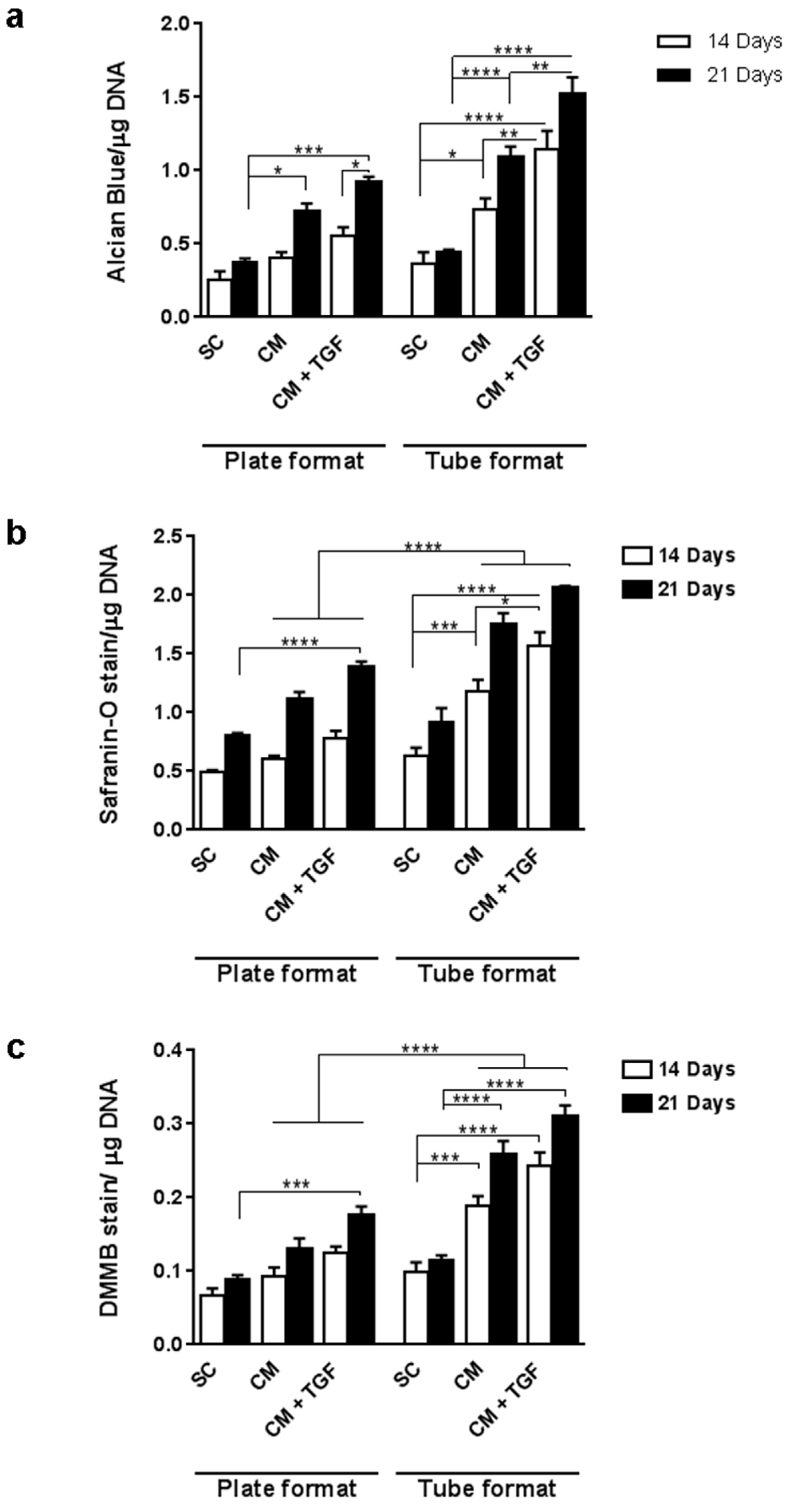
2.3. Comparing the Standard 3D Tube and the Micropellet Format for Chondrogenic Differentiation
3. Discussion
3.1. Chondrogenic Response in 2D vs. 3D Culture Formats
3.2. 3D In-Well Micropellets for Chondrogenic Screening
3.3. Quantitative In-Well Assays for a High Throughput Differentiation Format
4. Materials and Methods
4.1. Cell Culture
4.2. Alcian Blue Quantification of GAG Production
4.3. 1,9-Dimethylmethylene Blue (DMMB) Assay of GAG Production
4.4. Quantification of GAG Production Using Safranin-O
4.5. Pellet Histology
4.6. Quantification of Luciferase Expression
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| hMSC | human mesenchymal stem cells |
| DMMB | 1,9-dimethylmethylene blue |
| GAG | glycosaminoglycans |
References
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Noth, U.; Tuli, R.; Osyczka, A.M.; Danielson, K.G.; Tuan, R.S. In vitro engineered cartilage constructs produced by press-coating biodegradable polymer with human mesenchymal stem cells. Tissue Eng. 2002, 8, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Sottile, V.; Halleux, C.; Bassilana, F.; Keller, H.; Seuwen, K. Stem cell characteristics of human trabecular bone-derived cells. Bone 2002, 30, 699–704. [Google Scholar] [CrossRef]
- Lee, R.H.; Kim, B.; Choi, I.; Kim, H.; Choi, H.S.; Suh, K.; Bae, Y.C.; Jung, J.S. Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell. Physiol. Biochem. 2004, 14, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal Stem Cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Haynesworth, S.E.; Caplan, A.I.; Bruder, S.P. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J. Cell. Biochem. 1997, 64, 295–312. [Google Scholar] [CrossRef]
- Johnstone, B.; Hering, T.M.; Caplan, A.I.; Goldberg, V.M.; Yoo, J.U. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Ratner, B.D.; Hoffman, A.S.; Schoen FJLemons, J.E. Biomaterials Science: An Introduction to Materials in Medicine; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Yoo, J.U.; Barthel, T.S.; Nishimura, K.; Solchaga, L.; Caplan, A.I.; Goldberg, V.M.; Johnstone, B. The chondrogenic potential of human bone-marrow-derived mesenchymal progenitor cells. J. Bone Jt. Surg. Am. 1998, 80, 1745–1757. [Google Scholar] [CrossRef]
- Wang, W.G.; Lou, S.Q.; Ju, X.D.; Xia, K.; Xia, J.H. In vitro chondrogenesis of human bone marrow-derived mesenchymal progenitor cells in monolayer culture: Activation by transfection with TGF-beta2. Tissue Cell 2003, 35, 69–77. [Google Scholar] [CrossRef]
- Yang, H.N.; Park, J.S.; Woo, D.G.; Jeon, S.Y.; Do, H.J.; Lim, H.Y.; Kim, S.W.; Kim, J.H.; Park, K.H. Chondrogenesis of mesenchymal stem cells and dedifferentiated chondrocytes by transfection with SOX Trio genes. Biomaterials 2011, 32, 7695–7704. [Google Scholar] [CrossRef]
- France, L.A.; Scotchford, C.A.; Grant, D.M.; Rashidi, H.; Popov, A.A.; Sottile, V. Transient serum exposure regimes to support dual differentiation of human mesenchymal stem cells. J. Tissue Eng. Regen. Med. 2014, 8, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Tataria, M.; Quarto, N.; Longaker, M.T.; Sylvester, K.G. Absence of the p53 tumor suppressor gene promotes osteogenesis in mesenchymal stem cells. J. Pediatr. Surg. 2006, 41, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Sottile, V.; Seuwen, K. A high-capacity screen for adipogenic differentiation. Anal. Biochem. 2001, 293, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Okabe, T.; Ikawa, T.; Iida, T.; Yasuda, H.; Nakamura, H.; Wakitani, S. Articular cartilage repair with autologous bone marrow mesenchymal cells. J. Cell. Physiol. 2010, 225, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Grassel, S.; Lorenz, J. Tissue-engineering strategies to repair chondral and osteochondral tissue in osteoarthritis: Use of mesenchymal stem cells. Curr. Rheumatol. Rep. 2014, 16, 452. [Google Scholar] [CrossRef]
- Bornes, T.D.; Adesida, A.B.; Jomha, N.M. Mesenchymal stem cells in the treatment of traumatic articular cartilage defects: A comprehensive review. Arthritis Res. Ther. 2014, 16, 432. [Google Scholar] [CrossRef]
- Penick, K.J.; Solchaga, L.A.; Welter, J.F. High-throughput aggregate culture system to assess the chondrogenic potential of mesenchymal stem cells. Biotechniques 2005, 39, 687–691. [Google Scholar] [CrossRef] [Green Version]
- Bjornsson, S. Quantitation of proteoglycans as glycosaminoglycans in biological fluids using an alcian blue dot blot analysis. Anal. Biochem. 1998, 256, 229–237. [Google Scholar] [CrossRef]
- Chang, C.H.; Lin, H.Y.; Fang, H.W.; Loo, S.T.; Hung, S.C.; Ho, Y.C.; Chen, C.C.; Lin, F.H.; Liu, H.C. Chondrogenesis from Immortalized Human Mesenchymal Stem Cells: Comparison Between Collagen Gel and Pellet Culture Methods. Artif. Organs 2008, 32, 561–566. [Google Scholar] [CrossRef]
- Mueller, M.; Blunk, T.; Appel, B.; Maschke, A.; Goepferich, A.; Zellner, J.; Englert, C.; Prantl, L.; Kujat, R.; Nerlich, M.; et al. Insulin is essential for in vitro chondrogenesis of mesenchymal progenitor cells and influences chondrogenesis in a dose-dependent manner. Int. Orthop. 2013, 37, 153–158. [Google Scholar] [CrossRef]
- Ji, Y.H.; Ji, J.L.; Sun, F.Y.; Zeng, Y.Y.; He, X.H.; Zhao, J.X.; Yu, Y.; Yu, S.H.; Wu, W. Quantitative proteomics analysis of chondrogenic differentiation of C3H10T1/2 mesenchymal stem cells by iTRAQ labeling coupled with on-line two-dimensional LC/MS/MS. Mol. Cell. Proteom. 2010, 9, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Fukada, K.; Shibata, S.; Suzuki, S.; Ohya, K.; Kuroda, T. In situ hybridisation study of type I, II, X collagens and aggrecan mRNas in the developing condylar cartilage of fetal mouse mandible. J. Anat. 1999, 195 Pt 3, 321–329. [Google Scholar] [CrossRef]
- Kato, M.; Takaishi, H.; Yoda, M.; Tohmonda, T.; Takito, J.; Fujita, N.; Hosogane, N.; Horiuchi, K.; Kimura, T.; Okada, Y.; et al. GRIP1 enhances estrogen receptor alpha-dependent extracellular matrix gene expression in chondrogenic cells. Osteoarthr. Cartil. 2010, 18, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Minina, E.; Schneider, S.; Rosowski, M.; Lauster, R.; Vortkamp, A. Expression of Fgf and Tgfbeta signaling related genes during embryonic endochondral ossification. Gene Expr. Patterns 2005, 6, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Pelton, R.W.; Saxena, B.; Jones, M.; Moses, H.L.; Gold, L.I. Immunohistochemical localization of TGF beta 1, TGF beta 2, and TGF beta 3 in the mouse embryo: Expression patterns suggest multiple roles during embryonic development. J. Cell Biol. 1991, 115, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Bertolo, A.; Arcolino, F.; Capossela, S.; Taddei, A.R.; Baur, M.; Potzel, T.; Stoyanov, J. Growth Factors Cross-Linked to Collagen Microcarriers Promote Expansion and Chondrogenic Differentiation of Human Mesenchymal Stem Cells. Tissue Eng. Part A 2015, 21, 2618–2628. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.K.; Huey, D.J.; Hu, J.C.; Athanasiou, K.A. TGF-beta1, GDF-5, and BMP-2 stimulation induces chondrogenesis in expanded human articular chondrocytes and marrow-derived stromal cells. Stem Cells 2015, 33, 762–773. [Google Scholar] [CrossRef]
- Suchorska, W.M.; Lach, M.S.; Richter, M.; Kaczmarczyk, J.; Trzeciak, T. Bioimaging: An Useful Tool to Monitor Differentiation of Human Embryonic Stem Cells into Chondrocytes. Ann. Biomed. Eng. 2016, 44, 1845–1859. [Google Scholar] [CrossRef]
- Henderson, S.E.; Santangelo, K.S.; Bertone, A.L. Chondrogenic effects of exogenous retinoic acid or a retinoic acid receptor antagonist (LE135) on equine chondrocytes and bone marrow-derived mesenchymal stem cells in monolayer culture. Am. J. Vet. Res. 2011, 72, 884–892. [Google Scholar] [CrossRef]
- Wang, Z.C.; Sun, H.J.; Li, K.H.; Fu, C.; Liu, M.Z. Icariin promotes directed chondrogenic differentiation of bone marrow mesenchymal stem cells but not hypertrophy. Exp. Ther. Med. 2014, 8, 1528–1534. [Google Scholar] [CrossRef]
- Shirasawa, S.; Sekiya, I.; Sakaguchi, Y.; Yagishita, K.; Ichinose, S.; Muneta, T. In vitro chondrogenesis of human synovium-derived mesenchymal stem cells: Optimal condition and comparison with bone marrow-derived cells. J. Cell. Biochem. 2006, 97, 84–97. [Google Scholar] [CrossRef]
- Goncalves, A.; Costa, P.; Rodrigues, M.T.; Dias, I.R.; Reis, R.L.; Gomes, M.E. Effect of flow perfusion conditions in the chondrogenic differentiation of bone marrow stromal cells cultured onto starch based biodegradable scaffolds. Acta Biomater. 2011, 7, 1644–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsey, R.W.; Tare, R.; Oreffo, R.O.; Chaudhuri, J.B. Perfusion bioreactor studies of chondrocyte growth in alginate-chitosan capsules. Biotechnol. Appl. Biochem. 2012, 59, 142–152. [Google Scholar] [CrossRef]
- Kock, L.M.; Malda, J.; Dhert, W.J.; Ito, K.; Gawlitta, D. Flow-perfusion interferes with chondrogenic and hypertrophic matrix production by mesenchymal stem cells. J. Biomech. 2014, 47, 2122–2129. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.T.B.; Tanavde, V.M.; Hui, J.H.; Lee, E.H. Upregulation of Adipogenesis and Chondrogenesis in MSC Serum-Free Culture. Cell Med. 2011, 2, 27–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welter, J.F.; Solchaga, L.A.; Penick, K.J. Simplification of aggregate culture of human mesenchymal stem cells as a chondrogenic screening assay. Biotechniques 2007, 42, 732–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashidi, H.; Strohbuecker, S.; Jackson, L.; Kalra, S.; Blake, A.J.; France, L.; Tufarelli, C.; Sottile, V. Differences in the pattern and regulation of mineral deposition in human cell lines of osteogenic and non-osteogenic origin. Cells Tissues Organs 2012, 195, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Farndale, R.W.; Sayers, C.A.; Barrett, A.J. A direct spectrophotometric microassay for sulfated glycosaminoglycans in cartilage cultures. Connect. Tissue Res. 1982, 9, 247–248. [Google Scholar] [CrossRef]
- Kan, A.; Ikeda, T.; Saito, T.; Yano, F.; Fukai, A.; Hojo, H.; Ogasawara, T.; Ogata, N.; Nakamura, K.; Chung, U.I.; et al. Screening of chondrogenic factors with a real-time fluorescence-monitoring cell line ATDC5-C2ER: Identification of sorting nexin 19 as a novel factor. Arthritis Rheum. 2009, 60, 3314–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prosser, A.; Scotchford, C.; Roberts, G.; Grant, D.; Sottile, V. Integrated Multi-Assay Culture Model for Stem Cell Chondrogenic Differentiation. Int. J. Mol. Sci. 2019, 20, 951. https://doi.org/10.3390/ijms20040951
Prosser A, Scotchford C, Roberts G, Grant D, Sottile V. Integrated Multi-Assay Culture Model for Stem Cell Chondrogenic Differentiation. International Journal of Molecular Sciences. 2019; 20(4):951. https://doi.org/10.3390/ijms20040951
Chicago/Turabian StyleProsser, Amy, Colin Scotchford, George Roberts, David Grant, and Virginie Sottile. 2019. "Integrated Multi-Assay Culture Model for Stem Cell Chondrogenic Differentiation" International Journal of Molecular Sciences 20, no. 4: 951. https://doi.org/10.3390/ijms20040951
APA StyleProsser, A., Scotchford, C., Roberts, G., Grant, D., & Sottile, V. (2019). Integrated Multi-Assay Culture Model for Stem Cell Chondrogenic Differentiation. International Journal of Molecular Sciences, 20(4), 951. https://doi.org/10.3390/ijms20040951